
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Ферментативная активность лизоцима
Лизоцим — это фермент, разрушающий определённые бактериальные клетки (расщепляет полисахаридные цепи клеточной стенки, после чего клетка разрывается под действием осмотического шока — входа воды внутрь клетки). Полисахарид клеточной стенки — полимер, в котором чередуются остатки сахаров двух типов — N-ацетилглюкозамин (NAG) и N-ацетилмурамовая кислота (NAM). Сахара, имеющие β -конфигурацию относительно С1-атома, образуют полимерную цепь с помощью гликозидных связей между С1- одного сахарного кольца и С4-атомом следующего. Структура лизоцима. Лизоцим (из белка куриного яйца) состоит из одной полипептидной цепи, насчитывающей 129 остатков и имеющей 4 дисульфидных мостика. Фермент может связываться с ингибиторами, химически сходными с полисахаридом клеточной стенки. С помощью рентгеноструктурного анализа Филипсом и др. была определена пространственная структура белка как такового и белка, закристаллизованного вместе с ингибитором. Оказалось, что лизоцим состоит из двух доменов (α и β -типов), образующих щель, в которой находится активный центр, способный связать гексосахарид, причём, для связывания каждого из шести сахарных колец на ферменте имеется свой участок (это участки A, B, C, D, E, F). Расщепление полисахарида осуществляется путём гидролиза гликозидной связи между сахарными кольцами, располагающимися в участках D и E. Анализ кристаллографических данных позволил предположить, что непосредственно перед и после гидролиза сахарное кольцо в участке D имеет не обычную конформацию кресла, а конформацию полукресла, характеризующуюся тем, что 5 из 6 атомов, образующих сахарное кольцо, лежат практически в одной плоскости. Центральную роль в функционировании лизоцима играют остатки глутаминовой кислоты Glu35 и аспарагиновой кислоты Asp 52. Боковые цепи этих остатков располагаются близко к гликозидной связи (~ 0.3 нм) между сахарными кольцами, локализованными в участках D и E. Glu 35 находится в неполярном окружении, и поэтому его карбоксильная группа остаётся протонированной (т.е. в форме СООН). Окружение Asp 52, наоборот, полярно и эта группа депротонирована (–СОО- ). Реакцию гидролиза можно подразделить на несколько этапов: 1. –СООН-группа Glu 35 предоставляет свой протон гликозидному кислороду, что приводит к разрыву связи между этим атомом кислорода и С1-атомом сахарного кольца, располагающегося в участке D. Получившийся в результате фрагмент исходного полисахарида, включающий в себя сахарные кольца, которые находятся в участках E и F, является продуктом и может освободиться из комплекса с ферментом. 2. Сахарное кольцо, располагающееся в D–участке, имеет искажённую конформацию, соответствующую конформации переходного состояния. При этом С1-атом оказывается положительно заряженным. Такое состояние углеродного атома называется карбоний-ионом. Оно стабилизируется с помощью отрицательного заряда близко расположенного остатка Asp 52.
Рис.15.Связывание сахаров с лизоцимом стабилизируется при помощи гидрофобных взаимодействий и водородных связей. 3. Гидроксильный ион ОН-, донором которого является Н2О из окружающей среды, присоединяется к карбоний-иону, после чего второй фрагмент расщепленного полисахарида становится продуктом реакции. Одновременно из-за связывания иона водорода (Н+) протонируется карбоксильная группа Glu 35, переходя в форму –СООН. 4. Теперь фермент находится в первоначальном состоянии и готов осуществить следующую реакцию расщепления полисахарида. Этот пример позволяет проследить некоторые общие принципы ферментативного катализа. 1. Увеличение энергии субстрата за счёт искажения структуры сахарного кольца NAM, находящегося в D-участке. 2. Наличие необычного окружения Glu 35, обусловливающее появление реакционноспособного протона. 3. Правильная ориентация протона в Glu 35, необходимая для атаки гликозидной связи. 4. Уменьшение свободной энергии переходного состояния за счёт стабилизации карбоний-иона карбоксильной группой остатка Asp52. Глава 5. Физика нуклеиновых кислот.
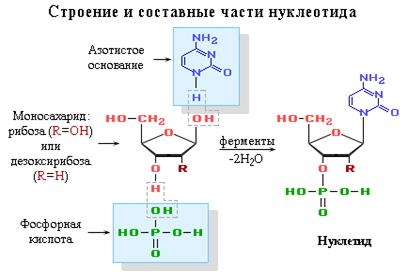
Основная характеристика Нуклеиновые кислоты - это биополимеры, макромолекулы которых состоят из многократно повторяющихся звеньев - нуклеотидов. Поэтому их называют также полинуклеотидами. Важнейшей характеристикой нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида - структурного звена нуклеиновых кислот - входят три составные части:
Строение пуриновых оснований:
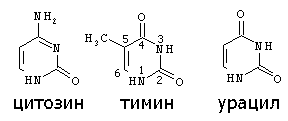
Строение пиримидиновых оснований:
Строение моносахаридов:
В конце 40-х — начале 50-х годов, когда появились такие методы исследования, как хроматография на бумаге и УФ-спектроскопия, были проведены многочисленные исследования нуклеотидного состава НК (Чаргафф, А. Н. Белозерский). Полученные данные позволили решительно отбросить старые представления о нуклеиновых кислотах, как о полимерах, содержащих повторяющиеся тетрануклеотидные последовательности (так называемая тетрануклеотидная теория строения ПК, господствовавшая в 30—40-е годы), и подготовили почву для создания современных представлений не только о первичной структуре ДНК и РНК, но и об их макромолекулярной структуре и функциях. Метод определения состава НК основан на анализе гидролизатов, образующихся при их ферментативном или химическом расщеплении. Обычно используются три способа химического расщепления НК. Кислотный гидролиз в жестких условиях (70%-ная хлорная кислота, 100°С, 1 ч или 100%-ная муравьиная кислота, 175°C, 2 ч), применяемый для анализа как ДНК, так и РНК, приводит к разрыву всех N-гликозидных связей и образованию смеси пуриновых и пиримидиновых оснований. При исследовании РНК могут использоваться как мягкий кислотный гидролиз (1 н соляная кислота, 100°C, 1 ч), в результате которого образуются пуриновые основания и пиримидиновые нуклеозид-2'(3')-фосфаты, так и щелочной гидролиз (0, 3 н. едкое кали, 37°С, 20 ч), дающий смесь нуклеозид -2' (3') -фосфатов. Поскольку в НК число нуклеотидов каждого вида равно числу соответствующих оснований, для установления нуклеотидного состава данной НК достаточно определить количественное соотношение оснований. Для этой цели из гидролизатов с помощью хроматографии на бумаге или электрофореза (когда в результате гидролиза получают нуклеотиды) выделяют индивидуальные соединения. Каждое основание независимо от того, связано оно с углеводным фрагментом или нет, обладает характерным максимумом поглощения в УФ, интенсивность которого зависит от концентрации. По этой причине, исходя из УФ-спектров выделенных соединений, можно определить количественное соотношение оснований, а следовательно, и нуклеотидный состав исходной НК. При количественном определении минорных нуклеотидов, особенно таких неустойчивых, как дигидроуридиловая кислота, пользуются ферментативными методами гидролиза (ФДЭ змеиного яда и селезенки). Использование описанных выше аналитических приемов показало, что НК различного происхождения состоят за редким исключением из четырех основных нуклеотидов и что содержание минорных нуклеотидов может меняться в значительных пределах. Как будет показано далее, при изучении нуклеотидного состава ДНК были получены данные, которые помогли установить ее пространственную структуру. Нуклеиновые кислоты, полинуклеотиды, важнейшие биологически активные биополимеры, имеющие универсальное распространение в живой природе. Содержатся в каждой клетке всех организмов. НК были открыты в 1868 швейцарским учёным Ф. Мишером в клеточных ядрах (отсюда название: лат. nucleus - ядро), выделенных из гноя, а также из спермы лосося. Позднее нуклеиновые кислоты были обнаружены не только в ядре, но и в цитоплазме. Различают два главных типа нуклеиновых кислот - дезоксирибонуклеиновые кислоты, или ДНК, содержащиеся преимущественно в ядрах клеток, и рибонуклеиновые кислоты, или РНК, находящиеся главным образом в цитоплазме. Молекулы нуклеиновых кислот - длинные полимерные цепочки с молекулярной массой 2, 5 104 – 4 109, построенные из мономерных молекул - нуклеотидов так, что гидроксильные группы у 3' и 5' углеродных атомов углевода соседних нуклеотидов связаны остатком фосфорной кислоты. В состав РНК в качестве углевода входит рибоза, а азотистые компоненты представлены аденином, гуанином (пуриновые основания), урацилом и цитозином (пиримидиновые основания). В ДНК углеводным компонентом является дезоксирибоза, а урацил заменен тимином (5-метилурацилом). Фосфат и сахар составляют неспецифическую часть в молекуле нуклеотида, а пуриновое или пиримидиновое основание - специфическую. В составе большинства нуклеиновых кислот обнаружены в небольших количествах также некоторые другие (главным образом метилированные) производные пуринов и пиримидинов - т. н. минорные основания. Нуклеиновые кислоты имеют различающийся состав. В частности, дезоксирибонуклеиновые кислоты (ДНК) содержат дезоксирибозу, а рибонуклеиновые кислоты (РНК) - рибозу. Эти и другие отличия в составе нуклеиновых кислот приведены в таблице:
Цепи нуклеиновых кислот содержат от нескольких десятков до многих тысяч нуклеотидных остатков, расположенных линейно в определённой последовательности, уникальной для данной нуклеиновой кислоты. Т.е., как РНК, так и ДНК представлены огромным множеством индивидуальных соединений. Линейная последовательность нуклеотидов определяет первичную структуру нуклеиновых кислот. Вторичная структура нуклеиновых кислот возникает в результате сближения определённых пар оснований, а именно: гуанина с цитозином и аденина с урацилом (или тимином) по принципу комплементарности за счёт водородных связей, а также гидрофобных взаимодействий между ними. Биологическая роль нуклеиновых кислот заключается в хранении, реализации и передаче наследственной информации, " записанной" в молекулах нуклеиновых кислот в виде последовательности нуклеотидов - т. н. генетического кода. При делении клеток - митозе - происходит самокопирование ДНК - её репликация, в результате чего каждая дочерняя клетка получает равное количество ДНК, заключающей программу развития всех признаков материнской клетки. Реализация этой генетической информации в определённые признаки осуществляется путём биосинтеза молекул РНК на молекуле ДНК (транскрипция) и последующего биосинтеза белков с участием разных типов РНК (трансляция). Исследование строения и функций нуклеиновых кислот в 50-70-х гг. 20 в. обусловило огромные успехи молекулярной генетики и молекулярной биологии. Важнейшим этапом в изучении химии и биологии НК было создание в 1953 Дж. Уотсоном и Ф. Криком модели ДНК (двойная спираль), что позволило объяснить многие её свойства и биологические функции. Нуклеиновые кислоты обнаружены также в клеточных органеллах (хлоропластах, митохондриях и др.), где функции их изучаются. Сравнительный анализ нуклеиновых кислот в разных группах организмов играет важную роль при решении вопросов систематики и эволюции. Каждый вид организмов содержит специфичные нуклеиновые кислоты (как РНК, так и ДНК). Степень сходства в строении нуклеиновых кислот указывает на уровень филогенетической близости организмов. Нуклеозиды - соединения азотистого основания и углеводов (рибозы и дезоксирибозы). Нуклеозиды образуются за счет N-гликозидной связи между девятым атомом азота у пуриновых (первым атомом азота - у пиримидиновых) оснований и гидроксилом первого атома углерода рибозы или дезоксирибозы. Во избежание путаницы, нумерация атомов азотистых оснований осуществляется арабскими цифрами, а у атомов углерода рибоз - арабскими цифрами со “штрихом”.
Нуклеотиды отличаются от нуклеозидов наличием остатков фосфорной кислоты (от одного до трех), связанных простой эфирной связью с гидроксилом 5' атома углерода рибоз. Остатки фосфорных кислот между собой также связаны простой эфирной связью. В зависимости от числа остатков фосфорной кислоты в нуклеотидах различают моно-, ди- и трифосфонуклеотиды. Их номенклатура приведена в таблице:
5'-НО-G-A-A - T-C-T-A-C-A-…3' Вследствие наличия сильно диссоциирующих фосфатных групп, нуклеиновые кислоты легко образуют связи с основными белками с образованием нуклеопротеинов. Протеины отделяются от НК детергентами или после расщепления белков протеиназами НК осаждаются спиртом. Подобно белкам, ДНК имеют первичную, вторичную и третичную структуру.
Первичная структура Первичная структура есть последовательность чередования нуклеотидов в цепи ДНК и РНК. Сложность расшифровки структуры связана с наличием всего 4-х видов нуклеотидов при их огромном числе в молекуле.
Состав ДНК Исследуя нуклеотидный состав нативных ДНК различного происхождения, Чаргафф обнаружил следующие закономерности. 1. Все ДНК независимо от их происхождения содержат одинаковое число пуриновых и пиримидиновых оснований. Следовательно, в любой ДНК на каждый пуриновый нуклеотид приходится один пиримидиновый. 2. Любая ДНК всегда содержит в равных количествах попарно аденин и тимин, гуанин и цитозин, что обычно обозначают как А=Т и G=C. Из этих закономерностей вытекает третья. 3. Количество оснований, содержащих аминогруппы в положении 4 пиримидинового ядра и 6 пуринового (цитозин и аденин), равно количеству оснований, содержащих оксо-группу в тех же положениях (гуанин и тимин), т. е. A+C=G+T. Эти закономерности получили название правил Чаргаффа. Наряду с этим было установлено, что для каждого типа ДНК суммарное содержание гуанина и цитозина не равно суммарному содержанию аденина и тимина, т. е. что (G+C)/(A+T), как правило, отличается от единицы (может быть как больше, так и меньше ее). По этому признаку различают два основных типа ДНК: А-Т-тип с преимущественным содержанием аденина и тимина и G-C-тип с преимущественным содержанием гуанина и цитозина. Величину отношения содержания суммы гуанина и цитозина к сумме содержания аденина и тимина, характеризующую нуклеотидный состав данного вида ДНК, принято называть коэффициентом специфичности. Каждая ДНК имеет характерный коэффициент специфичности, который может изменяться в пределах от 0, 3 до 2, 8. При подсчете коэффициента специфичности учитывается содержание минорных оснований, а также замены основных оснований их производными, например, при подсчете коэффициента специфичности для ДНК зародышей пшеницы, в которой содержится 6% 5-метилцитозина. Последний входит в сумму содержания гуанина (22, 7%) и цитозина (16, 8%). Смысл правил Чаргаффа для ДНК стал понятным после установления ее пространственной структуры.
Состав РНК Первые сведения о нуклеотидном составе РНК относились к препаратам, представляющим собой смеси клеточных РНК (рибосомных, информационных и транспортных) и называемым обычно суммарной фракцией РНК. Правила Чаргаффа в этом случае не соблюдаются, хотя определенное соответствие между содержанием гуанина и цитозина, а также аденина и урацила все же имеет, место. Данные, полученные в последние годы при анализе индивидуальных РНК, показывают, что и на них правила Чаргаффа не распространяются. Однако различия в содержании аденина и урацила, а также гуанина и цитозина для большинства РНК невелики и что, следовательно, тенденция к выполнению указанных правил все же наблюдается. Этот факт объясняется особенностями макроструктуры РНК. Характерными структурными элементами некоторых РНК являются минорные основания. Соответствующие им нуклеотидные остатки обычно входят в состав транспортных и некоторых других РНК в очень небольших количествах, поэтому определение полного нуклеотидного состава таких РНК представляет собой иногда весьма сложную задачу.
ФРАГМЕНТ МОЛЕКУЛЫ РНК. Основное отличие от ДНК – наличие группировок ОН в рибозе (красный цвет) и фрагмента урацила (синий цвет). |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 524; Нарушение авторского права страницы

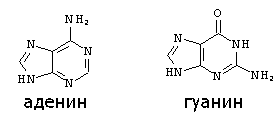




 Собственно нуклеиновые кислоты представляют собой полинуклеотидмоно-фосфаты. Полимерная цепь образуется за счет фосфодиэфирной связи между 3'- гидроксилом одного нуклеотида и 5'- гидроксилом другого. Таким образом, первичная структура нуклеиновых кислот представляет собой порядок чередования нуклеотидов в полинуклеотидной цепи. Один из концов этой цепи (изображаемый слева) имеет свободный гидроксил при 5' - атоме С, а другой (изображаемый справа) - свободный гидроксил при 3'- атоме углерода рибоз. Поскольку основой нуклеиновых кислот является сахарофосфатный остов, в сокращенных написаниях участков цепи используют лишь однобуквенные символы, соответствующего азотистого основания. Полное и схематичное обозначения участка полинуклеотидной цепи приведены ниже:
Собственно нуклеиновые кислоты представляют собой полинуклеотидмоно-фосфаты. Полимерная цепь образуется за счет фосфодиэфирной связи между 3'- гидроксилом одного нуклеотида и 5'- гидроксилом другого. Таким образом, первичная структура нуклеиновых кислот представляет собой порядок чередования нуклеотидов в полинуклеотидной цепи. Один из концов этой цепи (изображаемый слева) имеет свободный гидроксил при 5' - атоме С, а другой (изображаемый справа) - свободный гидроксил при 3'- атоме углерода рибоз. Поскольку основой нуклеиновых кислот является сахарофосфатный остов, в сокращенных написаниях участков цепи используют лишь однобуквенные символы, соответствующего азотистого основания. Полное и схематичное обозначения участка полинуклеотидной цепи приведены ниже: 
