|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
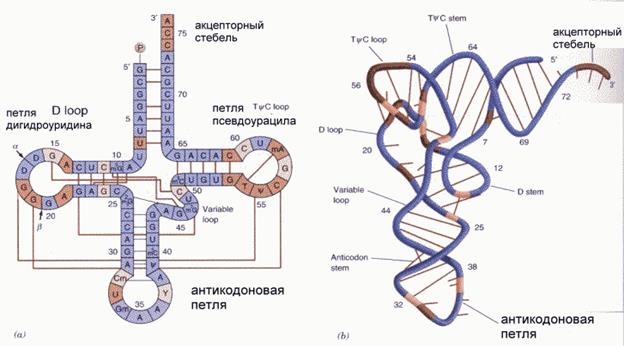
Макромолекулярная структура тРНК
РНК состоит из одной полинуклеотидной цепи, закрученной на себя, образует короткие двуспиральные шпильки в палиндромных участках (Г с Ц, А с У). Различают три вида РНК: рибосомная (рРНК) – 80%, транспортная (тРНК) -15%, и информационная или матричная (мРНК) – 5%. тРНК - самые мелкие молекулы (ММ=23-30 тыс.). Они являются переносчиками аминокислот. Каждая тРНК переносит только одну аминокислоту, но на одну аминокислоту имеется более одной тРНК. Всего известно 61 тРНК.
Строение тРНК. Имеет акцептирующий участок (присоединяет АК, при участии АТФ), общий участок (петля дигидроуридина) обеспечивает связь со специфическим ферментом, характерный участок (петля псевдо урацила) всегда содержит последовательность 5'-TЦГ-3', этой петлей взаимодействует с рибосомой. Антикодонавая петля – содержит 3 нуклеотида, комплементарных кодону данной аминокислоты в мРНК, например, кодону 5'-ГЦЦ-3' в мРНК соответствует антикодон 3'-ЦГГ-5' в тРНК. Этим обеспечивается специфичность взаимодействия с матричной РНК.
Физико-химические свойства ДНК Молекулярная масса ДНК определяется рядом методов, в том числе: а) ультрацентрифугированием в градиенте CsCl (ММ от 200000 до 109); б) по вязкости растворов (М > 109, поскольку при центрифугировании такие длинные молекулы разрываются под действием собственного веса). Вязкость Растворы нативной ДНК очень вязкие. Однако это не означает, что вся макромолекула ДНК является жестким стержнем. Данные, полученные методами рассеяния света, седиментации, вискозиметрии показывают, что двойная спираль нативной ДНК свернута в рыхлый клубок [3].
Уравнение Флори - Манделькерна:
где Мω -средневесовая молекулярная масса полимера, S – константа седиментации, [η ] - характеристическая вязкость, η 0- вязкость растворителя, NA - число Авогадро, ρ - плотность растворителя, β - эмпирический коэффициент. Для ДНК β =2, 5· 106 и β =2, 2· 106 для денатурированной [3]. Формулы Зимма и Крозера:
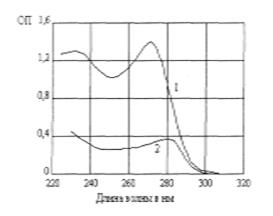
Оптические свойства Поглощение света хромофорными группами зависит от дипольных моментов перехода, в частности от их взаимодействия, если они находятся близко друг от друга, как это имеет место в двойной спирали ДНК. В случае, когда дипольные моменты перехода расположены хаотически, взаимодействия их взаимно нейтрализуются. При переходе спираль—клубок будет наблюдаться увеличение оптической плотности - гиперхромный эффект, а при переходе клубок-спираль - уменьшение оптической плотности, т.е. гипохромный эффект. Если нанести тонкую пленку полинуклеотида из раствора на кварц, подсушить и осторожно ориентировать растяжением, то макромолекулы ЦНК оказываются ориентированными преимущественно вдоль оси растяжения. Такой образец обладает большим дихроизмом, т.е. по-разному поглощает свет, поляризованный вдоль направления ориентации пленки и перпендикулярно ей. На рис. представлены кривые дихроизма в ориентированных пленках полицитидиловой кислоты.
Максимум поглощения приходится на 270 нм. Поглощается главным образом свет, поляризованный перпендикулярно оси макромолекулы. Это означает, что дипольные моменты перехода ориентированы перпендикулярно оси спирали, т.е. лежат в плоскости гетероциклических оснований. В случае полинуклеотидов свет поглощается электронами гетероциклических ядер. Эти электроны образуют делокализованные π -орбиты, и поглощение означает переход электрона с π -орбиты на возбужденный уровень π → π *[3]. Более слабый максимум поглощения наблюдается при 280 нм. К гетероциклическому кольцу присоединены такие группы (например, С=О), в которых электроны локализованы и не входят в сопряженную систему π -связей. Электрон, локализованный в связи С=О - n-состояние, может быть возбужден на π *-уровень, т. е. переходит в кольцо. Дипольные моменты перехода расположены перпендикулярно плоскости кольца. Сильнее поглощается свет, поляризованный параллельно оси макромолекулы [3].
КД спектры очень чувствительны к изменению конформации биополимеров. На рис.2 приведены КД-спектры ДНК в А-, В- и С-форме [46].
Оптическая активность [α ] сильно зависит от длины волны света (уменьшается с ростом длины волны). В полосе поглощения света вращение становится особенно большим аномальная дисперсия оптической активности [3].
Денатурация и ренатурация Денатурация ДНК сводится к разрушению двойной спирали. Нагревание раствора нативной ДНК вызывает разделение двойной спирали на две цепи, сворачивающиеся в статистические клубки. При этом значительно уменьшаются вязкость и оптическая активность. При денатурации гипохромизм исчезает, т.е. на 30-40% возрастает интенсивность поглощения в области 260 нм [13]. Для определения средней степени перехода Θ т.е. доли звеньев, находящихся в клубкообразном состоянии, можно использовать соотношение:
где через
Денатурация происходит также при увеличении рН раствора до уровня, при котором разрушаются водородные связи между основаниями. Поскольку для разрушения двух водородных связей АТ-пар требуется меньше энергии, чем для разрыва трех водородных связей GC-nap, значения температуры и рН, при которых происходит денатурация, зависят от нуклеотидного состава ДНК.
В качестве простейшей характеристики плавления данной ДНК обычно используют температуру плавления Tm, которая определяется как температура, при которой половина звеньев молекулы находится в клубкообразном состоянии. Линейная зависимость температуры плавления ДНК от содержания GC- пар дает при экстраполяции предельные значения Tm=69 С для поли-AT и 110°С для поли-GC, хорошо согласующиеся с экспериментальными значениями для соответствующих синтетических полинуклеотидов (65 и 104°С) [4]. При заданном составе растворителя температура плавления:
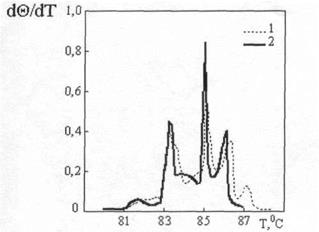
где через ТAT и TGC обозначены температуры плавления молекул ДНК, состоящих только из AT- и только из GC-nap, соответственно; xGC— доля GC-пар. Температура плавления ДНК растет с увеличением ионной силы раствора приблизительно пропорционально логарифму концентрации катионов. При физиологических условиях в растворе Tm=85-95°С. Tm возрастает на 16, 6°C при каждом десятикратном увеличении концентрации моновалентных ионов [13]. Кривая плавления ДНК, т.е. зависимость Θ от T, обладает тонкой структурой, если длина ДНК не превышает несколько десятков тысяч пар оснований. Особенно ярко эта тонкая структура проявляется на дифференциальной кривой плавления, т.е. зависимости
Теоретический и экспериментальный анализы показали, что пики на дифференциальных кривых плавления связаны с выплавлением в интервале в несколько десятых градуса отдельных участков молекулы с характерным размером в несколько сотен пар оснований. Кольцевые замкнутые ДНК, фигурирующие в фагах и бактериях, вследствие топологической невозможности свободного раскручивания с образованием петель, должны плавиться при более высокой температуре, чем те же ДНК в линейной форме. Денатурация - процесс обратимый, последующее восстановление двухцепочечной структуры ДНК может происходить даже при полном расхождении цепей. Процесс воссоединения, называемый ренатурацией или отжигом, происходит при понижении температуры или рН. Если температура или рН понижаются постепенно, то цепи соединяются правильно, с восстановлением всех исходных пар оснований. При резком понижении температуры или рН правильное воссоединение затрудняется из-за спаривания оснований локально комплементарных участков в пределах одной или разных цепей. Как и в случае ДНК, двухцепочечные участки в РНК разрушаются при повышении температуры или рН, но, в отличие от ДНК, при высоких значениях рН в РНК разрушаются и фосфодиэфирные связи. Поскольку протяженность спирализованных участков в одноцепочечной РНК невелика, а сами спирали несовершенны, разрушаются они довольно легко. Однако полностью комплементарная двухцепочечная РНК плавится в довольно узком температурном интервале, как и двухцепочечная ДНК.
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 577; Нарушение авторского права страницы
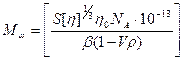
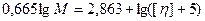
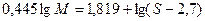
 Молекулы, которые не могут быть совмещены со своим зеркальным изображением, называются хиральными. Один из источников хиральности -образование спиральных конформаций. Спираль может быть правой и левой. В растворе хиральных молекул электромагнитное излучение, поляризованное по кругу вправо или влево, имеет различные показатели преломления и различные молярные коэффициенты экстинкции (коэффициенты пропорциональности между оптической плотностью с одной стороны, и толщиной поглощающего слоя и концентрацией вещества с другой). Различие молярных коэффициентов экстинкции для излучения, поляризованного по кругу вправо ε R и влево ε L, называется круговым дихроизмом. Коэффициент дихроичного поглощения:
Молекулы, которые не могут быть совмещены со своим зеркальным изображением, называются хиральными. Один из источников хиральности -образование спиральных конформаций. Спираль может быть правой и левой. В растворе хиральных молекул электромагнитное излучение, поляризованное по кругу вправо или влево, имеет различные показатели преломления и различные молярные коэффициенты экстинкции (коэффициенты пропорциональности между оптической плотностью с одной стороны, и толщиной поглощающего слоя и концентрацией вещества с другой). Различие молярных коэффициентов экстинкции для излучения, поляризованного по кругу вправо ε R и влево ε L, называется круговым дихроизмом. Коэффициент дихроичного поглощения:  Δ ε = ε L - ε R является функцией длины волны.
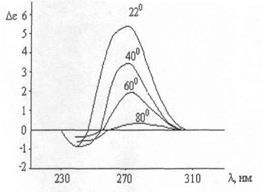
Δ ε = ε L - ε R является функцией длины волны.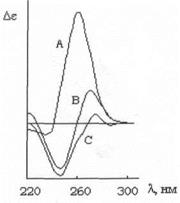 КД ДНК имеет положительный максимум при 273 нм и отрицательный - при 243 нм. При нагревании раствора КД начинает увеличиваться, проходит через максимум при 45° и затем падает до величины меньше первоначальной при 80 (рис.1). Это превращение при охлаждении обратимо [45].
КД ДНК имеет положительный максимум при 273 нм и отрицательный - при 243 нм. При нагревании раствора КД начинает увеличиваться, проходит через максимум при 45° и затем падает до величины меньше первоначальной при 80 (рис.1). Это превращение при охлаждении обратимо [45].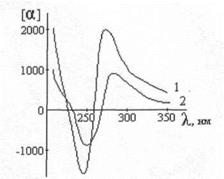
 Биополимеры обладают естественной оптической активностью, т. е вращают плоскость поляризации света.
Биополимеры обладают естественной оптической активностью, т. е вращают плоскость поляризации света.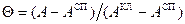 ,
, 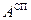 и
и 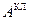 обозначено поглощение ДНК в полностью спиральном и полностью клубкообразном состоянии соответственно.
обозначено поглощение ДНК в полностью спиральном и полностью клубкообразном состоянии соответственно.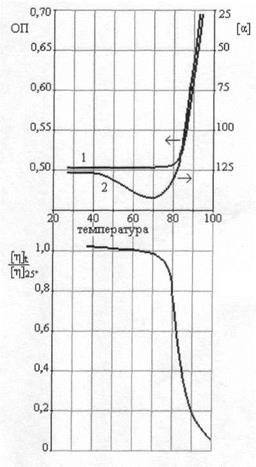
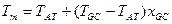
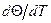 . Пример такой дифференциальной кривой приведен на рис.5. Конкретный профиль плавления, который отражают такие кривые, определяется последовательностью оснований в исследуемой ДНК.
. Пример такой дифференциальной кривой приведен на рис.5. Конкретный профиль плавления, который отражают такие кривые, определяется последовательностью оснований в исследуемой ДНК.