|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Природа межнуклеотидных связей
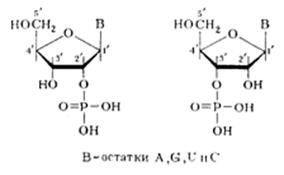
Работы по определению способа соединения нуклеотидов в полимерных молекулах НК были успешно завершены в начале 50-х годов сразу после того, как была установлена структура нуклеотидов и изучены некоторые свойства их производных (главным образом эфиров). К этому же времени были разработаны методы выделения и очистки ДНК и РНК, так что исследование природы межмономерных связей проводилось с использованием чистых, хотя и сильно деградированных препаратов НК. Первые сведения о типе межмономерной, или, как ее принято называть, межнуклеотидной связи были получены с помощью потенциометрического титрования. Эти сведения свидетельствовали о наличии, как в РНК, так и в ДНК только одной гпдроксильной группы у каждой фосфатной группы (рКа~1). На основании этого было сделано заключение, что НК содержит структурную единицу дизамещенной фосфорной кислоты. Естественно было предположить, что фосфатные остатки «сшивают» нуклеозиды за счет двух своих гидроксилов, а один остается свободным. Оставалось выяснить, какие части нуклеозидных фрагментов участвуют в образовании связи с фосфатными группами. Поскольку НК могут быть дезаминированы действием азотистой кислоты, очевидно, что аминогруппы пиримидиновых и пуриновых оснований не принимают участия в образовании межнуклеотидной связи. Помимо этого потенциометрическое титрование указывало, что и оксо (окси)-группы остатков гуанина и урацила, входящих в состав НК, свободны. На основании этих данных было сделано заключение о том, что межнуклеотидные связи образованы фосфатной группой и гидроксильными группами углеводных остатков (т. е. что они являются фосфодиэфирными), которые, следовательно, и являются ответственными за образование полимерной цепи (НК). Таким образом, то, что принято обычно называть межнуклеотидной связью, представляет собой по существу узел, включающий систему связей:
(где С — первичный или вторичный атомы углерода остатка углевода). При гидролизе ДНК и РНК в зависимости от условий реакции, образуются нуклеотиды с разным положением фосфатного остатка:
Если предположить, что в НК все межнуклеотидные связи идентичны, то, очевидно, что они могут включать помимо фосфатного остатка только 3'-гидроксильную группу одного нуклеозидного звена и 5'-гидроксильную группу другого нуклеозидного звена. В случае же их неравноценности в полимерной цепи ДНК могли бы одновременно существовать три типа связей: 3'—5', 3'—3' и 5'—5'. Для РНК за счет участия 2'-гидpoкcильнoй группы число типов связи должно было быть еще больше. Установить истинную природу межнуклеотидных связей в нативных ДНК и РНК удалось в результате направленного расщепления биополимеров с помощью химического и ферментативного гидролиза и последующего выделения и идентификации полученных при этом фрагментов.
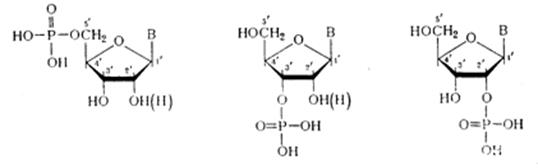
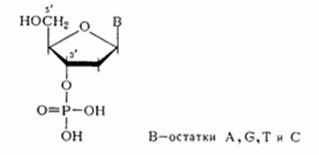
Межнуклеотидная связь в ДНК Химический гидролиз ДНК как метод деградации полимера с целью установления природы межнуклеотидной связи оказался практически непригодным. ДНК не расщепляется при щелочных значениях рН, что хорошо согласуется с предположением о фосфодиэфирной природе межнуклеотидной связи (устойчивость диалкилфосфатов в щелочной среде обсуждалась в разделе). При обработке кислотой даже в мягких условиях ДНК расщепляется как по фосфодиэфирным, так и по N-гликозидным связям, образованным пуриновыми основаниями. Вследствие этого расщепление полимера протекает неоднозначно, но из продуктов кислотного гидролиза ДНК все же удалось выделить дифосфаты пиримидиновых дезоксинуклеозидов, которые оказались идентичными синтетическим 3', 5'-дифосфатам дезоксицитидина и дезокситимидина:
Здесь же важно отметить, что наличие этих соединений в продуктах деградации ДНК указывает на участие обеих гидроксильных групп, по крайней мере, пиримидиновых мономерных компонентов, в образовании межнуклеотидной связи. Более специфическим оказалось ферментативное расщепление ДНК. При обработке препаратов ДНК фосфодиэстеразой (ФДЭ) змеиного яда полимер практически полностью гидролизуется до дезоксннуклеозид-5'-фосфатов, структура которых была Остановлена сравнением с соответствующими нуклеотидами, полученными встречным синтезом.
Эти данные свидетельствуют об участии 5'-гидроксильных групп всех четырех дезоксинуклеозидов, входящих в состав ДНК, в образовании межнуклеотидной связи. Аналогично, но до 3'-фосфатов дезоксинуклеозидов расщепляется ДНК в присутствии ФДЭ, выделенной из микрококков или из селезенки.
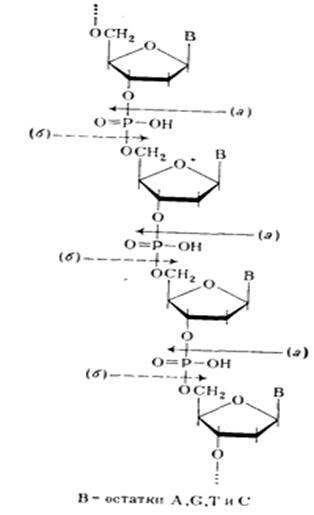
Из данных гидролиза ДНК фосфодиэстеразами различной специфичности становится очевидным, что связь нуклеозидных остатков в ДНК осуществляется фосфатной группой, которая одновременно этерифицирует гидроксильную группу у вторичного атома углерода (положение 3') одного нуклеозидного звена и гидроксильную группу у первичного атома углерода (положение 5') другого нуклеотидного звена. Таким образом, было убедительно доказано, что в ДНК межнуклеотидная связь осуществляется за счет фосфатной группы, а также 3'- и 5'-гидроксильных групп нуклеозидных остатков [(а) и (б) — направления расщепления полинуклеотидной цепи ДНК фосфодиэстеразами соответственно змеиного яда и селезенки или микрококков]:
Предположение о возможности иного строения полимера с регулярно перемежающимися связями нуклеозидных остатков по типу 3'—3' и 5'—5' было отвергнуто, так как оно не удовлетворяло всем экспериментальным данным. Так, полимер такого типа не должен был бы полностью гидролизоваться (до мономеров) в присутствии ФДЭ змеиного яда, избирательно расщепляющей только алкиловые эфиры нуклеозид-5'–фосфатов. То же можно сказать о ФДЭ селезенки, селективно гидролизирующей алкиловые эфиры нуклеозид-3'-фосфатов. Конформационный анализ ДНК Молекула ДНК обычно находится в форме двойной спирали, образуемой двумя полинуклеотидными цепями, обвивающимися одна вокруг другой. Два дезоксирибозных остова, расположенные по периферии молекулы, имеют антипараллельную ориентацию. Наиболее часто встречается В-форма.
Основная причина различия конформаций связана с геометрией сахарного кольца у форм В- и А-семейств. Фуранозное кольцо рибозы и дезоксирибозы неплоское. Обычно атом С2, либо атом СЗ выходят из Главное структурное отличие А-формы от В-формы - это большое отверстие в центре спирали у А-формы, которое занимает почти половину диаметра всей спирали (d~20Å, у В-формы d~9Å ). Молекулярным «переключателем» для перехода двойной спирали ДНК между семействами форм В- и А- служит сахарное кольцо пентозы.
Необычные структуры ДНК К ним относятся крестообразные структуры, левозакрученная Z-форма (для нее характерно чередование пуриновых и пиримидиновых нуклеотидов), различного рода тройные и четвертные спирали, структура со сдвинутыми петлями SLS [7]. Важное значение имеет проблема функционирования в клетке сверх скрученных кольцевых замкнутых ДНК. Для закрученного нормального двуспирального комплекса общее число оборотов α одной нити относительно другой равно числу витков β в ненапряженной двойной спирали плюс число супервитков τ, т.е. α =β +τ Величина β всегда положительна, τ может быть и отрицательным - это лево спиральные супервитки, противоположные по знаку двойные правые спирали. Различные характеристики ДНК зависят от плотности супервитков (степени суперспиральности) σ, равной числу этих супервитков τ на 10 пар оснований в состоянии, когда все пары в двойной спирали закрыты. Для природных кольцевых молекул ДНК вирусов α, чаще всего отрицательна. Наиболее типичное значение -σ =0, 05 (-5 супервитков на 1000 пар оснований). С ростом плотности сверхспиральности σ вероятность раскрытия пары возрастает, причем наиболее резко в области изменения плотности от 0, 06 до 0, 12. Здесь начинают раскрываться не только отдельные пары, но и более протяженные области, что приводит к появлению крестообразных структур. Плотность сверхспирализации не может превысить некоторого предельного значения (σ =0, 085), поскольку дальнейшее увеличение -σ будет компенсироваться соответствующим раскрытием пар оснований в двойной спирали [7]. Состояние сверхспирализации наблюдается не только у кольцевых ДНК, но также на отдельных участках линейной ДНК. Сверхспирализация отдельных доменов происходит под влиянием специальных белков, активирующих фермент ДНК-гиразу, который превращает «расслабленную» не сверхспирализованную ДНК в сверхспираль. Имеются данные о том, что транскрипция в клетке идет на ДНК, находящейся в напряженном сверхспирализованном состоянии. Н-форма связана с существованием тройного комплекса, состоящего из двух полипиримидиновых (С-Т) цепей и только одной полипуриновой (A-G) цепи. Структура со сдвинутыми петлями была обнаружена для ДНК и РНК. SLS-форма образуется, если в цепях ДНК содержатся короткие прямыеповторы. Она получается при проскальзывании относительно друг друга комплементарных нуклеотидных цепей. При этом возникают две однонитевые петли, выступающие из каждой полинуклеотидной цепи и разделенные длиной повтора [7]. В растворе двухнитчатая ДНК имеет специфическую третичную структуру. В присутствии полиэтиленгликоля ДНК приобретает компактную форму тора. При этом наблюдается изменение КД (кругового дихроизма). По-видимому, одновременно происходит агрегация молекул ДНК. Диаметры торов варьируют от 80 до 140 нм. Образование тора есть обратимый конформационный переход [4].
Физические модели ДНК Существуют две физические модели ДНК: модель гибкого стержня и модель спираль-клубок. В обычных полимерных цепях гибкость определяется поворотной изомерией. Однако в модели, где ДНК представляется в виде упругого стержня, ее жесткость характеризуется двумя параметрами: жесткостью на изгиб оси и жесткостью кручения. Жесткость на изгиб оси двойной спирали определяет способность ДНК укладываться в вирусных частицах, хромосомах и т.д. Энергия изгиба цепочки, состоящей из N+1 звеньев или N+1 пар оснований, с углами
где
где l - длина звена, а - персистентная длина, kT - средняя энергия колебаний. Отсюда, зная персистентную длину a=50, 0 нм, можно найти В модели гибкого стержня изменения конформации ДНК связываются с небольшим изгибом и кручением двойной спирали в каждой паре оснований, которые накапливаются на большой длине. В результате происходит непрерывное изгибание молекулы ДНК с образованием кольца диаметром около 10, 0 нм. Согласно другой точке зрения, укладка двойной спирали при компактизации ДНК может происходить за счет редких изломов на большой угол вследствие нарушения межплоскостных (стекинг) взаимодействий между парами оснований. Наряду с флуктуационными изгибами оси спирали и поворотами соседних пар оснований в ДНК может происходить и раскрытие отдельных пар оснований. Хотя этот процесс идет с очень малой вероятностью, тем не менее, он приводит к сильным изменениям конформации и играет определяющую роль в реакциях ДНК с химическими агентами (например, с формальдегидом), которые могут реагировать лишь с раскрытыми нуклеотидами. Эти маловероятные, но значительные изменения структуры учитываются в моделях спираль-клубок. При обычных физиологических температурах в ДНК происходят в основном флуктуационные раскрытия отдельных пар оснований с вероятностями не более 10-5. Вероятности одновременного раскрытия двух и большего числа пар оснований соответственно на один и два порядка меньше.
Третичная структура ДНК ДНК имеют линейную, кольцевую, 2-х и 1-о цепочечную формы.
Третичная структура ДНК у эукариотических клеток отличается тем, что многократная спирализация ДНК сопровождается образованием комплексов с белками. 46 хромосом (хроматид) человека организованы в 23 пары. Средняя длина хромосомы составляет 130 млн. пар оснований и имеет длину 5 см. Хромосома №1- 263 млн. п.о., хромосома № 46 –меньше 50 млн. п.о. Если проложить все ДНК в В-конформации в линию, то их общая длина превысит 2 метра. Человеческая хромосома 16 имеет 2, 5 мкм в длину, а длина самой ДНК- 3, 7 см. Понятно, что уместить такой длины ДНК в ядре возможно только путем ее определенной упаковки. При образовании третичной структуры ДНК человека происходит в среднем уменьшение ее размеров в 100 тысяч раз.
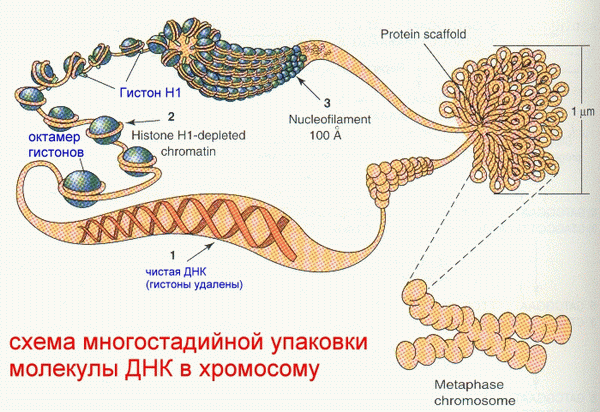
При образовании третичной структуры нуклеиновых кислот возможно образование трех и четырех нитевых участков. Образование трех нитевых участков благодаря так называемым Хугстэновским взаимодействиям (Hoogsteen base pairs), когда одновременно взаимодействуют три основания: А-А-Т, Т-А-Т, Г-Г-Ц, Ц-Г-Ц. Аналогичным образом происходит образование тетрамерных участков ДНК. Точный биологический смысл появления трех и четырех нитевых участков ДНК пока не выяснен. Имеются только лишь предположения о том, что такие участки возникают в местах, наиболее ответственных за процессы репликации и транскрипции. Материал хромосом – хроматин – содержит кроме самой ДНК также гистоны, негистоновые белки, небольшое количество РНК. Нуклеосомный кор содержит октамер гистонов (2 х (Н2а+Н2b+H3+H4)). Гистон – простой белок (примерно 50% хроматина). Нуклеосомный корпус образуется при оборачивании октамера гистонов двунитевой спирализованной ДНК на 1, 5 оборота, отдельно включается дополни-тельный белок – гистон Н1. Все вместе называется хроматосомом.
Хроматосомы образуются на двунитевой спирали ДНК на дистанциях от 20 до 90 пар нуклеотидов и напоминают нанизанные на нитку бусины. Следующий этап- сворачивание в спираль очень длинной последовательности “бус”. Эта спираль, в свою очередь, претерпевает сворачивание в двужильные канаты, из которых образуются гроздья, являющиеся небольшой частью хромосомы:
Межнуклеотидная связь в РНК Уже на первых этапах изучения строения РНК был установлен факт чрезвычайной неустойчивости се при щелочном гидролизе. Основными продуктами щелочного гидролиза РНК являются рибонуклсозид-2'- и рибонуклеозид-3'-фосфаты, образующиеся практически в равных количествах.
Рибонуклеозид-5'-фосфаты при этом не образуются. Эти данные не укладывались в представления о фосфодиэфирной природе межнуклеотидной связи в РНК и требовали всестороннего изучения. Очень важную роль в таком исследовании, которое выполнили в начале 50-х гг. Тодд с сотрудниками, сыграли синтетические алкиловые эфиры рибонуклеотидов, которые были получены специально, чтобы промоделировать тот или иной тип фосфодиэфирной связи. Полученные школой Тодда данные о механизмах превращения алкиловых эфиров рибонуклеотидов в щелочной среде позволили предположить, что в РНК, так же как и в ДНК, межнуклеотидная связь осуществляется фосфатной группой и 3'- и 5'-гидроксильными группами углеводных остатков. Подобная связь в РНК должна очень легко расщепляться в щелочной среде, так как соседняя 2'-гидроксильная группа должна катализировать этот процесс при рН> 10, когда начинается ионизация гидроксильных групп рибозы. Очень важно подчеркнуть, что промежуточными соединениями при щелочном расщеплении должны быть все четыре рибонуклеозид-2', 3'-циклофосфата, а конечными — образующиеся при их гидролизе рибонуклеозид-3'-фосфаты и рибонуклеозид-2'-фосфаты (четыре пары изомеров). Данные щелочного гидролиза ограничили количество возможных для РНК типов межнуклеотидных связей, но не прояснили вопроса о том, как построен этот полимер. Более точные сведения о типе межнуклеотидной связи в РНК, как и в случае ДНК, были получены с помощью ферментативного гидролиза. Гидролиз РНК с использованием ФДЭ змеиного яда, протекающий до рибонуклеозид-5'-фосфатов, подтвердил уже прямым путем предположение об участии 5'-гидроксильных групп в образовании фосфодиэфирной связи между мономерными звеньями. Позднее это было окончательно установлено в результате открытия фосфоролиза РНК в присутствии фермента полинуклеотидфосфорилазы (ПНФаза), приводящего к образованию рибонуклеозид-5'-пирофосфатов:
Таким образом, оставалось выяснить природу второй гидроксильной группы, участвующей в образовании межнуклеотидной связи. Частично решить эту задачу помог еще один фермент, который использовался для направленного расщепления РНК, — пиримидиловая рибонуклеаза (РНаза). Ранее было показано, что этот фермент расщепляет только алкиловые эфиры пиримидиновых рибонуклеозид-3'-фосфатов до рибонук-леозид-3'-фосфатов (через промежуточный рибонуклеозид-2', 3'-циклофосфат). Оказалось, что аналогичным образом этот фермент действует и на РНК. В экспериментах с любыми образцами очищенной РНК было обнаружено, что количество фосфорной кислоты, которая образуется при обработке полимера последовательно пиримидиловой РНазой и фосфомоноэстеразой (ФМЭ), а также количество иодной кислоты, затрачиваемой на последующее окисление, эквивалентно количеству остатков пиримидинов в данном образце РНК. Это говорило в пользу того, что, по крайней мере, пиримидиновые нуклеотиды в РНК связаны с соседними нуклеотидами только посредством 3'—5'-межнук-леотидной связи. Этот вывод подтверждают данные щелочной обработки ферментативных гидролизатов РНК, полученных после действия на нее РНК-азы: в щелочной среде миграция фосфатного остатка в рибонуклеозид-3'- и -2'-фосфатах невозможна, и наличие в соответствующих гидролизатах только пиримидиновых рибонуклеозид-З'-фосфатов делает очевидным 3'—5'-тип межнуклеотидной связи для пиримидиновых нуклеотидов. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 919; Нарушение авторского права страницы



 плоскости других четырех атомов
плоскости других четырех атомов между звеньями цепи имеет вид
между звеньями цепи имеет вид
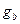 константа жесткости на изгиб двойной оси.
константа жесткости на изгиб двойной оси.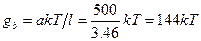
 Дж.
Дж.  6эВ.Жесткость на кручение или торсионная жесткость ДНК, определяется как изменение энергии при отклонении угла между соседними парами оснований от его равновесного значения. Согласно расчетам, числовое значение торсионной жесткости
6эВ.Жесткость на кручение или торсионная жесткость ДНК, определяется как изменение энергии при отклонении угла между соседними парами оснований от его равновесного значения. Согласно расчетам, числовое значение торсионной жесткости  для кольцевой полимерной цепи составляет
для кольцевой полимерной цепи составляет  Двухцепочечные ДНК с " липкими" концами могут образовывать кольцо, которое далее ковалентно сшивается по сахарофосфатной цепи при помощи ДНК-лигазы.
Двухцепочечные ДНК с " липкими" концами могут образовывать кольцо, которое далее ковалентно сшивается по сахарофосфатной цепи при помощи ДНК-лигазы.

 Н1 – очень богат ЛИЗ; Н2а, Н2b – умеренное количество ЛИЗ; Н3 – есть ЦИС, умеренно - АРГ; Н4 – богат АРГ и ГЛИ.
Н1 – очень богат ЛИЗ; Н2а, Н2b – умеренное количество ЛИЗ; Н3 – есть ЦИС, умеренно - АРГ; Н4 – богат АРГ и ГЛИ.