|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Химический состав костной тканиСтр 1 из 31Следующая ⇒
Межклеточный органический матрикс компактной кости составляет около 20%, неорганические вещества – 70% и вода – 10%. В губчатой кости преобладают органические компоненты, которые составляют более 50%, на долю неорганических соединений приходится 33-40%. Количество воды приблизительно то же, что и в компактной кости. Органический матрикс костной ткани. Приблизительно 95% органического матрикса приходится на коллаген типа I. Данный тип коллагена входит также в состав сухожилий и кожи, однако коллаген костной ткани обладает некоторыми особенностями. В нем несколько больше оксипролина, а также свободных аминогрупп лизиновых и оксилизиновых остатков. Это обусловливает наличие большего количества поперечных связей в коллагеновых волокнах и их большую прочность. По сравнению с коллагеном других тканей костный коллаген характеризуется повышенным содержанием фосфата, который в основном связан с остатками серина. Белки неколлагеновой природы представлены гликопротеинами, белковыми компонентами протеогликанов. Принимают участие в росте и развитии кости, процессе минерализации, водно-солевом обмене. Альбумины участвуют в транспорте гормонов и других веществ из крови. Преобладающим белком неколлагеновой природы является остеокальцин. Он присутствует только в костях и зубах. Это небольшой (49 аминокислотных остатков) белок, называемаый также костным глутаминовым белком или gla-белком. В молекуле остеокальцина обнаружены три остатка
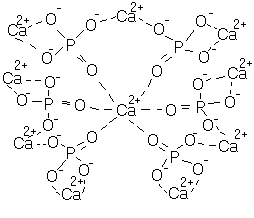
Рис. 34. Посттрансляционная модификация остеокальцина В состав органического матрикса костной ткани входят гликозаминогликаны, основным представителем которых является хондроитин-4-сульфат. Хондроитин-6-сульфат, кератансульфат и гиалуроновая кислота содержатся в небольших количествах. Окостенение сопровождается изменением гликозаминогликанов: сульфатированные соединения уступают место несульфатированным. Гликозаминогликаны участвуют в связывании коллагена с кальцием, регуляции водного и солевого обмена. Цитрат необходим для минерализации костной ткани. Он образует комплексные соединения с солями кальция и фосфора, обеспечивая возможность повышения концентрации их в ткани до такого уровня, при котором могут начаться кристаллизация и минерализация. Также принимет участие в регуляции уровня кальция в крови. Кроме цитрата, в костной ткани обнаружены сукцинат, фумарат, малат, лактат и другие органические кислоты. Костный матрикс содержит небольшое количество липидов. Липиды играют существенную роль в образовании ядер кристаллизации при минерализации кости. Остеобласты богаты РНК. Высокое содержание РНК в костных клетках отражает их активность и постоянную биосинтетическую функцию. Неорганический состав костной ткани. В раннем возрасте в костной ткани преобладает аморфныйм фосфат кальция Са3(РО4)2. В зрелой кости преобладающим становится кристаллический гидроксиапатит Са10(РО4)6(ОН)2 (рис. 35). Его кристаллы имеют форму пластин или палочек. Обычно аморфный фосфат кальция рассматривают как лабильный резерв ионов Са2+ и фосфата. В состав минеральной фазы кости входят ионы натрия, магния, калия, хлора и др. В кристаллической решетке гидроксиапатита ионы Са2+ могут замещаться другими двухвалентными катионами, тогда как анионы, отличные от фосфата и гидроксила, либо адсорбируются на поверхности кристаллов, либо растворяются в гидратной оболочке кристаллической решетки.
Рис. 35. Строение кристалла гидроксиапатита
Метаболизм костной ткани характеризуется двумя противоположными процессами: образованием новой костной ткани остеобластами и резорбцией (деградацией) старой остеокластами. В норме количество новообразованной ткани эквивалентно разрушенной. Костная ткань скелета человека практически полностью перестраивается в течение 10 лет. Образование костной ткани На 1 этапе остеобласты синтезируют сначала протеогликаны и гликозаминогликаны, образующие матрикс, а затем продуцируют фибриллы костного коллагена, которые распределяются в матриксе. Костный коллаген является матрицей для процесса минерализации. Необходимым условием процесса минерализации является пересыщение среды ионами кальция и фосфора. Образование кристаллов минерального остова кости запускают На 2 этапе в зоне минерализации при участии лизосомных протеиназ происходит деградация протеогликанов; усиливаются окислительные процессы, распадается гликоген, синтезируется необходимое количество АТФ. Кроме того, в остеобластах увеличивается количество цитрата, необходимого для синтеза аморфного фосфата кальция. По мере минерализации костной ткани кристаллы гидроксиапатита вытесняют не только протеогликаны, но и воду. Плотная, полностью минерализованная кость практически обезвожена. Фермент щелочная фосфатаза принимает участие в минерализации. Одним из механизмов ее действия является локальное увеличение концентрации ионов фосфора до точки насыщения, за которым следуют процессы фиксации кальций-фосфорных солей на органической матрице кости. При восстановлении костной ткани после переломов содержание щелочной фосфатазы в костной мозоли резко увеличивается. При нарушении костеобразования наблюдается уменьшение содержания и активности щелочной фосфатазы в костях, плазме и в других тканях. Ингибитором кальцификации является неорганический пирофосфат. Ряд исследователей считают, что процессу минерализации коллагена в коже, сухожилиях, сосудистых стенках препятствует постоянное наличие в этих тканях протеогликанов. Процессы моделирования и ремоделирования обеспечивают постоянное обновление костей, а также модификацию их формы и структуры. Моделирование (образование новой кости) имеет место в основном в детском возрасте. Ремоделирование является доминирующим процессом в скелете взрослых; в этом случае происходит лишь замена отдельного участка старой кости. Таким образом, в физиологических и патологическтх условиях происходит не только образование, но и резорбция костной ткани. Катаболизм костной ткани Практически одновременно имеет место «рассасывание» как минеральных, так и органических структур костной ткани. При остеолизе усиливается продукция органических кислот, что приводит к сдвигу рН в кислую сторону. Это способствует растворению минеральных солей и их удалению. Резорбция органического матрикса происходит под действием лизосомных кислых гидролаз, спектр которых в костной ткани довольно широк. Они участвуют во внутриклеточном переваривании фрагментов резорбируемых структур. При всех заболеваниях скелета происходят нарушения процессов ремоделирования кости, что сопровождается возникновением отклонений в уровне биохимических маркеров. Имеются общие маркеры формирования новой костной ткани, такие как костно-специфическая щелочная фосфатаза, остеокальцин плазмы, проколлаген I, пептиды плазмы. К биохимическим маркерам резорбции кости относятся кальций в моче и гидроксипролин, пиридинолин мочи и дезоксипиридинолин, являющиеся производными поперечных волокон коллагена, специфичных для хрящей и костей. Факторами, влияющими на метаболизм костной ткани, являются гормоны, ферменты и витамины. Минеральные компоненты костной ткани находятся практически в состоянии химического равновесия с ионами кальция и фосфата сыворотки крови. В регуляции поступления, депонирования и выделения кальция и фосфата важную роль играют паратгормон и кальцитонин. Действие паратгормона приводит к увеличению числа остеокластов и их метаболической активности. Остеокласты способствуют ускоренному растворению содержащихся в костях минеральных соединений. Таким образом, происходит активация клеточных систем, участвующие в резорбции кости. Паратгормон увеличивает также реабсорбцию ионов Са2+ в почечных канальцах. Суммарный эффект проявляется в повышении уровня кальция в сыворотке крови. Действие кальцитонина состоит в снижении концентрации ионов Са2+ за счет отложения его в костной ткани. Он активирует ферментную систему остеобластов, повышает минерализацию кости и уменьшает число остеокластов в зоне действия, т. е. угнетает процесс костной резорбции. Все это увеличивает скорость формирования кости. Витамин D участвует в биосинтезе Са2+-связывающих белков, стимулирует всасывание калиция в кишечнике, повышает реабсорбцию кальция, фосфора, натрия, цитрата, аминокислот в почках. При недостатке витамина D эти процессы нарушаются. Прием в течение длительного времени избыточных количеств витамина D приводит к деминерализации костей и увеличению концентрации кальция в крови. Кортикостероиды увеличивают синтез и секрецию паратгормона, усиливают деминерализацию кости; половые гормоны ускоряют созревание и сокращают период роста кости; тироксин усиливает рост и дифференцировку ткани. Действие витамина С на метаболизм костной ткани обусловлено, прежде всего, влиянием на процессе биосинтеза коллагена. Аскорбиновая кислота является кофактором пролил- и лизилгидроксилаз и необходима для осуществления реакции гидроксилирования пролина и лизина. Недостаток витамина С приводит также к изменениям в синтезе гликозаминогликанов: содержание гиалуроновой кислоты в костной ткани увеличивается в несколько раз, тогда как биосинтез хондроитинсульфатов замедляется. При недостатке витамина А происходит изменение формы костей, нарушение минерализации, задержка роста. Считают, что данный факт обусловлен нарушением синтеза хондроитинсульфата. Высокие дозы витамина А приводят к избыточной резорбции кости. При недостатке витаминов группы В рост кости замедляется, что связано с нарушением белкового и энергетического обмена. Особенности зубной ткани Основную часть зуба составляет дентин. Выступающая из десны часть зуба, коронка, покрыта эмалью, а корень зуба покрыт зубным цементом. Цемент, дентин и эмаль построены подобно костной ткани. Белковый матрикс этих тканей состоит главным образом из коллагенов и протеогликанов. Содержание органических компонентов в цементе – около 13%, в дентине – 20%, в эмали – всего 1-2%. Высокое содержание минеральных веществ (эмаль – 95%, дентин – 70%, цемент – 50%) определяет высокую твердость зубной ткани. Наиболее важным минеральным компонентом является гидроксиапатит [Са3РО4)2]3 • Са(ОН)2. Содержатся также карбонатный апатит, хлорапатит и стронцевый апатит. Эмаль, покрывающая зуб, полупроницаема. Она участвует в обмене ионами и молекулами со слюной. На проницаемость эмали влияют рН слюны, а также ряд химических факторов. В кислой среде ткань зуба подвергается атаке и утрачивает твердость. Такое распространенное заболевание, как кариес, вызывается микроорганизмами, живущими на поверхности зубов и выделяющими в качестве продукта анаэробного гликолиза органические кислоты, вымывающие из эмали ионы Са2+. Контрольные вопросы 1. Назовите основные органические компоненты костной ткани. 2. Какие неорганические соединения входят в состав костной ткани? 3. В чем различие биохимических процессов, протекающих в остеокластах и остеобластах? 4. Опишите процесс формирования кости. 5. Какие факторы влияют на формирование костной ткани и ее метаболизм? 6. Какие вещества могут быть биохимическими маркерами процессов, протекающих в костной ткани? 7. Каковы особенности биохимического состава зубной ткани? Литература 1. Березов, Т.Т. Биологическая химия. / Т.Т. Березов, Б.Ф. Коровкин. - М.: ОАО «Издательство «Медицина»», 2007. - 704 с. 2. Биохимия. / Под ред. Е.С. Северина. - М.: ГЭОТАР-Медиа, 2014. - 3. Биологическая химия с упражнениями и задачами. / Под ред. Е.С. Северина. - М.: ГЭОТАР-Медиа, 2013. - 624 с. 4. Зубаиров, Д.М. Руководство к лабораторным занятиям по биологической химии. / Д.М. Зубаиров, В.Н. Тимербаев, В.С. Давыдов. - М.: ГЭОТАР-Медиа, 2005. - 392 с. 5. Шведова, В.Н. Биохимия. /В.Н. Шведова. – М.: Юрайт, 2014. – 640 с. 6. Николаев, А.Я. Биологическая химия. / А.Я. Николаев. - М.: Медицинское информационное агентство, 2004. - 566 с. 7. Кушманова, О.Б. Руководство к лабораторным занятиям по биологической химии. / О.Б. Кушманова, Г.И. Ивченко. - М. - 1983. 8. Ленинджер, А. Основы биохимии / А. Ленинджер. - М., «Мир». - 1985. 9. Марри, Р. Биохимия человека. / Р. Марри, Д. Греннер, П. Мейес, В. Родуэлл. - Т. 1. - М.: Мир, 1993. - 384 с. 10. Марри, Р. Биохимия человека. / Р. Марри, Д. Греннер, П. Мейес, В. Родуэлл. - Т. 2. - М.: Мир, 1993. - 415 с. Популярное:
|
Последнее изменение этой страницы: 2017-03-11; Просмотров: 809; Нарушение авторского права страницы