|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Общение и групповое поведение
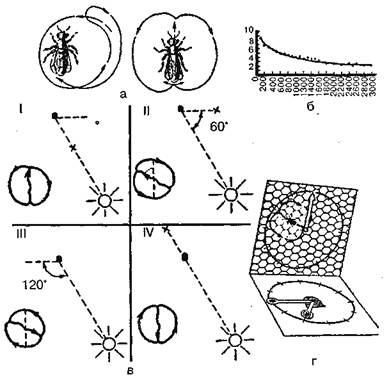
У высших беспозвоночных уже вполном объеме представлена категория поведения, которая, как было показано, только в зачаточной форме обнаруживается у некоторых представителей низших многоклеточных беспозвоночных. Речь идет об общении, которое у высших беспозвоночных проявляется иногда и в очень сложных формах подлинного группового поведения (в отличие от колониальной жизни, свойственной многим низшим беспозвоночным). Напомним, что общение, в точном значении этого термина, появляется только тогда, когда контакты между животными начинают осуществляться посредством специальных сигнальных действий. У головоногих моллюсков и членистоногих, особенно у насекомых, мы находим четкие системы коммуникации с передачей информации по разным сенсорным каналам. Это относится прежде всего к пчелам и муравьям, что, конечно, связано со сложной, высокодифференцированной структурой их сообществ и разделением функций между членами последних (распределение пищедобывательной, защитной и воспроизводительной функций между отдельными особями). У муравьев разные формы химической чувствительности играют, очевидно, большую роль, чем другие виды рецепции, и общение между особями осуществляется прежде всего по этому каналу: личинки муравьев выделяют специфические вещества, побуждающие взрослых рабочих особей кормить их, по запаху обитатели одного муравейника узнают «своих» и «чужих». Даже о том, жив или мертв муравей, его сородичи узнают по специфическим выделениям. Согласование поведения обитателей муравейника происходит путем подачи и приема химических сигналов. Достаточно указать на химические средства муравьев (а также пчел и ос), выделение которых служит сигналом тревоги. Эти вещества выделяются муравьями в случае опасности и быстро распространяются по воздуху. Их восприятие вызывает у других муравьев («солдат») защитную реакцию (готовность к нападению на врага), хотя источник опасности им еще не известен. Поскольку, однако, радиус действия такого сигнала, подаваемого одним муравьем (например, в случае вторжения в муравейник какого-либо насекомого), невелик, то он способен указать и место, откуда угрожает опасность. Чем больше эта опасность, тем больше муравьев выделяют химические сигналы тревоги и тем больше, следовательно, особей призывается на помощь. По другим выделениям, оставляемым на субстрате в виде следа, муравьи узнают, куда, в какую сторону ушел их сородич. К тому же интенсивность такого пахучего следа указывает на количество пищи, обнаруженной в том месте, куда ведет след, тем более что, направляясь к муравейнику, муравьи наносили по пути пахучие метки лишь в тех случаях, когда они находили корм. При этом тем больше оставляется меток, чем больше обнаружено корма. Поскольку эти метки довольно быстро испаряются, вновь идущие по этому следу муравьи будут воспринимать лишь свежие метки, а таких будет все меньше по мере приближения кормового объекта. Таким образом, муравьи в конце концов узнают о том, что пищевые ресурсы в данном месте исчерпаны, и перестанут к нему направляться. У некоторых пчел также существует химический способ передачи информации о нахождении пищи. Этоотносится, прежде всего, к пчелам — из подсемейства Meliponinae, у которых нет «языка танца». Эти широко распространенные в тропиках пчелы не имеют жала, но яростно кусаются. У мелких форм (самая маленькая имеет длину всего 1, 5 миллиметра) нашедшая пищу сборщица только приводит других пчел в возбужденное состояние, беспокойно бегая по сотам, толкая их и время от времени подбегая к летку. Вылетающие же пчелы ориентируются только по запаху цветов. У других видов возбуждение пчел первоначально достигается подобным же образом, но дополнительно сборщица, принесшая взяток, приводит соты толчками в вибрирующее состояние. После этого она возвращается к обнаруженным цветам, а на обратном пути оставляет на траве, ветках или камнях через каждые два-три метра пахучие метки, по которым затем приводит группами других пчел к источнику пищи. Это достигается тем, что пчела-разведчица летает взад и вперед между двумя соседними метками, пока другие пчелы не присоединяются к ней. Следовательно, здесь мы имеем дело с комплексной передачей информации по оптическому, тактильно-вибрационному и, возможно, акустическому каналам, но при этом ведущую роль играют оставляемые сборщицей запахи. У нашей медоносной пчелы также существует химический способ передачи информации, но только как вспомогательный, когда она имеет дело с кормом, лишенным собственного заметного запаха. В этих случаях место нахождения кормового объекта метится секретом из так называемой железы Насонова. Однако основным и наиболее совершенным способом передачи информации о кормовых объектах являются «танцы». Найдя источник пищи (нектара) и вернувшись в улей, пчела раздает другим пчелам-сборщицам пробы нектара и приступает к «танцу», который состоит из пробежек по сотам. Рисунок танца зависит от местонахождения обнаруженного источника пищи: если он находится рядом с ульем (на расстоянии 2-5 метров от него), то производится «танец-толчок»: пчела беспорядочно бежит посотам, время от времени виляя брюшком; если же корм обнаружен на расстоянии до 100 метров от улья, то выполняется один из описанных К.Фришем танцев — «круговой», который состоит из пробежек по кругу попеременно по часовой стрелке и против нее (рис. 38). Если же нектар обнаружен на большем расстоянии, то выполняется «виляющий» танец, также описанный и изученный Фришем. Это пробежки по прямой, сопровождающиеся виляющими движениями брюшка с возвращением к исходной точке то слева, то справа (рис. 38, а).Интенсивность виляющих движений указывает на расстояние находки: чем ближе находится кормовой объект, тем интенсивнее выполняется танец (рис. 38, б). Кроме того, как тоже показал Фриш, во второй форме танца угол между линией пробежки и вертикалью (на вертикально расположенных сотах) соответствует углу между линией полета пчелы от улья к кормовому объекту и положением солнца. Таким образом, горизонтальное направление истинного полета переносится на вертикальную плоскость, ориентация по положению солнца подменяется ориентацией по земному притяжению. При этом, если следует лететь навстречу солнцу, пчела танцует снизу вверх, если же от солнца, то сверху вниз (рис. 38, в).Таким образом, медоносная пчела способна к транспозиции с фотоменотаксисов на геоменотаксисы: фиксированный угол к видимому положению солнца переносится в темном улье к силе тяжести. Эта способность медоносной пчелы к транспозиции, коренящаяся в широко распространенной среди насекомых способности к переносу менотаксисных движений на вертикальную плоскость (рис. 39), обеспечивает, как мы видим, исключительную полноту и точность передачи информации. Такая транспозиция является частным случаем трансформации, лежащей в основе всякого общения между животными: свойства действительных предметов и ситуации трансформируются на языке животных с помощью видоспецифического «кода» в оптические, акустические и прочие условные сигналы, информирующие других осо-
Рис. 38. «Танцы пчел». Обозначения: а — «круговой танец» (слева) и «виляющий» (справа); б — интенсивность выполнения «виляющих танцев» как показатель расстояния до местонахождения источника пищи (на оси ординат отмечено число «танцев» за 15 секунд, на оси абсцисс — расстояние до пищи в метрах); в — «виляющий танец» как показатель направления полета к источнику пищи: I — навстречу солнцу, II — под углом 60° налево от положения солнца, III — направо от положения солнца, IV — от солнца. Точка — местонахождение улья, крестик — местонахождение пищи. Изображение «танцев» следует представить себе на вертикальной плоскости (по Фришу); г — схема, иллюстрирующая транспозицию угла полета по отношению к положению солнца (горизонтальная плоскость) на вертикальную плоскость сот, на которых пчела выполняет «виляющий танец» (по Кэстнеру) бей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи), — информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступают характер и функциональное значение психики у животных.
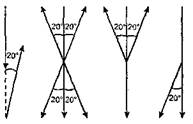
Рис. 39. Транспозиция оптической ориентации в другую сенсорную сферу у разных насекомых. Обозначения: а — менотаксисное движение по горизонтальной плоскости под углом 20° к источнику света. Вертикальная стрелка — направление светового луча; б—г — продолжение движения по вертикальной плоскости у муравья (б), божьей коровки (в), майского жука (г). Вертикальная стрелка — направление силы тяжести Вершиной развития инстинктивных компонентов общения является ритуализация поведения, которая особенно четко выступает в воспроизводительном поведении насекомых, в частности в «ухаживании» самцов за самками. Вот типичный брачный «ритуал» самца бабочки-бархатницы, описанный Тинбергеном: полетев вслед за самкой, самец садится рядом с ней, потом становится перед ней, начинает «дрожать» крыльями (передне-задние движения сомкнутых крыльев), затем повторно раскрывает и закрывает их, что сопровождается постукиванием усиками по крыльям самки. После этого самец обхватывает самку крыльями и зажимает между ними ее усики, которые при этом прикасаются к пахучему участку на крыльях самца. Только после этого, если самка не отвергает «ухаживания» самца, начинается спаривание. В этом примере обращает на себя внимание строгая фиксироваиность последовательности сигнальных поз и телодвижений, что и характерно для ритуализованного поведения. Как мы видим, при ритуализации двигательные акты всегда носят условный характер: выполняемые животным движения несут лишь сигнальную, но не непосредственно «механическую» функцию. Именно на этой основе зародился язык поз, телодвижений и звуков у животных во всем своем удивительном многообразии. При этом в условности сигналов находит непосредственное воплощение психический компонент общения. Наиболее сложные системы общения присущи, конечно, насекомым, живущим большими семьями. Это относится опять-таки в первую очередь к медоносной пчеле и муравьям. Дифференцированные и многообразные взаимоотношения между членами пчелиной семьи, опирающиеся на сложные формы коммуникации, являются уникальными в животном мире. По поводу этого сообщества высказывались самые различные мнения начиная от антропоморфичного толкования ее как копии человеческого общества или государства (сохранилось неудачное, вводящее в заблуждение название «общественные насекомые») до представления о некоей сумме независимо действующих «рефлекторных автоматов». Вместе с тем оказалось (исследования К.П.Трубецкой), что пчелиная семья представляет собой объединение особей, поведение которых носит самостоятельный, но не независимый характер. Самостоятельность поведения отдельных пчел проявляется прежде всего в резко выраженных индивидуальных различиях: предпочтение отдельных видов ульевых работ, различия в сроках и характере перехода от ульевой деятельности к летной и др. Большую сложность в количественном и качественном аспектах достигают взаимоотношения между обитателями улья, в результате чего функциональная настроенность одних пчел может оказать влияние на других. Рабочие особи с их повышенным уровнем нервной деятельности часто сообщают возбуждение другим, менее активным рабочим пчелам. Территориальное поведение Уже у кольчатых червей и низших моллюсков наблюдаются первые проявления территориального поведения, что выражается в приуроченности жизнедеятельности к постоянно занимаемому участку, на котором располагается и убежище («дом») животного. Правда, выполнение жизненных функций (или хотя бы части из них) на определенном участке составляет лишь часть территориального поведения, равно как и наличие «дома» само по себе еще не является критерием последнего. При полноценном территориальном поведении, встречающемся лишь на уровне перцептивной психики, животное-хозяин своим видом, голосом или запахом дает знать сородичам о своем присутствии на занятом участке. Важную роль играет при этом «маркировка местности», т.е. нанесение пахучих или иных меток на разные объекты, особенно на периферии участка. Такими метками могут быть испражнения или выделения кожных желез, а также оптические знаки — содранная со стволов деревьев кора, вытоптанная трава и т.п. Весьма существенным признаком территориального поведения является активная защита участка хозяином. Тинберген придает этому критерию наряду с привязанностью к местности особое значение. Враждебное отношение («агрессивность») проявляется хозяином по отношению к любому представителю того же вида, особенно того же пола. Часто такое поведение приурочено лишь к периоду размножения. Так, например, древесный таракан (Cryptocereus punctulatus), устраивающий гнездовую камеру в гнилой древесине, весьма интенсивно (и обычно успешно) защищает место, где находится эта камера, от вторжения других самцов. В случае поражения гнездовую камеру, где находится и самка, занимает победитель. В высокоразвитых формах территориальное поведение, как показал немецкий этолог А.Хеймер, встречается у стрекоз. Это интересно уже потому, что стрекозы относятся к самым древним насекомым и, появившись в палеозое, дожили до наших дней, не претерпев за эти 50 миллионов лет существенных морфологических изменений. Вместе с тем, как показывает Хеймер, архаичность строения сочетается у них с высокоразвитым поведением, причем не только в сфере территориальности. Этот, казалось бы, парадоксальный факт сохранения древнейших признаков строения при наличии весьма прогрессивных форм поведения вновь подтверждает общее правило несоответствия морфологических и поведенческих признаков, если иметь в виду морфофункциональные отношения на уровне целого организма. Что же касается территориального поведения стрекоз (речь идет конкретно о равнокрылых стрекозах-красотках Zygoptera), то половозрелые самцы отыскивают места, пригодные для постоянного пребывания, которые и становятся их индивидуальными участками. Эти участки маркируются оптически, а именно путем ежедневных многократных облетов, и энергично обороняются от самцов-сородичей. Внутри индивидуального участка имеются основные и дополнительные места отдыха, а также зона для откладки яиц, границы которой маркируются упомянутым образом. В эту зону самец приводит самку посредством совместно выполняемого с ней «танца» над поверхностью воды. Кроме того, самец «ухаживает» за самкой, выполняя на своей территории особый ритуализованный брачный полет, а во время откладки яиц он охраняет и сторожит самку или соответственно несколько самок, ибо один самец нередко спаривается подряд с несколькими самками. Охрана проявляется в том, что самец отгоняет других мужских особей, пытающихся спариваться с «его» самками, что мешает откладыванию яиц, сторожевая же функция состоит в том, что при попытке самок прервать это занятие самец заставляет их вернуться и возобновить его. Добавим еще, что территориальная борьба самцов выполняется в высокоритуализованных формах (подлинная борьба отсутствует! ). В целом все поведение самцов (как и самок) отличается большой пластичностью. Таким образом, у данных видов стрекоз существует вполне развитое территориальное поведение: занимаются индивидуальные участки, в которых выделяются функциональные зоны (отдыха, размножения), на этих участках выполняются все жизненные функции (за исключением сна: вечером самцы собираются в особых местах ночевок), хозяин дает знать о своем присутствии, маркирует и активно защищает участок. Интересные особенности территориального поведения муравьев были выявлены советским энтомологом А.А.Захаровым. Оказалось, что у муравьев существуют два основных типа использования кормовых участков: совместное использование угодий несколькими семьями и использование кормового участка населением лишь одного гнезда. При этом выявилась прямая связь между плотностью муравьев на участке и «агрессивностью»: у видов с низкой плотностью участки не охраняются (кроме пригнездовой зоны), при высокой же плотности на кормовых участках появляются охраняемые территории, а между ними «нейтральные зоны» (рис. 40). Как показал другой советский исследователь, Г.М.Длусский, муравьи того же вида, принадлежащие к другим семьям, равно как и представители других видов, в пределы этих территорий не допускаются.
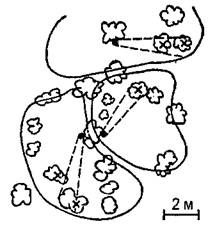
Рис. 40. Территориальное поведение муравьев (Acantholepis melanogaster). Территории трех семей с нейтральными зонами между ними. Точки — гнезда; крестики — колонии тлей, посещаемые муравьями; замкнутые кривые — кустарник; пунктирные линии — основные направления поиска и транспортировки пищи (по Захарову) Наибольшую сложность территориальное поведение достигло у рыжих лесных муравьев, у которых наблюдается и наибольшая плотность особей на кормовых участках. Охраняемые территории этих муравьев достигают значительно большей величины, чем у других видов. Центральными элементами такой территории являются муравьиные тропы (постоянные кормовые дороги), длина которых нередко превышает 100 метров, а протяженность всей сети троп — 1 километр. Каждой дорогой пользуется лишь определенная группа муравьев («колонна»), занимающая в муравейнике определенный сектор, примыкающий к «своей» дороге. Территория всей семьи подразделяется дорогами на отдельные части, составляющие территории отдельных колонн. Между ними имеются такие же нейтральные пространства, как и между территориями семей, хотя и не столь выраженные, как в последнем случае. Границы территории маркированы и препятствуют «рассеиванию» муравьев. Муравьи двигаются по территории не хаотично, а в соответствии со структурой территории: в центральной и средней ее частях — по кратчайшему пути от дорог или муравейника, на периферии же — параллельно границам территории. Таким образом, траектория движения каждого муравья детерминируется местом нахождения его на территории (исследования И.В.Стебаева). Наибольшую охотничью активность муравьи развивают на периферии территории (далее 10 метров от гнезда). Сбор строительного материала производится в пределах двухметровой полосы вдоль дорог. Захаров справедливо отмечает сходство территориального поведения муравьев с таковым у птиц и хищных млекопитающих. Популярное:
|
Последнее изменение этой страницы: 2017-03-03; Просмотров: 481; Нарушение авторского права страницы