
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Факторы врожденного иммунитета ⇐ ПредыдущаяСтр 6 из 6
Если рассматривать траекторию движения патогенного микроба в динамике инфекционного процесса, то легко заметить, что организм на этом пути выстраивает различные линии защиты (табл. 9.1). Прежде всего это покровный эпителий кожи и слизистых оболочек, обладающий колонизационной резистентностью. Если возбудитель вооружен соответствующими инвазивными факторами, то он проникает в субэпителиальную ткань, где развивается острая воспалительная реакция, ограничивающая возбудителя во входных воротах. Следующая станция на пути патогена - регионарные лимфатические узлы, куда он транспортируется лимфой по лимфатическим сосудам, дренирующим соединительную ткань. Лимфатические сосуды и узлы отвечают на внедрение развитием лимфангита и лимфаденита. После преодоления этого барьера микробы по эфферентным лимфатическим сосудам проникают в кровь - в ответ может развиться системный воспалительный от- вет. Если микроб не гибнет в крови, то он гематогенно разносится во внутренние органы - развиваются генерализованные формы инфекции. Таблица 9.1. Факторы и механизмы антиинфекционного иммунитета (принцип эшелонированности антимикробной защиты по Маянскому А.Н., 2003)
• кожу и слизистые оболочки; • клеточные факторы: нейтрофилы, макрофаги, дендритные клетки, эозинофилы, базофилы, естественные киллеры; • гуморальные факторы: система комплемента, растворимые рецепторы к поверхностным структурам микроорганизмов (pattern-структуры), антимикробные пептиды, интерфероны. Кожа и слизистые оболочки. Тонкий слой эпителиальных клеток, выстилающий поверхность кожи и слизистых оболочек, является тем барьером, который практически непроницаем для микроорганизмов. Он отделяет стерильные ткани организма от заселенного микробами внешнего мира. Кожа покрыта многослойным плоским эпителием, в котором различают два слоя: роговой и базальный. Кератиноциты рогового слоя - это погибшие клетки, устойчивые к агрессивным химическим соединениям. На их поверхности отсутствуют рецепторы для адгезивных молекул микроорганизмов, поэтому они обладают значительной устойчивостью к колонизации и являются самым надежным барьером на пути большинства бактерий, грибов, вирусов, простейших. Исключение составляют S. aureus, Pr. acnae, I. pestis, да и они скорее всего проникают либо через микротрещины, либо с помощью кровососущих насекомых, либо через устья потовых и сальных желез. Устье сальных и потовых желез, волосяных фоликулов в коже наиболее уязвимы, поскольку здесь слой ороговевшего эпителия истончается. В защите этих участков важную роль играют продукты потовых и сальных желез, содержащих молочную, жирные кислоты, ферменты, антибактериальные пептиды, оказывающие антимикробное действие. Именно в устьях придатков кожи располагается глубокая резидентная микрофлора, образующая микроколонии и продуцирующая защитные факторы (см. главу 4). В эпидермисе, помимо кератиноцитов, содержатся еще два типа клеток - клетки Лангерганса и клетки Гринстейна (отростчатые эпидермоциты, составляющие 1-3% кариоцитов базального слоя). Клетки Лангерганса и Гринстейна имеют миелоидное происхождение и относятся к дендритным. Предполагается, что по функции эти клетки являются оппозитными. Клетки Лангерганса участвуют в презентации антигена, индуцируют иммунный ответ, а клетки Гринстейна продуцируют цитокины, подавляющие им- мунные реакции в коже. Типичные кератиноциты и дендритные клетки эпидермиса в совокупности с лимфоидными структурами дермы принимают активное участие в реакциях приобретенного иммунитета (см. ниже). Здоровая кожа обладает высокой способностью к самоочищению. Это легко доказать, если нанести на ее поверхность нетипичные для кожи бактерии - через некоторое время такие микробы исчезают. На этом принципе основаны методы оценки бактерицидной функции кожи. Слизистые оболочки. Большинство инфекций начинается не с кожи, а со слизистых оболочек. Это связано, во-первых, с большей площадью их поверхности (слизистые оболочки около 400 м2, кожа около 2 м2), во-вторых, с меньшей защищенностью. Слизистые оболочки не имеют многослойного плоского эпителия. На их поверхности располагается лишь один слой эпителиоцитов. В кишечнике это однослойный цилиндрический эпителий, бокаловидные секреторные клетки и М-клетки (мембранные эпителиальные клетки), располагающиеся в слое эпителиоцитов, покрывающие лимфоидные скопления. М-клетки наиболее уязвимы для проникновения многих патогенных микроорганизмов изза целого ряда особенностей: наличия специфических рецепторов для некоторых микроорганизмов (сальмонелл, шигелл, патогенных эшерихий и др.), которые не обнаружены на соседних энтероцитах; истонченного слизистого слоя; способности к эндоцитозу и пипоцитозу, благодаря чему обеспечивается облегченный транспорт антигенов и микроорганизмов из кишечной трубки в мукозоассоциированную лимфоидную ткань (см. главу 12); отсутствия мощного лизосомального аппарата, характерного для макрофагов и нейтрофилов, благодаря чему бактерии и вирусы перемещаются в субэпителиальное пространство без разрушения. М-клетки относятся к эволюционно сформировавшейся системе облегченного транспорта антигенов к иммунокомпетентным клеткам, а бактерии и вирусы используют этот путь для своей транслокации через эпителиальный барьер. Аналогичные М-клеткам кишечника эпителиоциты, ассоциированные с лимфоидной тканью, имеются у слизистых оболочек бронхоальвеолярного дерева, носоглотки, половой системы. Колонизационная резистентность покровного эпителия. Любой инфекционный процесс начинается с адгезии возбудителя на по- верхности чувствительных эпителиоцитов (за исключением микроорганизмов, передающихся через укусы насекомых или вертикально, т.е. от матери к плоду). Только закрепившись, микробы приобретают возможность размножиться во входных воротах и образовывать колонию. В колонии накапливаются токсины, ферменты патогенности в количестве, необходимом для преодоления эпителиального барьера. Этот процесс называется колонизацией. Под колонизационной резистентностью понимают устойчивость эпителия кожи и слизистых оболочек к заселению посторонними микроорганизмами. Колонизационную резистентность слизистых оболочек обеспечивает муцин, секретируемый бокаловидными клетками и образующий на поверхности сложноорганизованную биопленку. В этот биослой встроены все защитные инструменты: резидентная микрофлора, бактерицидные вещества (лизоцим, лактоферрин, токсичные метаболиты кислорода, азота и т.д.), секреторные иммуноглобулины, фагоциты. Роль нормальной микрофлоры (см. главу 4.3). Важнейшим механизмом участия резидентной микрофлоры в колонизационной резистентности является их способность продуцировать бактериоцины (антибиотикоподобные субстанции), короткоцепочечные жирные кислоты, молочную кислоту, сероводород, перекись водорода. Такими свойствами обладают лакто-, бифидобактерии, бактероиды. Благодаря ферментативной активности анаэробных бактерий в кишечнике происходит деконъюгация желчных кислот с образованием дезоксихолиевой кислоты, токсичной для патогенных и условно-патогенных бактерий. Муцин наряду с полисахаридами, продуцируемыми резидентными бактериями (в частности, лактобактериями), образует на поверхности слизистых оболочек выраженный гликоналикс (биопленку), который эффективно экранирует сайты адгезии и делает их недоступными для случайных бактерий. Бокаловидные клетки образуют смесь сиало- и сульфомуцинов, соотношение которых различается в различных биотонах. Своеобразие состава микрофлоры в различных экологических нишах в значительной степени определяется количеством и качеством муцина. Фагоцитирующие клетки и продукты их дегрануляции. В слизистый биослой на поверхности эпителия мигрируют макрофаги и нейтрофилы. Наряду с фагоцитозом эти клетки выделяют биоцид- ные продукты наружу, содержащиеся в их лизосомах (лизоцим, пероксидаза, лактоферрин, дефанзины, токсичные метаболиты кислорода, азота), которые повышают антимикробные свойства секретов. Химические и механические факторы. В резистентности покровного эпителия слизистых оболочек важную роль играют секреты, обладающие выраженными биоцидными, антиадгезивными свойствами: слеза, слюна, желудочный сок, ферменты и желчные кислоты тонкой кишки, цервикальный и вагинальный секреты репродуктивной системы женщин. Благодаря целенаправленным движениям - перистальтике гладкой мускулатуры в кишечнике, ресничек мерцательного эпителия в респираторном тракте, мочи в мочевыводящей системе - образующиеся секреты вместе с содержащимися в них микроорганизмами перемещаются в направлении выхода и выводятся наружу. Колонизационная резистентность слизистых оболочек усиливается за счет секреторных иммуноглобулинов А, синтезируемых мукозоассоциированной лимфоидной тканью. Покровный эпителий мукозального тракта постоянно регенерирует за счет стволовых клеток, расположенных в толще слизистых оболочек. В кишечнике эту функцию выполняют клетки крипт, в которых наряду со стволовыми располагаются клетки Панета - особые клетки, синтезирующие антибактериальные белки (лизоцим, катионные пептиды). Эти белки защищают не только стволовые клетки, но и покровные эпителиоциты. При воспалении в стенке слизистой оболочки продукция этих белков усиливается. Колонизационная резистентность покровного эпителия обеспечивается всей совокупностью защитных механизмов врожденного и приобретенного (секреторные иммуноглобулины) иммунитета и является основой устойчивости организма к большинству микроорганизмов, обитающих во внешней среде. Отсутствие на эпителиоцитах специфических рецепторов для определенных микроорганизмов, по-видимому, является базовым механизмом генетической резистентности животных одного вида к микробам, патогенным для животных другого вида. Клеточные факторы Нейтрофилы и макрофаги. Способностью к эндоцитозу (поглощению частиц с образованием внутриклеточной вакуоли) обла- дают все эукариотические клетки. Именно таким образом внутрь клеток проникают многие патогенные микроорганизмы. Однако в большинстве инфицированных клеток отсутствуют механизмы (либо они слабы), обеспечивающие деструкцию патогена. В процессе эволюции в организме многоклеточных сформировались специализированные клетки, имеющие мощные системы внутриклеточного киллинга, основной «профессией» которых является фагоцитоз (от греч. phagos - пожираю, cytos - клетка) - поглощение частиц диаметром не менее 0, 1 мкм (в отличие от пиноцитоза - поглощения частиц меньшего диаметра и макромолекул) и уничтожение захваченных микробов. Такими свойствами обладают полиморфно-ядерные лейкоциты (в основном нейтрофилы) и мононуклеарные фагоциты (эти клетки иногда называют профессиональными фагоцитами). Впервые идея о защитной роли подвижных клеток (микро- и макрофагов) была сформулирована в 1883 г. И.И. Мечниковым, который за создание клеточно-гуморальной теории иммунитета (в соавторстве с П. Эрлихом) был удостоен в 1909 г. Нобелевской премии. Нейтрофилы и мононуклеарные фагоциты имеют общее миелоидное происхождение из стволовой кроветворной клетки. Однако эти клетки различаются рядом свойств. Нейтрофилы - наиболее многочисленная и подвижная популяция фагоцитов, созревание которых начинается и заканчивается в костном мозгу. Около 70% всех нейтрофилов сохраняется в виде резерва в костно-мозговых депо, откуда они под влиянием соответствующих стимулов (провоспалительных цитокинов, продуктов микробного происхождения, С5а-компонента комплемента, колониестимулирующих факторов, кортикостероидов, катехоламинов) могут экстренно перемещаться через кровь в очаг тканевой деструкции и участвовать в развитии острого воспалительного ответа. Нейтрофилы - это «отряд быстрого реагирования» в системе антимикробной защиты. Нейтрофилы - короткоживущие клетки, продолжительность их жизни около 15 сут. Из костного мозга они выходят в кровоток уже зрелыми клетками, утратившими способность к дифференцированию и пролиферации. Из крови нейтрофилы перемещаются в ткани, в которых они либо гибнут, либо выходят на поверхность слизистых оболочек, где и заканчивают свой жизненный цикл. Мононуклеарные фагоциты представлены промоноцитами костного мозга, моноцитами крови и тканевыми макрофагами. Моноциты, в отличие от нейтрофилов, - незрелые клетки, которые, попадая в кровяное русло и далее в ткани, созревают в тканевые макрофаги (плевральные и перитонеальные, купферовские клетки печени, альвеолярные, интердигитальные клетки лимфатических узлов, костного мозга, остеокласты, микроглиоциты, мезангиальные клетки почек, сертолиевы клетки яичек, клетки Лангерганса и Гринстейна кожи). Продолжительность жизни мононуклеарных фагоцитов от 40 до 60 сут. Макрофаги - не очень быстрые клетки, но они рассеяны во всех тканях, и, в отличие от нейтрофилов, им нет необходимости в столь срочной мобилизации. Если продолжить аналогию с нейтрофилами, то макрофаги в системе врожденного иммунитета - это «войска специального назначения». Важной особенностью нейтрофилов и макрофагов является наличие в их цитоплазме большого количества лизосом - гранул размером 200-500 нм, содержащих различные ферменты, бактерицидные и биологически активные продукты (лизоцим, миелопероксидаза, дефензины, бактерицидный протеин, лактоферрин, протеиназы, катепсины, коллагеназа и т.д.). Благодаря столь разнообразному «вооружению» фагоциты обладают мощным деструктивным и регуляторным потенциалом. Нейтрофилы и макрофаги чутко реагируют на любые изменения гомеостаза. Для этой цели они оснащены богатым арсеналом рецепторов, располагающихся на их цитоплазматической мембране (рис. 9.2): • рецепторы для распознавания чужого - Toll-подобные рецепторы (Toll-like receptor - TLR), впервые открытые А. Poltorak в 1998 г. у плодовой мушки и впоследствии обнаруженные у нейтрофилов, макрофагов и дендритных клеток. По значимости открытие Toll-подобных рецепторов сопоставимо с более ранним обнаружением антигенраспознающих рецепторов у лимфоцитов. Toll-подобные рецепторы узнают не антигены, разнообразие которых в природе чрезвычайно велико (около 1018 вариантов), а более грубые повторяющиеся молекулярные углеводные и липидные узоры - pattern-структуры (от англ. рattern - узор), которых нет на клетках организма хозяина, но которые присутствуют у простейших, грибов, бактерий, вирусов. Репертуар таких узоров невелик и составляет около 20 ва-
риантов. Toll-подобные рецепторы представляют собой семейство мембранных гликопротеидов, известно 11 типов таких рецепторов, способных узнавать всю палитру pattern-структур микроорганизмов (липополисахариды, глико-, липопротеи- ды, нуклеиновые кислоты, белки теплового шока и т.д.). Взаимодействие Toll-подобных рецепторов с соответствующими лигандами запускает транскрипцию генов провоспалительных цитокинов и ко-стимулирующих молекул, которые необходимы для миграции, адгезии клеток, фагоцитоза и представления антигенов лимфоцитам; • маннозно-фукозные рецепторы, распознающие углеводные компоненты поверхностных структур микроорганизмов; • рецепторы для мусора (scavenger receptor) - для связывания фосфолипидных мембран и компонентов собственных разрушенных клеток. Участвуют в фагоцитозе поврежденных и умирающих клеток; • рецепторы для С3в- и С4в-компонентов комплемента; • рецепторы для Fc-фрагментов IgG. Эти рецепторы, как и рецепторы для компонентов комплемента, играют важную роль в связывании иммунных комплексов и фагоцитозе бактерии, помеченных иммуноглобулинами и комплементом (эффект опсонизации); • рецепторы для цитокинов, хемокинов, гормонов, лейкотриенов, простагландинов и т.д. позволяют взаимодействовать с лимфоцитами и реагировать на любые изменения внутренней среды организма. Основной функцией нейтрофилов и макрофагов является фагоцитоз. Фагоцитоз - это процесс поглощения клеткой частиц или крупных макромолекулярных комплексов. Он складывается из нескольких последовательно протекающих этапов: • активация и хемотаксис - целенаправленное движение клетки к объекту фагоцитоза в сторону повышающейся концентрации хемоаттрактантов, роль которых играют хемокины, компоненты комплемента и микробной клетки, продукты деградации тканей организма; • адгезия (прикрепление) частиц к поверхности фагоцита. В адгезии важную роль играют Toll-подобные рецепторы, а также рецепторы к Fc-фрагменту иммуноглобулина и С3в- компоненту комплемента (такой фагоцитоз называется иммунным). Иммуноглобулины M, G, С3в-, С4в-компоненты комплемента усиливают адгезию (являются опсонинами), служат мостиком между микробной клеткой и фагоцитом; • поглощение частиц, их погружение в цитоплазму и образование вакуоли (фагосомы); • внутриклеточный киллинг (убийство) и переваривание. После поглощения частицы фагосомы сливаются с лизосомами - образуется фаголизосома, в которой бактерии гибнут под действием бактерицидных продуктов гранул (кислороднезависимая система бактерицидности). Одновременно в клетке усиливается потребление кислорода и глюкозы - развивается так называемый респираторный (окислительный) взрыв, что приводит к образованию токсичных метаболитов кислорода и азота (Н2О2, супероксиданиона О2, гипохлорной кислоты, пироксинитрита), обладающих высокой бактерицидностью (кислородзависимая система бактерицидности). Не все микроорганизмы чувствительны к бактерицидным системам фагоцитов. Гонококки, стрептококки, микобактерии и другие выживают после контакта с фагоцитами, такой фагоцитоз называется незавершенным. Фагоциты, помимо фагоцитоза (эндоцитоза), могут осуществлять свои цитотоксические реакции путем экзоцитоза - выделения своих гранул наружу (дегрануляция) - таким образом фагоциты осуществляют внеклеточный киллинг. Нейтрофилы, в отличие от макрофагов, способны образовывать внеклеточные бактерицидные ловушки - в процессе активации клетка выбрасывает наружу нити ДНК, в которых располагаются гранулы с бактерицидными ферментами. Благодаря липкости ДНК бактерии приклеиваются к ловушкам и под действием фермента погибают. Нейтрофилы и макрофаги являются важнейшим звеном врожденного иммунитета, однако их роль в защите от различных микробов неодинакова. Нейтрофилы эффективны при инфекциях, вызванных внеклеточными патогенами (гноеродные кокки, энтеробактерии и др.), индуцирующими развитие острого воспалительного ответа. При таких инфекциях эффективна кооперация нейтрофил-комплемент-антитело. Макрофаги защищают от внутриклеточных патогенов (микобактерии, риккетсии, хламидии и др.), вызывающих развитие хронического гранулематозного воспаления, где главную роль играет кооперация макрофаг-Т- лимфоцит. Помимо участия в антимикробной защите, фагоциты участвуют в удалении из организма отмирающих, старых клеток и продуктов их распада, неорганических частиц (уголь, минеральная пыль и др.). Фагоциты (особенно макрофаги) являются антигенпред- ставляющими, они обладают секреторной функцией, синтезируют и выделяют наружу широкий спектр биологически активных соединений: цитокины (интерлейкины-1, 6, 8, 12, фактор некроза опухоли), простагландины, лейкотриены, интерфероны α и γ. Благодаря этим медиаторам фагоциты активно участвуют в поддержании гомеостаза, в процессах воспаления, в адаптивном иммунном ответе, регенерации. Эозинофилы относятся к полиморфно-ядерным лейкоцитам. Они отличаются от нейтрофилов тем, что обладают слабой фагоцитарной активностью. Эозинофилы поглощают некоторые бактерии, но внутриклеточный киллинг у них менее эффективен, чем у нейтрофилов. Основная функция эозинофилов заключается в защите от крупных паразитов. После активации эти клетки выделяют токсичные продукты своих гранул, оказывающих губительное действие на гельмины. К таким продуктам относят: катионный белок - РНКазу, контакт с которой приводит к образованию мембранных каналов в оболочке паразита; пероксидазу эозинофилов, которая, в отличие от пероксидазы нейтрофилов, окисляя субстраты, приводит к образованию гипогалидов, высокотоксичных для некоторых паразитов; главный основной белок эозинофилов - основной компонент гранул, который способен полимеризоваться в оболочке паразита с образованием трансмембранных пор, через которые внутрь мишени проникают другие медиаторы. Естественные киллеры. Естественные киллеры - большие лимфоцитоподобные клетки, которые происходят из лимфоидных предшественников. Они содержатся в крови, тканях, особенно их много в печени, слизистой оболочке репродуктивной системы женщин, селезенке. Естественные киллеры, как и фагоциты, содержат лизосомы, но фагоцитарной активностью не обладают. Естественные киллеры распознают и элиминируют клеткимишени, на которых изменены или отсутствуют маркеры, характерные для здоровых клеток. Известно, что такое происходит прежде всего с клетками, мутировавшими или пораженными вирусом. Именно поэтому естественные киллеры играют важную роль в противоопухолевом надзоре, уничтожении клеток, зараженных вирусами. Свое цитотоксическое действие естественные киллеры оказывают с помощью особого белка перфорина, который подобно мембраноатакующему комплексу комплемента образует поры в мембранах клеток-мишеней. Гуморальные факторы Система комплемента. Система комплемента - это многокомпонентная полиферментная самособирающаяся система сывороточных белков, которые в норме находятся в неактивном состоянии. При появлении во внутренней среде микробных продуктов запускается процесс, который называют активацией комплемента. Активация протекает по типу каскадной реакции, когда каждый предшествующий компонент системы активирует последующий. В процессе самосборки системы образуются активные продукты распада белков, которые выполняют три важнейшие функции: вызывают перфорацию мембран и лизис клеток, обеспечивают опсонизацию микроорганизмов для их дальнейшего фагоцитоза и инициируют развитие сосудистых реакций воспаления. Комплемент под названием «алексин» был описан в 1899 г. французским микробиологом Ж. Борде, а затем немецким микробиологом П. Эрлихом назван комплементом (complement - дополнение) как фактор, дополнительный к антителам, вызывающим лизис клеток. В систему комплемента входит 9 основных белков (обозначаемых как С1, С2-С9), а также субкомпоненты - продукты расщепления этих белков (Clg, С3в, С3а и т.д.), ингибиторы. Ключевым событием для системы комплемента является его активация. Она может происходить тремя путями: классическим, лектиновым и альтернативным (рис. 9.3). Классический путь. При классическом пути активирующим фактором являются комплексы антиген-антитело. При этом Fс-фрагмент и IgG иммунных комплексов активирует Сгсубкомпонент, Сг расщепляется с образованием Cls, гидролизующей С4, который расщепляется на С4а (анафилотоксин) и С4в. С4в активирует С2, который, в свою очередь, активизирует С3- компонент (ключевой компонент системы). С3-компонент расщепляется на анафилотоксин С3а и опсонин С3в. Активация С5- компонента комплемента также сопровождается образованием двух активных фрагментов белков: С5а - анафилотоксина, хемоаттрактанта для нейтрофилов и С5в - активирующего С6-компонент. В итоге образуется комплекс С5, б, 7, 8, 9, который называется мембраноатакующим. Терминальная фаза активации комплемента - это образование трансмембранной поры в клетке, выход ее содержимого наружу. В итоге клетка набухает и лизируется.
Лектиновый путь. Он во многом аналогичен классическому. Различие заключается лишь в том, что при лектиновом пути один из белков острой фазы - связывающий маннозу лектин взаимодействует с маннозой на поверхности микробных клеток (прообраз комплекса антиген-антитело), и этот комплекс активирует С4 и С2. Альтернативный путь. Он идет без участия антител и минуя первые 3 компонента С1-С4-С2. Инициируют альтернативный путь компоненты клеточной стенки грамотрицательных бактерий (липополисахариды, пептидогликаны), вирусы, которые связываются последовательно с белками Р (пропердин), В и D. Эти комплексы напрямую конвертируют С3-компонент. Сложная каскадная реакция комплемента протекает только в присутствии ионов Са и Mg. Биологические эффекты продуктов активации комплемента: • вне зависимости от пути активация комплемента завершается образованием мембраноатакующего комплекса (С5, б, 7, 8, 9) и лизисом клеток (бактерий, эритроцитов и других клеток); • образующиеся С3а-, С4а- и С5а-компоненты являются анафилотоксинами, они связываются с рецепторами кровяных и тканевых базофилов, индуцируют их дегрануляцию - выброс гистамина, серотонина и других вазоактивных медиаторов (медиаторов воспалительного ответа). Кроме этого С5а является хемоаттрактантом для фагоцитов, он привлекает эти клетки в очаг воспаления; • С3в, С4в являются опсонинами, повышают адгезию иммунных комплексов с мембранами макрофагов, нейтрофилов, эритроцитов и тем самым усиливают фагоцитоз. Растворимые рецепторы для патогенов. Это белки крови, непосредственно связывающиеся с различными консервативными, повторяющимися углеводными или липидными структурами микробной клетки (pattern-структурами). Эти белки обладают опсоническими свойствами, некоторые из них активируют комплемент. Основную часть растворимых рецепторов составляют белки острой фазы. Концентрация этих белков в крови быстро нарастает в ответ на развитие воспаления при инфекции или повреждении тканей. К белкам острой фазы относятся: • С-реактивный белок (он составляет основную массу белков острой фазы), получивший название вследствие способности связываться с фосфорилхолином (С-полисахаридом) пневмококков. Образование комплекса С-реактивный белок- фосфорилхолин способствует фагоцитозу бактерий, поскольку комплекс связывается с Clg и активирует классический путь комплемента. Белок синтезируется в печени, и его концентрация быстро нарастает в ответ на интерлейкин-б; • сывороточный амилоид Р близок по структуре и функции к С-реактивному белку; • маннозосвязывающий лектин активирует комплемент по лектиновому пути, является одним из представителей сывороточных белков-коллектинов, распознающих углеводные остатки и действующих как опсонины. Синтезируется в печени; • белки сурфактанта легких также принадлежат к семейству коллектинов. Обладают опсоническим свойством, особенно в отношении одноклеточного гриба Pneumocystis carinii; • другую группу белков острой фазы составляют белки, связывающие железо, - трансферрин, гаптоглобин, гемопексин. Такие белки препятствуют размножению бактерий, нуждающихся в этом элементе. Антимикробные пептиды. Одним из таких пептидов является лизоцим. Лизоцим - это фермент муромидаза с молекулярной массой 14 000-1б 000, вызывающий гидролиз муреина (пептидогликана) клеточной стенки бактерий и их лизис. Открыт в 1909 г. П.Л. Лащенковым, выделен в 1922 г. А. Флемингом. Лизоцим содержится во всех биологических жидкостях: сыворотке крови, слюне, слезе, молоке. Он продуцируется нейтрофилами и макрофагами (содержится в их гранулах). Лизоцим в большей степени действует на грамположительные бактерии, основу клеточной стенки которых составляет пептидогликан. Клеточные стенки грамотрицательных бактерий также могут повреждаться лизоцимом, если на них предварительно подействовал мембраноатакующий комплекс системы комплемента. Дефензины и кателицидины - пептиды, обладающие антимикробной активностью. Они образуются клетками многих эукариот, содержат 13-18 аминокислотных остатков. На сегодняшний день известно около 500 таких пептидов. У млекопитающих бактерицидные пептиды относятся к семействам дефензинов и кателицидинов. В гранулах человеческих макрофагов, нейтрофилов содержатся α -дефензины. Они синтезируются также эпителиальными клетками кишечника, легких, мочевого пузыря. Семейство интерферонов. Интерферон (ИФН) был открыт в 1957 г. А. Айзексом и Ж. Линдеманом при изучении интерференции вирусов (от лат. inter - между, ferens - несущий). Интерференция - это явление, когда ткани, инфицированные одним вирусом, становятся устойчивыми к заражению другим вирусом. Было установлено, что такая резистентность связана с продукцией зараженными клетками особого белка, который и был назван интерфероном. В настоящее время интерфероны хорошо изучены. Они представляют собой семейство гликопротеидов с молекулярной массой от 15 000 до 70 000. В зависимости от источника получения эти белки делят на интерфероны I и II типов. I тип включает ИФН α и β, которые продуцируются инфицированным вирусом клетками: ИФН-α - лейкоцитами, ИФН-β - фибробластами. В последние годы описаны три новых интерферона: ИФН-τ /ε (трофобластный ИФН), ИФН-λ и ИФН-К. В противовирусной защите участвуют ИФН-α и β. Механизм действия ИФН-α и β не связан с прямым влиянием на вирусы. Он обусловлен активацией в клетке ряда генов, блокирующих репродукцию вируса. Ключевое звено - индукция синтеза протеинкиназы R, которая нарушает трансляцию вирусной мРНК и запускает апоптоз зараженных клеток через Вс1-2 и каспазазависимые реакции. Другой механизм - это активация латентной РНК-эндонуклеазы, которая вызывает деструкцию вирусной нуклеиновой кислоты. II тип включает интерферон γ. Он продуцируется Т-лимфоцитами и естественными киллерами после антигенной стимуляции. Интерферон синтезируется клетками постоянно, его концентрация в крови в норме мало меняется. Однако продукция ИФ усиливается при заражении клеток вирусами или действии его индукторов - интерфероногенов (вирусной РНК, ДНК, сложных полимеров). В настоящее время интерфероны (как лейкоцитарные, так и рекомбинантные) и интерфероногены широко применяются в клинической практике для профилактики и лечения острых вирусных инфекций (грипп), а также с терапевтической целью при хронических вирусных инфекциях (гепатиты В, С, герпес, рассеянный склероз и др.). Поскольку интерфероны обладают не только противовирусной, но и противоопухолевой активностью, они применяются также для лечения онкологических заболеваний. 11. Клеточные факторы врожденного иммунитета полости рта: определение понятия, названия клеток, их функции. (выше) 12. Местный приобретённый иммунитет полости рта: определение понятия. Основные механизмы. Особенности структуры секреторных иммуноглобулинов, их подклассы, функции, место выработки. Местный иммунитет (колонизационная резистентность) — это сложный комплекс защитных приспособлений различной природы, сформировавшийся в процессе эволюционного развития и обеспечивающий защиту слизистых тех органов, которые непосредственно сообщаются с внешней средой. Основная его функция — сохранение гомеостаза внутренней среды макроорганизма, т.е. он является первым барьером на пути микроорганизма и любого антигена. С этой точки зрения, местный иммунитет — неразрывная часть общего иммунитета, и в то же время он составляет четко очерченную и автономную в своих функциях систему. Основные защитные механизмы местного иммунитета полости рта представлены на рис. 11
13. Гуморальные факторы врожденного иммунитета полости рта: определение понятия, названия факторов (молекул), их функции.(выше) 14. Микрофлора при кариесе зубов. Местные факторы, способствующие развитию кариеса. Роль зубной бляшки в развитии кариеса. В настоящее время известно, что кариозный процесс может развиваться при следующих условиях: 1. Наличие достаточного количества углеводов в пище; 2. Наличие микроорганизмов в полости рта; 3. Контакт углеводов и микроорганизмов с зубами. Ярким доказательством роли углеводов в возникновении кариозного процесса являются проведенные экспериментальные исследования. Все кариесогенные диеты содержат более 50 % сахарозы. Без контакта зубов с углеводами кариозный процесс не возникает. Несомненно, важная роль принадлежит составу и структуре эмали зуба, слюне, а также характеру питания, составу питьевой воды. При начальных поражениях обнаруживаются микроорганизмы, которые с точки зрения их биохимической активности могут быть подразделены на две группы: протеолитическую и кислотообразующую. В протеолитическую группу входят бактероиды и пептококки. Они вырабатывают ферменты, способные расщеплять органические вещества кариозного зуба. К кислотообразующей группе относятся стрептококки, лактобактерии и актиномицеты. Из числа стрептококков здесь чаще всего присутствуют энтерококки. Все эти микроорганизмы могут участвовать в процессе деминерализации твердых тканей кариозного зуба, т.к. они интенсивно расщепляют углеводы и образуют много органических кислот. В кариозной полости присутствуют все представители постоянной флоры полости рта, главным образом строгие анаэробы. На кариесогенную активность оральных микроорганизмов влияет слюна - ее агрегирующие факторы, которые, с одной стороны, способствуют прикреплению микробных клеток к поверхности зуба, а с другой - удаляют их при омывании полости рта. Противокариозным действием обладает система буферов, бикарбонат - карбоновая кислота, а также протеин и спалим, находящиеся в слюне. Профилактика кариеса может быть направлена на уменьшение количества кариесогенных микроорганизмов в полости рта. Эффективно применение различных бактерицидных и бактериостатических препаратов. Хорошие результаты получают с помощью антисептиков, в частности 0, 2 % хлоргексидина. При этом количество клеток S. mutans в зубных бляшках снижается на 80 - 85 %, а в слюне на 55 %. Покрывая зубную поверхность, хлоргескидин не только оказывает на микроорганизмы бактерицидное действие, но и препятствует их адгезии, не нарушая при этом микробного равновесия. Угнетающим действием на микроорганизмы обладает фтор и его соединения. Другой путь снижения кислотообразования и накопления глюканов - замена сахарозы другими углеводами, при ферментативном расщеплении которых эти продукты не образуются. Местные факторы: 1. Зубная бляшка и зубной налет, изобилующие микроорганизмами. 2.Нарушение состава и свойств ротовой жидкости, являющейся индикатором состояния организма в целом. Популярное:
|
Последнее изменение этой страницы: 2016-07-13; Просмотров: 9215; Нарушение авторского права страницы
 К факторам врожденного иммунитета относят:
К факторам врожденного иммунитета относят: 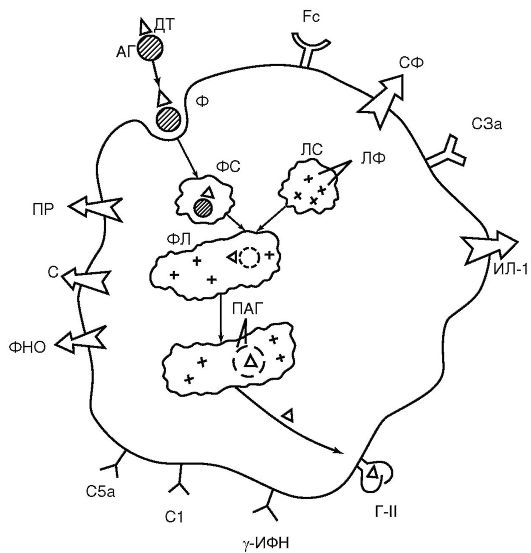 Рис. 9.2. Функциональные структуры макрофага (схема): АГ - антиген; ДТ - антигенная детерминанта; ФС - фагосома; ЛС - лизосома; ЛФ - лизосомальные ферменты; ФЛ - фаголизосома; ПАГ - процессированный антиген; Г-II - антиген гистосовместимости II класса (МНС II); Fc - рецептор для Fc-фрагмента молекулы иммуноглобулина; С1, С3а, С5а - рецепторы для компонентов комплемента; γ -ИФН - рецептор для γ -МФН; С - секреция компонентов комплемента; ПР - секреция перекисных радикалов; ИЛД-1 - секреция; ФНО - секреция фактора некроза опухолей; Сф - секреция ферментов
Рис. 9.2. Функциональные структуры макрофага (схема): АГ - антиген; ДТ - антигенная детерминанта; ФС - фагосома; ЛС - лизосома; ЛФ - лизосомальные ферменты; ФЛ - фаголизосома; ПАГ - процессированный антиген; Г-II - антиген гистосовместимости II класса (МНС II); Fc - рецептор для Fc-фрагмента молекулы иммуноглобулина; С1, С3а, С5а - рецепторы для компонентов комплемента; γ -ИФН - рецептор для γ -МФН; С - секреция компонентов комплемента; ПР - секреция перекисных радикалов; ИЛД-1 - секреция; ФНО - секреция фактора некроза опухолей; Сф - секреция ферментов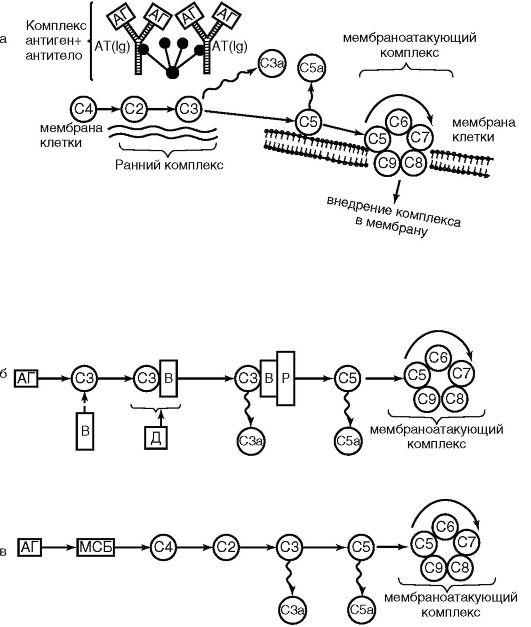 Рис. 9.3. Пути активации комплемента: классический (а); альтернативный (б); лектиновый (в); С1-С9 - компоненты комплемента; АГ - антиген; АТ - антитело; ВиД - протеины; Р - пропердин; МСБ - маннозосвязывающий белок
Рис. 9.3. Пути активации комплемента: классический (а); альтернативный (б); лектиновый (в); С1-С9 - компоненты комплемента; АГ - антиген; АТ - антитело; ВиД - протеины; Р - пропердин; МСБ - маннозосвязывающий белок