|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
В потенциал-зависимом натриевом канале имеются активационные и инактивационные ворота (заслонки). Активационные и инактивационные заслонки меняют конформацию при различном мембранном потенциале.
Механизм работы Na+/K+ насоса как пример активного транспорта, использующего энергию АТФ Для переноса ионов против концентрационного градиента Na+/K+ насос (или Na+/K+ АТФ-аза) это тоже, как и ионные каналы комплекс интегральных мембранных белков, которые могут не просто открыть путь для перехода иона по градиентуконцентрации, а активно перемещать ионы против градиента концентрации. Механизм работы насоса представлен на рисунке 11.
5. Происходит перенос калия, освобождение его во внутриклеточную среду и присоединение молекулы АТФ – насос вернулся в конформацию Е1, вновь приобрел сродство к ионам натрия и включается в новый цикл. Рисунок 11. Механизм работы Na+/K+ насоса
Обратите внимание на то, что Na+/K+насос переносит 3 иона натрия из клетки в обмен на 2 иона калия. Поэтому насос является электрогенным: суммарно за один цикл из клетки удаляется один положительный заряд.
Концентрация ионов внутри и вне клетки
Итак, есть два факта, которые необходимо учесть, чтобы понять механизмы, поддерживающие мембранный потенциал покоя. 1. Концентрация ионов калия в клетке значительно выше, чем во внеклеточной среде. 2. Мембрана в покое избирательно проницаема для ионов К+, а для Nа+ проницаемость мембраны в покое незначительна. Если принять проницаемость для калия за 1, то проницаемость для натрия в покое составит лишь 0, 04. Следовательно, существует постоянный поток ионов К+ из цитоплазмы по градиенту концентрации. Калиевый ток из цитоплазмы создает относительный дефицит положительных зарядов на внутренней поверхности, для анионов клеточная мембрана непроницаема, в результате цитоплазма клетки оказывается заряженной отрицательно по отношению к окружающей клетку среде. Эта разность потенциалов между клеткой и внеклеточным пространством называется мембранным потенциалом покоя (МПП). В покое клеточная мембрана поляризована – обладает различным зарядом на внутренней и внешней стороне. Если этот заряд уменьшается, говорят о деполяризации клетки, а если увеличивается – о гиперполяризации. Возбудимость тканей может изменяться, поэтому необходимо знать причины изменения возбудимости и результаты изменения, т.е. к чему это может привести. Например, возбудимостьнервных клеток повышается при накоплении в них ионов кальция (положительный ион), в результате действия медиатора симпатичекой нервной системы - норадреналина. Такое повышение возбудимости приведет к тому, что нейроны будут возбуждаться в ответ на малейшее раздражении. Возбудимость повышается и при накоплении в них ионов натрия. Вместе с тем, зная, что ионы калия из крови будут обязательно доставлены в клетку с помощью Na/К насоса, а повышение концентрации ионов калия в клетке снижает ее возбудимость, можно использовать эти знания для воздействия на структуры ЦНС. Причиной изменения возбудимости, кроме нарушения внутриклеточной концентрации ионов, могут быть метаболические процессы, связанные, например, с нарушением синтеза АТФ, в этом случае возбудимость может снизиться, т.к. нарушатся процессы реполяризации клеток. Кроме того, ионные каналы это белки, которые чувствительны к действию различных токсинов (ядов), поэтому следует помнить о том, что возбудимость у лиц, злоупотребляющих, например, алкоголем отличается от нормальной В химических синапсах возбуждение передается с помощью химического вещества - МЕДИАТОРА. В структуре синапса принято выделять пресинаптическую мембрану, образованную терминалью аксона, синаптическую щель и постсинаптическую мембрану, каждая структура выполняет свою функцию (рис.19).
Рисунок 19. Структура синапса.
Пресинаптическая мембрана терминали аксона представлена, как правило, утолщением, в котором содержится множество гранул диаметром от 20 - 40 до 120 нм. С помощью гистохимических методов было установлено, что в гранулах содержится химические вещества, различные для различных нейронов, эти химические вещества были названы медиаторами – химическими «посредниками» в передаче электрического импульса с пресинаптической к постсинаптической мембране. Медиаторы могут быть: — биогенными аминами: норадреналин, дофамин, гистамин, серотонин; — аминокислотами: глутаминовая кислота, глицин; — олигопептидами: вещество Р, энкефалины, эндорфины; — метаболитами: аденозин, АТФ, АДФ; — широко распространен и в центральных структурах, и на периферии медиатор ацетилхолин.
Химические медиаторы являются веществами, синтезируемыми в цитоплазме нейронов. Нейроны могут: 1) синтезировать один медиатор; 2) транспортировать медиатор по аксону с помощью микротрубочек; 3) упаковывать медиатор в гранулы; 4) хранить эти гранулы в терминалях аксонов; 5) освобождать медиатор при возбуждении; 6) захватывать назад в терминаль аксона медиатор или его фрагменты; 7) восстановить гранулы и вновь заполнить их медиатором.
Медиатор выделяется в синаптическую щель размером 10-50 мкм, и взаимодействует с мембранным рецептором постсинаптической мембраны. В синаптической щели возможно разрушение медиатора специфическими для каждого медиатора ферментами, метаболиты возвращаются в нейрон для ресинтеза. Такими ферментами могут быть ацетилхолинэстераза – фермент для разрушения ацетилхолина, моноаминооксидаза (МАО) – для разрушения норадреналина или дофамина. Возможен обратный захват не разрушенного медиатора терминалью аксона. Постсинаптическая мембрана имеет мембранные рецепторы – сложные комплексы интегральных белков, пронизывающих клеточную мембрану, как правило, состоящие из нескольких субъединиц. В мембранных рецепторах одна часть способна распознавать собственный медиатор и связываться с ним, а вторая – представлена или ионным каналом, или системой специальных мембранных ферментов, которые называются внутриклеточными посредниками. Если взаимодействие медиатора с рецептором приводит к активации ионных каналов, а результатом является изменение потенциала мембраны, то такие рецепторы называются ионотропными. Если запускается система вторичных внутриклеточных посредников, происходит изменение активности ферментов и темпа метаболических процессов (гликолиз, липолиз), то такие рецепторы называются метаботропными. Вернитесь к рисунку 9, на котором представлена активация натриевого канала при взаимодействии ацетилхолина с холинорецептором – это пример ионотропного рецептора.
Для выделения медиатора необходимо сочетание двух событий: 1) волна деполяризации (ПД), достигшая терминали аксона, эта волна обусловлена активацией быстрых натриевых каналов и поступлением ионов натрия в клетку и 2) открытие медленных, потенциал-зависимых кальциевых каналов пресинаптической мембраны. Поступление кальция в клетку (по градиенту концентра
Рисунок 20. Выделение медиатора и взаимодействие с рецепторами Этапы выделения медиатора: I) Активация натриевых каналов, ток натрия в терминаль аксона (в клетку) по градиенту концентрации и деполяризация пресинаптической мембраны II) Активация потенциалзависимых медленных кальциевых каналов и поступление ионов кальция в клетку по градиенту концентрации. Ионы кальция обеспечивают освобождение медиатора. III) Диффузия медиатора к постсинаптической мембране и взаимодействие со специфическими мембранными рецепторами. IV) Судьба медиатора в синаптической щели 1)Возвращение метаболитов в терминаль аксона 2)Разрушение медиатора специфическими ферментами 3)Обратный захват медиатора
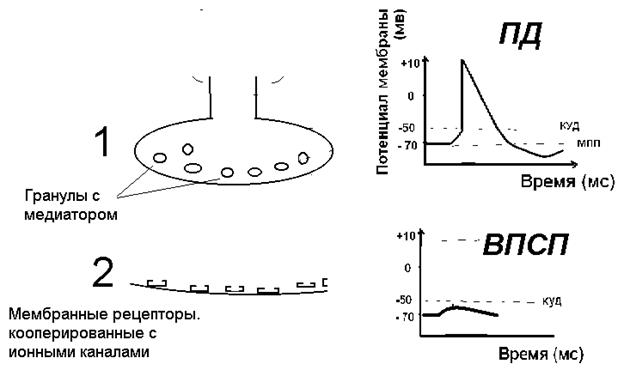
Результат взаимодействия медиатора с мембранными рецепторами постсинаптической мембраны приводит к изменениям проницаемости этой мембраны для ионов путем активации или инактивации ионных каналов. На постсинаптической мембране могут открываться (или закрываться) натриевые, кальциевые, калиевые или хлорные каналы. Результатом этого процесса будет движение ионов по градиенту концентрации и изменение мембранного потенциала постсинаптической мембраны. В случае активации натриевых каналов ионы натрия по градиенту концентрации будут поступать в клетку и произойдет деполяризация постсинаптической мембраны. Если деполяризующий ток ионов велик (натриевые каналы), на постсинаптической мембране возникает импульс возбуждения – потенциал действия. Чаще же на постсинаптической мембране возникает лишь незначительная, не достигающая порогового уровня деполяризация, локальный ответ (ЛО). В зависимости от расположения синапса этот ЛО называется или ВПСП (возбуждающий постсинаптический потенциал в центральных синапсах) или ПКП (потенциал концевой пластинки в нервномышечных синапсах).
Рисунок 22. Изменение потенциалов пресинаптической и постсинаптической мембраны 1 – пресинаптическая мембрана, 2 – постсинаптическая мембрана
Популярное:
|
Последнее изменение этой страницы: 2016-07-14; Просмотров: 1324; Нарушение авторского права страницы


 ции) стимулирует освобождение гранул с медиатором. Гранулы выделяются путем экзоцитоза, поэтому медиатор появляется в синаптической щели не отдельными молекулами, а квантами, примерно из нескольких тысяч молекул.
ции) стимулирует освобождение гранул с медиатором. Гранулы выделяются путем экзоцитоза, поэтому медиатор появляется в синаптической щели не отдельными молекулами, а квантами, примерно из нескольких тысяч молекул.