|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Обеспечивающие жизнедеятельность клетокСтр 1 из 12Следующая ⇒
Рисунок 1. Параметры внутренней среды, Обеспечивающие жизнедеятельность клеток
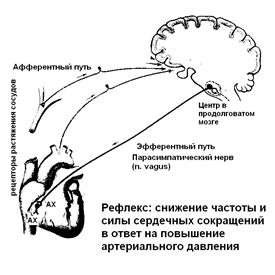
Среди параметров внутренней среды есть жесткие константы – их отклонение от нормальных значений грозит гибелью клеткам - это, например, рН среды, содержание кислорода и углекислого газа, осмотическая концентрация, концентрация натрия и калия. Есть более пластичные константы, их изменение не так быстро приводит к гибели клеток, хотя и нарушает их метаболизм – концентрация глюкозы, белков в плазме крови. Параметров внутренней среды множество, каждый из них имеет определенное значение для сохранности и функционирования клеток и работы ферментов. Величина определенных параметров является результатом регуляции. К результатам относятся жизненно важные метаболические показатели внутренней среды организма, такие как артериальное давление, рН крови, уровень глюкозы, осмотическое давление крови и др. В живом организме сочетаются два принципа регуляции: регуляция по принципу отклонения и регуляция по принципу возмущения. В первом случае – регуляция по принципу отклонения - деятельность регуляторных систем начинается, если какой либо из параметров внутренней среды отклоняется от нормальной величины. Передача информации об отклонении включает регуляторную систему, достижение результата – приведение параметра к норме - является сигналом для прекращения деятельности системы регуляции. В этом случае работают совершенно различные регуляторные системы для приведения к норме разных параметров внутренней среды, например, изменение артериального давления регулируется путем изменения частоты и силы сердечных сокращений и просвета сосудов, а регуляция осмотической концентрации внутренней среды путем изменения количества выведенной из организма воды.. В случае возникновения сигнала опасности, появления угрозы жизни организма включается регуляция по принципу возмущения – это перестройка деятельности всего организма, направленная в первую очередь на выживание. Такой вариант регуляции всегда стандартный и начинается с повышения активности симпатической нервной системы. Нервные и гуморальные механизмы в данном случае работают с опережением, «на всякий случай» готовят организм к активной деятельности. Сравнительная характеристика нервной и гуморальной регуляции
РЕФЛЕКС - ответная реакция организма на раздражение рецепторов, которая заключается в возникновении, изменении или прекращении функциональной активности органов, тканей или целостного организма и осуществляется при обязательном участии центральной нервной системы. Структурной основой рефлекса является рефлекторная дуга, в состав которой входит несколько обязательных компонентов, или звеньев, каждое из которых выполняет собственную функцию. И.П. Павловым были сформулированы основные принципы рефлекторной деятельности: детерминизма, анализа и синтеза и структурности. Принцип структурности понятен из названия - абсолютно необходимым условием для осуществления рефлекса является структурная и функциональная целостность всех звеньев рефлекторной дуги. Принцип детерминизма или причинности заключается в том, что для осуществления любого рефлекса нужна причина, следовательно, действие раздражителя. Рефлекторная реакция осуществляется на основе анализа входящей информации, которая поступает, как правило, из нескольких источников и синтеза - принятия решения на основании этой информации. Анализ и синтез осуществляется и на уровне рецепторов, и в ЦНС.
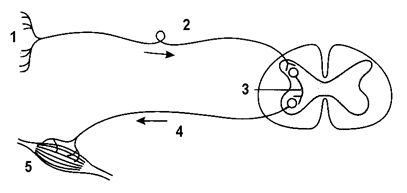
РЕФЛЕКТОРНАЯ ДУГА - совокупность образований, необходимых для осуществления рефлекса. Любая рефлекторная дуга состоит из пяти обязательных звеньев, которые представлены на рисунке 1: рецептора -1, афферентного звена -2, центрального звена -3, эфферентного звена -4 и эффектора -5. На рисунке 1 центральным звеном соматической рефлекторной дуги являются передние рога серого вещества спинного мозаг. Центрами рефлекторных дуг могут быть любые отделы ЦНС.
Рисунок 2. Структура соматической рефлекторной дуги Рисунок 3. Виды рецепторов Классификация рецепторов по модальности
Рецепторы воспринимают определенный вид раздражения (тепло, холод, давление, свет, колебания, растяжение и т.д.) и трансформируют в нервный импульс, который и передается в ЦНС. Задача рецепторов – перевести разнообразные раздражения на язык нервной системы: нервные импульсы, передающиеся с различной частотой. Скопление рецепторов, раздражение которых вызывает строго определенные рефлексы называют РЕЦЕПТИВНЫМИ ПОЛЯМИ или РЕФЛЕКСОГЕННЫМИ ЗОНАМИ. По расположению рецепторов можно выделить экстеро-, интерорецептивные и проприорецептивные, рефлексы – рефлексы, которые возникают при раздражении рецепторов, расположенных на поверхности тела, во внутренних органах или скелетных мышцах. Например, одергивание руки при ожоге – экстероцептивный рефлекс, коленный рефлекс, который мы наблюдаем в кабинете невропатолога в ответ на удар молоточком – проприоцептивный.
АФФЕРЕНТНОЕ ЗВЕНО РЕФЛЕКТОРНОЙ ДУГИ представлено нейронами, отростки которых связывают рецепторы с нервными центрами. Это звено обеспечивает центростремительное проведение возбуждения от рецепторов в структуры Ц Н С. Большинство афферентных нервных волокон, несущих информацию от тактильных, температурных, болевых и механорецепторов относятся к группе А и проводят возбуждение от рецептивных полей в нервные центры со скоростью 80 - 120 м/сек. НЕРВНЫЕ ЦЕНТРЫ - совокупность нервных клеток, " ансамбль" нейронов, которые включаются в регуляцию определенной функции или в осуществление конкретного рефлекторного акта. Свойства нервных центров мы подробно разберем позднее, а пока отметим лишь функциональное значение центров. В нервном центре происходит интеграция всей поступающей по афферентным путям информации, а из центра поступает команда к действию. Центры рефлексов могут быть расположены в любом отделе ЦНС. В зависимости от расположения нервных центров различают рефлексы СПИНАЛЬНЫЕ - нервные центры находятся в сегментах спинного мозга, БУЛЬБАРНЫЕ - в продолговатом мозге, МЕЗЭНЦЕФАЛЬНЫЕ - в структурах среднего мозга, КОРТИКАЛЬНЫЕ - в различных областях коры большого мозга. Например, одергивание руки при ожоге – спинальный рефлекс. В соответствии с выполняемой функцией среди нервных центров можно выделить чувствительные центры, центры вегетативных функций, двигательные центры. Кроме того, можно выделить структурно организованные центры, которые регулируют определенную функцию: сосудодвигательный центр, дыхательный центр, центр слюноотделения. ЭФФЕРЕНТНОЕ ЗВЕНО РЕФЛЕКТОРНОЙ ДУГИ – путь, по которому возбуждение распространяется центробежно, от ЦНС на периферию, к рабочим органам. ЭФФЕКТОРЫ - специализированные органы и ткани, реагирующие на импульсы, поступающие к ним по эфферентным путям. Эффектор это рабочий орган, действия которого должны привести к результату – восстановлению параметра внутренней среды в случае его отклонения и прекращению раздражения рецепторов. Например, одергивание руки при ожоге приводит к прекращению раздражения терморецепторов; при снижении артериального давления сердце, как рабочий орган увеличивает сердечный выброс и восстанавливает важный параметр – АД. К эффекторам относятся мышцы, внутренние органы, железы. Основные функции эффекторов: 1) деятельность по восстановлению параметров внутренней среды организма (регуляция по отклонению) и 2) деятельность, направленная на сохранение целостности организма (регуляция по возмущению) По виду эффектора рефлексы делятся на СОМАТИЧЕСКИЕ и ВЕГЕТАТИВНЫЕ. Эффекторами соматических рефлексов являются поперечнополосатые скелетные мышцы, с помощью которых поддерживается поза тела и выполняются произвольные движения. Например, одергивание руки при ожоге – соматический рефлекс. Характерной особенностью соматических рефлексов является возможность произвольного управления движением скелетных мышц. Эффекторами вегетативных рефлексов являются все внутренние органы, сосуды, железы внешней и внутренней секреции, произвольное управление этими эффекторами невозможно. Вегетативная нервная система - часть нервной системы, иннервирующая внутренние органы, кровеносные сосуды, железы, гладкую и отчасти поперечно-полосатую мускулатуру. В вегетативной нервной системе различают симпатический и парасимпатический отделы. Эти отделы отличаются по характеру влияний на внутренние органы. Симпатический отдел вегетативной нервной системы – это система активности, готовности к деятельности, во время которой интенсивно работают сердечно-сосудистая, дыхательная системы, повышается активность ЦНС, расходуются метаболические запасы организма. Парасимпатический отдел вегетативной нервной системы– система покоя, восстановления запасов.
Рисунок 4. Функциональное значение отделов Анатомической особенностью вегетативной нервной системы является то, что аксоны ее центральных нейронов направляются не прямо к органу, а контактируют с нервными клетками, образующими периферические нервные ганглии. Ганглии парасимпатического отдела расположены либо в стенах органа (интрамурально) или рядом (параорганно). В симпатическом отделе ганглии расположены в симпатическом стволе, чревном и брызжеечном сплетениях. Таким образом, вегетативная нервная система отличается от соматической эфферентным звеном, в котором выделяют преганглионарное и постганглионарное волокна. Вегетативная нервная система отличается от соматической и расположением центров рефлекторных дуг. Центры симпатических и парасимпатических рефлекторных дуг расположены в различных отделах ЦНС. Парасимпатические центры находятся в среднем и продолговатом мозге и крестцовом отделе спинного мозга, симпатические расположены в боковых рогах грудного и поясничного отделов спинного мозга, однако и те и другие связаны с вышележащими структурами головного мозга. На приведенных ниже рисунках представлены схемы соматического и вегетативных рефлексов.
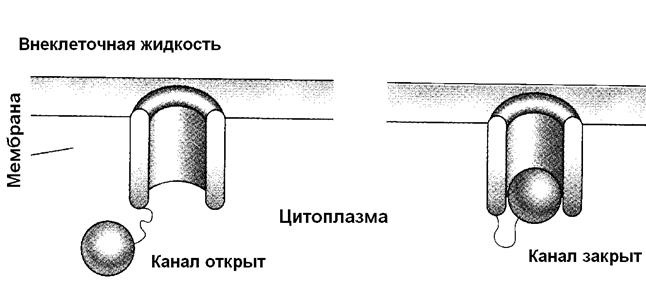
Вегетативная нервная система оказывает три вида влияний на работу органов: Пусковое влияние возбуждает орган, который работает непостоянно, например, потовые железы начинают выделять свой секрет под влиянием симпатической нервной системы при повышения температуры окружающей среды. Корригирующее влияние - усиление или ослабление деятельности постоянно работающих органов. Например, увеличение или уменьшение частоты и силы сердечных сокращений под действием симпатических или блуждающего нервов. Адаптационно-трофическое влияние вегетативной нервной системы заключается во включении в регуляцию деятельности организма систем обмена веществ, обеспечивающих координированное функционирование органов и систем при нагрузке, приспособление к изменяющимся внешним условиям, восстановление после физической нагрузки, при выздоровлении. Например, увеличение темпа метаболических процессов в интенсивно работающем миокарде. Рисунок 5. Функциональная организация центральной и периферической нервной системы. Разность потенциалов между внутренней и наружной поверхностью клетки называется мембранный потенциал покоя (МПП), для разных клеток он колеблется от – 60 до –90 милливольт. Величина внутриклеточного электрического потенциала, например, у нервных клеток, составляет приблизительно -70 мв. Возбудимые клетки способны реагировать на раздражение и возбуждаться потому, что в исходном состоянии они поляризованы, т.е. обладают разностью потенциалов между внутренней стороной мембраны, которая заряжена отрицательно, и окружающей средой. Клетка, не имеющая такого потенциала, не способна ответить на раздражение, поэтому во всех возбудимых клетках постоянно протекают процессы, поддерживающие этот потенциал. Таким образом – мембранный потенциал покоя – основа возбудимости клетки. Клеточная мембрана, отделяющая клетку от окружающего пространства, построена из двух слоев жироподобных молекул, которые называются фосфолипиды. Есть белки, которые пронизывают мембрану насквозь, такие белки называются интегральными. Интегральные белки могут образовывать в мембране пору (отверстие, заполненное водой) для транспорта ионов. Ионные каналы Ионные каналы представлены интегральными белками мембраны. Эти белки способны, при определенных воздействиях, изменять свою конформацию (форму и свойства) таким образом, что пора, через которую может пройти какой-либо ион открывается или закрывается. Известны натриевые, калиевые, кальциевые, хлорные каналы, иногда канал может пропускать два иона, например известны натрий – кальциевые каналы. Через ионные каналы осуществляется только пассивный транспорт ионов. Это значит, что для перемещения иона необходим не только открытый канал, но и градиент концентрации для этого иона. В этом случае, будет движение иона по градиенту концентрации – из области с большей концентрацией в область с меньшей концентрацией. Необходимо помнить, что мы говорим об ионах – заряженных частицах, транспорт которых обусловлен еще и зарядом. Возможны ситуации, когда движение по градиенту концентрации может быть направлено в одну сторону, а существующие заряды противодействуют этому переносу. Ионные каналы обладают двумя важнейшими свойствами: 1) избирательностью (селективностью) по отношению к определенным ионам и 2) способностью открываться (активироваться) и закрываться. При активации канал открывается и пропускает ионы (рис. 8). Таким образом, в комплекс интегральных белков, формирующих канал, должны обязательно входить два элемента: структуры, распознающие «свой» ион и способные его пропустить, и структуры, которые позволяют узнать – когда пропускать этот ион. Селективность канала определяется теми белками, которые его образуют, «свой» ион распознается по размерам и заряду. Активация каналов возможна несколькими путями. Во-первых, каналы могут открываться и закрываться при изменении потенциала мембраны. Изменение заряда приводит к изменению конформации белковых молекул, и канал становится проницаемым для иона. Для изменения свойств канала достаточно ничтожного колебания потенциала мембраны. Такие каналы называются потенциал-зависимые (или электроуправляемые). Во-вторых, каналы могут быть частью сложного белкового комплекса, который называется мембранный рецептор. В этом случае изменение свойств канала обусловлено конформационнй перестройкой белков, которая происходит в результате взаимодействия рецептора с биологически активным веществом (гормоном, медиатором). Такие каналы называются хемозависимые (или рецептор-управляемые ). Кроме того, каналы могут открываться при механическом воздействии – давлении, растяжении (рис.9). Механизм, который обеспечивает активацию, называется воротами канала. По скорости, с которой открываются и закрываются каналы их можно разделить на быстрые и медленные. Большинство каналов (калиевые, кальциевые, хлорные) могут находиться в двух состояниях: открытом и закрытом. В работе натриевых каналов есть некоторые особенности. Этим каналам, как и калиевым, кальциевым, хлорным свойственно находиться или в открытом, или в закрытом состоянии, однако, натриевый канал может быть и инактивирован, этот состояние, в котором канал закрыт и не может быть открыт никаким воздействием (рис.10).
Рисунок 8. Состояния ионных каналов
Рисунок 9. Пример работы рецептор-управляемого канала. АЦХ – ацетилхолин. Взаимодействие молекулы АЦХ с мембранным рецептором изменяет конформацию воротного белка таким образом, что канал начинает пропускать ионы.
Рисунок 10 Пример потенциал-зависимого канала Рисунок 19. Структура синапса.
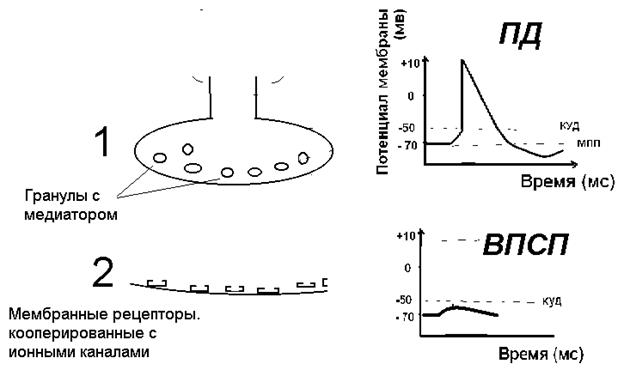
Пресинаптическая мембрана терминали аксона представлена, как правило, утолщением, в котором содержится множество гранул диаметром от 20 - 40 до 120 нм. С помощью гистохимических методов было установлено, что в гранулах содержится химические вещества, различные для различных нейронов, эти химические вещества были названы медиаторами – химическими «посредниками» в передаче электрического импульса с пресинаптической к постсинаптической мембране. Медиаторы могут быть: — биогенными аминами: норадреналин, дофамин, гистамин, серотонин; — аминокислотами: глутаминовая кислота, глицин; — олигопептидами: вещество Р, энкефалины, эндорфины; — метаболитами: аденозин, АТФ, АДФ; — широко распространен и в центральных структурах, и на периферии медиатор ацетилхолин.
Химические медиаторы являются веществами, синтезируемыми в цитоплазме нейронов. Нейроны могут: 1) синтезировать один медиатор; 2) транспортировать медиатор по аксону с помощью микротрубочек; 3) упаковывать медиатор в гранулы; 4) хранить эти гранулы в терминалях аксонов; 5) освобождать медиатор при возбуждении; 6) захватывать назад в терминаль аксона медиатор или его фрагменты; 7) восстановить гранулы и вновь заполнить их медиатором.
Медиатор выделяется в синаптическую щель размером 10-50 мкм, и взаимодействует с мембранным рецептором постсинаптической мембраны. В синаптической щели возможно разрушение медиатора специфическими для каждого медиатора ферментами, метаболиты возвращаются в нейрон для ресинтеза. Такими ферментами могут быть ацетилхолинэстераза – фермент для разрушения ацетилхолина, моноаминооксидаза (МАО) – для разрушения норадреналина или дофамина. Возможен обратный захват не разрушенного медиатора терминалью аксона. Постсинаптическая мембрана имеет мембранные рецепторы – сложные комплексы интегральных белков, пронизывающих клеточную мембрану, как правило, состоящие из нескольких субъединиц. В мембранных рецепторах одна часть способна распознавать собственный медиатор и связываться с ним, а вторая – представлена или ионным каналом, или системой специальных мембранных ферментов, которые называются внутриклеточными посредниками. Если взаимодействие медиатора с рецептором приводит к активации ионных каналов, а результатом является изменение потенциала мембраны, то такие рецепторы называются ионотропными. Если запускается система вторичных внутриклеточных посредников, происходит изменение активности ферментов и темпа метаболических процессов (гликолиз, липолиз), то такие рецепторы называются метаботропными. Вернитесь к рисунку 9, на котором представлена активация натриевого канала при взаимодействии ацетилхолина с холинорецептором – это пример ионотропного рецептора.
Для выделения медиатора необходимо сочетание двух событий: 1) волна деполяризации (ПД), достигшая терминали аксона, эта волна обусловлена активацией быстрых натриевых каналов и поступлением ионов натрия в клетку и 2) открытие медленных, потенциал-зависимых кальциевых каналов пресинаптической мембраны. Поступление кальция в клетку (по градиенту концентра
Рисунок 20. Выделение медиатора и взаимодействие с рецепторами Этапы выделения медиатора: I) Активация натриевых каналов, ток натрия в терминаль аксона (в клетку) по градиенту концентрации и деполяризация пресинаптической мембраны II) Активация потенциалзависимых медленных кальциевых каналов и поступление ионов кальция в клетку по градиенту концентрации. Ионы кальция обеспечивают освобождение медиатора. III) Диффузия медиатора к постсинаптической мембране и взаимодействие со специфическими мембранными рецепторами. IV) Судьба медиатора в синаптической щели 1)Возвращение метаболитов в терминаль аксона 2)Разрушение медиатора специфическими ферментами 3)Обратный захват медиатора
Результат взаимодействия медиатора с мембранными рецепторами постсинаптической мембраны приводит к изменениям проницаемости этой мембраны для ионов путем активации или инактивации ионных каналов. На постсинаптической мембране могут открываться (или закрываться) натриевые, кальциевые, калиевые или хлорные каналы. Результатом этого процесса будет движение ионов по градиенту концентрации и изменение мембранного потенциала постсинаптической мембраны. В случае активации натриевых каналов ионы натрия по градиенту концентрации будут поступать в клетку и произойдет деполяризация постсинаптической мембраны. Если деполяризующий ток ионов велик (натриевые каналы), на постсинаптической мембране возникает импульс возбуждения – потенциал действия. Чаще же на постсинаптической мембране возникает лишь незначительная, не достигающая порогового уровня деполяризация, локальный ответ (ЛО). В зависимости от расположения синапса этот ЛО называется или ВПСП (возбуждающий постсинаптический потенциал в центральных синапсах) или ПКП (потенциал концевой пластинки в нервномышечных синапсах).
Рисунок 22. Изменение потенциалов пресинаптической и постсинаптической мембраны 1 – пресинаптическая мембрана, 2 – постсинаптическая мембрана
Гибель нейронов Удивительной особенностью развития нервной системы является то, что многим ее нейронам суждено погибнуть во время развития. У беспозвоночных интенсивная гибель нейронов сопровождается стремительными изменениями, происходящими во время метаморфоза, и регулируется гормонами. Однако в развивающейся ЦНС как позвоночных, так и беспозвоночных гибель клеток происходит также и в отсутствие значительных морфологических изменений. Эксперименты, выполненные Гамбургером и Леви-Монтальчини, впервые выявили запрограммированную гибель нейронов эмбрионов позвоночных, а также показали, что степень гибели клеток может зависеть от размеров ткани-мишени. Они показали, например, что в развивающейся конечности в то время, когда формируются первые синаптические связи с миофибриллами, от 40 до 70 % мотонейронов, которые послали аксоны в эту конечность, погибают. Имплантация дополнительной конечности уменьшает долю погибающих мотонейронов, а удаление одного из зачатков конечностей приводит к увеличению числа гибнущих клеток, что дает основания для предположения, что мотонейроны конкурируют за некие трофические вещества, выделяемые тканью-мишенью. Избыточная продукция нейронов, после которой следует период гибели клеток, является характерной чертой развития нервной системы позвоночных. Некоторые из нейронов, которые погибают, возможно, не смогли образовать синапсов, или образовали их с неподходящей клеткой-мишенью. В подобных случаях гибель клеток связана с особенностью иннервации. Однако большинство клеток, которые гибнут, достигают и иннервируют правильные клетки-мишени. Таким образом, гибель клеток является основным механизмом, который поддерживает равновесие между количеством нейронов и их мишеней. Неожиданной находкой явилось то, что ингибиторы образования м-РНК или синтеза белка в нейронах предотвращали их гибель. Результаты этих и более поздних экспериментов показали, что гибель нейронов типично происходит путем апоптоза. Апоптоз - это процесс, который активирует внутреннюю «суицидальную» программу клетки, которая приводит к упорядоченному (запрограммированному) распаду ДНК и белков в клетке. Существенно, что для этого необходим предварительный синтез протеолитических ферментов или их активаторов. Развитие синапсов Многие исследования функции и формирования синапсов были сфокусированы на нейромышечных синапсах - синапсах между двигательными нейронами и скелетными мышцами, из-за их относительной доступности и простоты. Они показали, что формирование синапсов нуждается в сложном обмене информацией между нейроном и его мишенью, это делает возможным взаимное распознавание и инициирует сигнальные каскады, которые ведут к дифференцировке и формированию пресинаптических и постсинаптических участков мембраны со всем комплексом структур, необходимых для передачи импульса возбуждения. Полученная информация с достоверностью показала, что принципиальной разницы между процессами формирования нервномышечных и центральных синапсов нет. Следовательно нейромышечные синапсы могут рассматриваться как хорошая модель синапсов. Во время развития аксоны от двигательных нейронов выходят из спинного мозга и проводятся к своим мишеням с помощью клеточных и молекулярных сигналов, которые могут или привлекать или отталкивать их. Оказавшись в непосредственно близи к мишени, аксон замедляет рост и в конечном итоге останавливается. Кончик аксона контактирует с мишенью и тем самым инициируется дифференцировка пресинаптических окончаний. Комплекс последовательных событий запускается уже при первом контакте врастающего аксона с растущей мышцей. Когда аксон впервые достигает мышечной массы, миобласты всё ещё сливаются вместе, заканчивая формирование первичных и вторичных миобибрилл. Одним из наиболее важных событий, которое следует сразу после контакта аксона с дифференцирующимся мышечным волокном, является быстрый сбор (в течение минут) до этого диспергированных ацетилхолиновых рецепторов под формирующимся нервным окончанием. Эта быстрая агрегация холинорецепторов является характерной особенностью образования синапса. Установлено, что синаптическая базальная мембрана содержит в виде различных белков всю информацию, необходимую для запуска и управления всеми аспектами образования синапса. Одним из белков, запускающих агрегацию холинорецепторов, является агрин. Этот белок синтезируется и в мышцах и нервах, секретируется и проникает в базальную мембрану. Описано несколько форм агрина, однако только тот, который синтезируется в двигательных нейронах эффективно собирает холинорецепторы постсинаптической мембраны точно на вершинах складок постсинаптической мембраны в непосредственной близости от места выделения медиатора, так же собирает в комплексы натриевые каналы, активирует синтез холинэстеразы.
НЕОНАТАЛЬНЫЙ ПЕРИОД Созревание ЦНС. В неонатальном периоде отмечаются дальнейший рост нервной ткани, усиление процессов миелинизации нервных волокон, дифференцировки нейрофибрилл, совершенствование механизмов проницаемости клеточных мембран, повышение возбудимости нейронов, развитие их шипикового аппарата, установление ассоциативных связей, что в итоге приводит к постепенному совершенствованию базовых нейродинамических процессов. У новорожденных спинной мозг, структуры ствола мозга, бледное ядро, зрительный бугор развиты в целом хорошо. Красное ядро, руброспинальный путь миелинизированы. Двигательные области коры большого мозга (поля 4 и 6 по Бродману) к моменту рождения развиты недостаточно. В спинном мозге, стволе и гипоталамусе у новорожденных обнаруживаются ацетилхолин, гамма-аминомасляная кислота, серотонин, норадреналин, дофамин. Содержание медиаторов низкое и составляет 10—50 % от такового у взрослых. В постсинаптических мембранах нейронов к моменту рождения появляются специфические для перечисленных медиаторов рецепторы. Созревание структур ЦНС усиливается гормонами щитовидной железы. Стимулирующая роль в ходе созревания и функционального становления ЦНС отводится афферентным потокам импульсов, поступающих в структуры мозга из внешней среды. Спонтанные периодические движения новорожденного беспорядочны, хаотичны, в них участвуют конечности, голова и туловище. Тем не менее, наблюдаются и координированные ритмические сгибания и разгибания. Периоды двигательной активности отчетливо преобладают над периодами полного покоя. Для проснувшегося новорожденного характерны пространственно ориентированные движения головы, направленные на поиск материнской груди, полноценные сосательные движения. Мышечный тонус у новорожденных поддерживается импульсами, идущими от проприорецепторов, кожных терморецепторов и даже рецепторов растяжения легких, активирующихся при вдохе. Для новорожденного, как и для плода, характерна ортотоническая поза как следствие некоторой гипертензии мышц-сгибателей. Характеристика рефлексов. Отличительными особенностями рефлексов новорожденных являются генерализованный характер их проявления и обширность рефлексогенной зоны вызова того или иного рефлекса. Эти свойства рефлексов объясняются, во-первых, отсутствием над ними контроля со стороны головного мозга, во-вторых, относительно облегченной иррадиацией процесса возбуждения в ЦНС. Причиной более выраженной иррадиации возбуждения является слабость процессов торможения. С возрастом рефлексы становятся более совершенными: генерализованность уменьшается, рефлексогенные зоны рефлексов сужаются. Так, в возрасте 1—5 дней рефлексогенной зоной сосательного рефлекса являются губы и кожа всего лица; 6—10 дней — губы и кожа вокруг рта; 15 дней — только губы. Ряд рефлексов новорожденного постепенно исчезает, но многие из них подвергаются лишь угнетению в результате развивающихся тормозных влияний со стороны вышележащих отделов мозга, в особенности коры большого мозга, на нижележащие центры. Всю совокупность рефлексов новорожденного целесообразно разделить на пять групп. 1. Пищевые: сосательный и глотательный рефлексы появляются при механическом, тепловом и вкусовом раздражении рецепторов ротовой и околоротовой областей. Так, если вложить ребенку в рот соску, он начинает совершать активные сосательные движения. Сосательный рефлекс исчезает к концу первого года жизни. 2. Защитные: мигательный рефлекс — мигание при освещении глаз светом или при раздражении поверхности носа, век, ресниц, роговой оболочки глаз; зрачковый рефлекс — уменьшение диаметра зрачка при освещении; рефлекс отдергивания конечности в ответ на болевое раздражение. 3. Двигательные. • Хватательный рефлекс (рефлекс Робинсона) — схватывание и прочное удерживание предмета (пальца, карандаша, игрушки) при прикосновении им к ладони — исчезает на 2—4-м месяцах. • Рефлекс обхватывания (рефлекс Моро) — отведение рук в стороны и разгибание пальцев с последующим возвращением рук в исходное положение. Для вызова рефлекса ребенка, находящегося на руках у врача, резко опускают на 20 см и затем поднимают до исходного уровня. Рефлекс можно вызвать при ударе по поверхности, на которой лежит ребенок, а также при быстром подъеме его из положения на спине. Рефлекс исчезает после 4 месяца жизни. • Подошвенный рефлекс (рефлекс Бабинского) — изолированное тыльное разгибание большого пальца и подошвенное сгибание (иногда веерообразное расхождение) остальных при раздражении подошвы по наружному краю стопы от пятки к пальцам; исчезает после первого года жизни. • Коленный рефлекс — сгибание (у взрослых разгибание) в коленном суставе при ударе по сухожилию четырехглавой мышцы ниже коленной чашечки. Сгибание у новорожденных связано с преобладанием у них тонуса мышц-сгибателей; заменяется разгибательным рефлексом на 2-м месяце. • Хоботковый рефлекс - выпячивание губ хоботком в результате сокращения круговой мышцы рта при легком ударе пальцем по губам ребенка или поколачива-нии кожи вокруг рта на уровне десен; исчезает к концу первого полугодия жизни. • Поисковый рефлекс (поиск груди матери) — опускание губ, отклонение языка и поворот головы в сторону раздражителя при поглаживании кожи в области угла рта. Рефлекс ярче выражен у голодного ребенка; исчезает к концу 1-го года жизни. Популярное:
|
Последнее изменение этой страницы: 2016-07-14; Просмотров: 755; Нарушение авторского права страницы





 ции) стимулирует освобождение гранул с медиатором. Гранулы выделяются путем экзоцитоза, поэтому медиатор появляется в синаптической щели не отдельными молекулами, а квантами, примерно из нескольких тысяч молекул.
ции) стимулирует освобождение гранул с медиатором. Гранулы выделяются путем экзоцитоза, поэтому медиатор появляется в синаптической щели не отдельными молекулами, а квантами, примерно из нескольких тысяч молекул.