|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Термодинамика плавления двойной спирали (переходов спираль - клубок)
Рассмотрим простую двойную спираль, построенную из двух комплементарных гомополинуклеотидных цепей. В области перехода спираль-клубок такая молекула состоит из чередующихся спиральных и неупорядоченных участков — петель. Если обозначить число разорванных пар N1, число связанных пар N2 и число двуспиральных участков, равное числу петель, n, то свободная энергия системы запишется в виде:
где G1 и G2 - свободные энергии полностью разделенных и полностью двуспиральных молекул, отнесенные к одной паре оснований, GS - свободная энергия, необходимая для возникновения петли, т. е. расплавленной области между двумя спиральными. S0 энтропия смешения спиральных и не спиральных участков, т. е. (в расчете на моль)
Минимум G при данной температуре соответствует условию
где
Таким образом, гомополимер в области перехода является последовательностью спиральных и не спиральных участков, размеры которых определяются σ . Это связано с одномерностью системы, которая, согласно теореме Ландау и Лившица, не может разделиться на фазы. Кривая плавления, т. е. зависимость доли неупорядоченных пар При σ =1 кооперативность отсутствует; при σ =0 кооперативность полная. В середине интервала плавления модуль производной
Расчет, основанный на модели Изинга, дает:
где Δ H - разность энтальпий спиральной и не спиральной молекул в расчете на пару оснований. Из экспериментальных значений Δ T для синтетических гомополинуклеотидов получается σ ~10-4-10-5, т.е. GS≈ 30 кДж/моль. Степень кооперативности очень высока [4].
Рассмотрение гетерогенности требует учета двух факторов - различий стабильности пар AT и GC и добавочного укрепления или ослабления двойной спирали лигандами. Эти факторы влияют по-разному, т.к. лиганды могут перераспределяться по цепи в процессе плавления, а первичная структура неизменна. Допустим, что m1 молекул лиганда связано не спиральными и m2 спиральными участками полимера. Тогда где g1 и g2 - свободные энергии лигандов соответственно в не спиральных и спиральных участках. Последний член содержит энтропию смешения полимера с лигандами. Условие минимума ∂ G/∂ n=0 такое же, как и в предыдущем случае, т.е. средняя длина спиральных участков при данном Θ не зависит от присутствия лигандов. С другой стороны, из условия ∂ G/∂ N1=0 получается:
где c1=m1/N1, c2=m2/N2. Эти концентрации лигандов выражаются через концентрацию лиганда в растворе c0 и константы связывания K1 и K2
Кривая плавления гомополиера без лигандов описывается функцией Θ =f(s), причем Θ =0, 5 при sпл=1, т.е. G1=G2. При наличии лигандов Θ =f(s*) и Θ =0, 5 при s*, т.е. при
Сдвиг температуры плавления по сравнению с таковой для чистого полимера T0 определяется как
Изменение интервала температур плавления есть
Полная концентрация лигандов в растворе и на полинуклеотиде равна
где р - концентрация связывающих лиганды фосфатных групп полинуклеотида. Если с » р, то
При прочном связывании лигандов полимером во всей области перехода, т. е. при K1p »1 или K2p »1 и c ≈ p
где q=K2/K1. Такимобразом, лиганды действуют в качестве «скрепок», стабилизирующих двойную спираль [4]. Особенность гетерополимера состоит в относительно малом числе микросостояний, отвечающих данной энергии. Поэтому энтропия смешения не может существенно влиять на плавление. Вместе с тем появляется новый энергетический фактор, определяемый тем, что при уменьшении средней длины расплавленных участков содержание в них более стабильных пар GC должно уменьшаться. Конкуренция этого фактора и фактора, обусловленного невыгодностью «стыков» спиральных и не спиральных участков, должна приводить к чередованию спиральных и не спиральных участков определенной длины при данном значении Θ [4]. Приближенное решение задачи о плавлении гетерополимера с беспорядочной последовательностью пар получено Лазуркиным и Франк-Каменецким. Они разделили молекулу на одинаковые отрезки, каждый из которых содержит λ пар. Если λ достаточно велико, то распределение концентрации пар GC по этим отрезкам будет гауссовым, т. е.:
где Общее число пар GC в расплавленных участках минимально, если все отрезки с содержанием этих пар, меньшим некоторого предельного значения xλ расплавлены и все отрезки с x > xλ спиральны. Значение xλ определяется условием:
Средняя концентрация GC в расплавленных участках равна
Так как состав каждого отрезка не зависит от состава других, вероятность того, что расплавленная область состоит из r отрезков, следующих друг за другом, равна
и среднее число отрезков расплавленной области
Среднее число пар оснований в расплавленной области
и число расплавленных областей во всей молекуле
Свободная энергия полимера
где GS - энтропийный вклад, определяемый смешением областей. Вводя условия N1+N2 =N, < x1> N1+< x2> N2=x0N, выражая n через λ, обозначая Δ GGC=G1GC-G2GC, Δ GAT=G1AT-G2AT и отбрасывая постоянные члены, получим
Равновесные значения N1 и λ находятся из условия минимума выражения G(N1, λ ).
Интервал плавления гетерополимера
где TGC и TAT - температуры плавления для соответствующих полимеров. Средняя длина спиральной области:
Определения оптической плотности (и, следовательно, Θ ) и размеров молекулы ДНК в растворе путем измерения характеристической вязкости в процессе плавления позволили найти средние длины спиральных участков: 1000-2500 пар при Θ =0, 8-0, 9 и 400-500 пар при Θ =0, 5. Ширина интервала плавления молекул ДНК со случайной последовательностью нуклеотидов составляет 2, 5-3°С. ДНК бактерий - около 5°С, ДНК высших организмов - около 10°С, что свидетельствует об их блочном строении. Молекулярная масса блоков (5-15)· 109 [4]. Процессинг ДНК и РНК
1.Репликация (от ДНК к ДНК) 2.Транскрипция (от ДНК к РНК) 3. Трансляция (от РНК к белку) 4.Обратная транскрипция (от РНК к кДНК) Долгое время считалось, что передача информации от РНК к ДНК невозможна, однако, впоследствии выяснилось, что это не так. Некоторые вирусы способны встраивать информацию со своей вирусной РНК в ДНК генома клетки-хозяина. Возможность " обратного" направления информации в настоящее время все шире используется в различных целях, от исследовательских до терапевтических. Так называемые энзимы - реверс-транскриптазы - способны осуществлять синтез ДНК на матрице РНК. О происходящих в клетках млекопитающих (эукариот) процессах передачи информации известно достаточно много, но далеко не все, и изложение хотя бы известных на данный момент времени сведений потребовалось бы слишком много места. Поэтому далее будут изложены лишь самые основы протекающих в клетках простейших организмов (прокариот) этапов передачи наследственной информации.
Репликация В процессе копирования информации происходит синтез дочерних молекул ДНК на основе информации, " записанной" в родительской молекуле ДНК. Ясно, что дочерние молекулы должны представлять собой точные копии родительской. Репликация может осуществляться тремя способами: а) консервативным; б) полуконсервативным; в) дисперсивным. При консервативной репликации вновь синтезированные цепи ДНК находятся в дочерней молекуле. При полуконсервативной репликации полученные молекулы состоят из родительской и вновь синтезированной цепей. Дисперсивный способ репликации означает наличие перемежающихся родительских и вновь синтезированных участков на каждой из цепей образованных молекул ДНК. Для животных организмов и человека характерен только полуконсервативный путь репликации ДНК. В процессе репликации участвует целый ряд энзимов (ферментов) с определенными функциями. Только синтезирующих ферментов в клетках прокариот насчитывается три. Их называют ДНК - полимеразами I, II и III. Сведения о функциональных особенностях ДНК-полимераз приведены в таблице.
Фрагмент Кленова – результат частичного протеолиза ДНК-полимеразы I E. Coli субтилизином. Основная функция полимеразы III – синтез цепи, полимеразы I – синтез и исправление ошибочно вставленных нуклеотидов. Полимераза II осуществляет особые, специализированные функции. Репликация начинается с расплетания цепей ДНК специальными расплетающими белками, которые называют геликазами (или Rep-протеином). Геликазы используют энергию АТФ в процессе расплетания цепей. Скорость расплетания составляет около 6000 мин-1. Для того чтобы расплетенные цепи не могли вновь соединиться, имеются специальные SSB-белки (single-strand binding proteins), которые присоединяются к комплементарным цепям, удерживая их от ассоциации. По мере продвижения репликационной вилки SSB-протеины передвигаются по цепи, диссоциируя с одного места и присоединяясь на другом. Этот процесс не требует затрат энергии АТФ. После освобождения достаточного места начинается синтез праймера - затравки, необходимой для работы ДНК-полимеразы. Наличие затравки является необходимым условием функционирования ДНК-полимераз (как и наличие комплементарной цепи). В качестве затравки на каждой из разделенных цепей синтезируются маленькие отрезки молекул РНК при помощи фермента примазы. Синтез новой цепи ДНК осуществляется всегда в направлении 5'-3', поэтому если по одной матричной цепи возможен непрерывный синтез, то по комплементарной ей цепи синтез осуществляется только участками. Эти участки синтеза называют фрагментами Оказаки. Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента, РНК-овый праймер удаляется имеющейся у полимераз 5'-3' полимеразной активностью и достраивается дезоксирибонуклеотидами. После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи фермента ДНК-лигазы.
Частота возникновения ошибок при репликации и транскрипции не превышает 10-8-10-9, то есть, возможна лишь одна ошибка на сотни миллионов нуклеотидов. Такая точность не может быть обеспечена одним только лишь правилом комплементарности нуклеотидов (обеспечивающим точность 1: 10000-1: 100000). Репликационный аппарат имеет собственные механизмы " поддержания точности" копирования генетической информации. Этими функциями наделена ДНК-полимераза I. Модель ее структуры и функциональных участков показана на рисунке. Она имеет три зоны активности: полимеризующую в направлении 5'-3', и экозонуклеазные в направлениях 5'-3' и 3'-5'. Области активности разделены пространственно. Вперед (по ходу продвижения полимеразы по матричной цепи ДНК) обращена зона 5'-3' экзонуклеазной активности. Она служит для удаления попадающихся на пути РНК-овых праймеров (затравок). Далее идет собственно синтетическая зона и, наконец, зона с экзонуклеазной активностью в направлении 3'-5'. С этой зоной связана так называемая PROOF-READING активность (способность узнавать неправильно встроенные нуклеотиды) и исправлять их вырезанием ряда уже встроенных нуклеотидов. Для этого молекула ДНК-полимеразы смещается (не отсоединяясь от ДНК-овой матрицы) к месту синтеза и последовательно вырезает нуклеотиды, после чего возобновляется нормальный синтез.
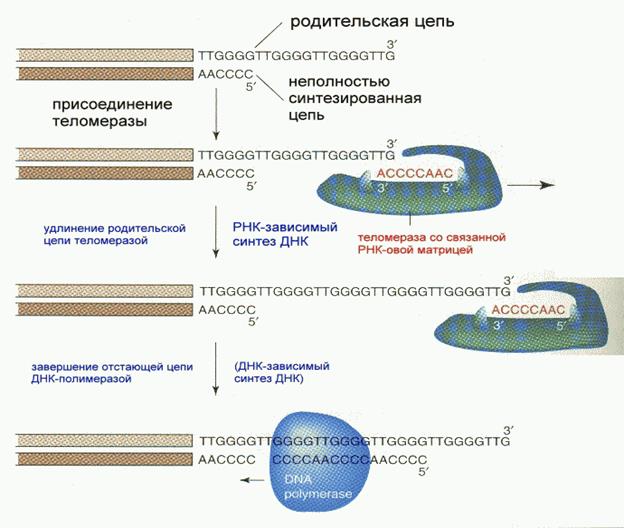
В процессе репликации одна из вновь синтезированных цепей (а именно- синтезированная на лидирующей цепи) получается короче на несколько десятков нуклеотидов вследствие того, что содержала РНК-овый праймер (затравку), впоследствии удаленный. Однако получение более коротких копий является совершенно недопустимым в процессе репликации явлением. Положение исправляется при помощи так называемых теломераз. Теломераза, содержащая в себе последовательность нуклеотидов, за несколько приемов удлиняет укороченную цепь, создавая пространство для работы примазы и ДНК-полимеразы, после чего избытки нуклеотидов удаляются:
Воздействие на организм неблагоприятных факторов (химические соединения, ультрафиолет и др.) приводит к постоянному накоплению ошибок в геноме, которые, в конечном итоге, вызывают появление патологии, в частности, невыясненный до сих пор механизм раковых заболеваний. Пока лишь существуют только предположения о том, что причиной раковых заболеваний являются дефекты в носителях информации- ДНК.
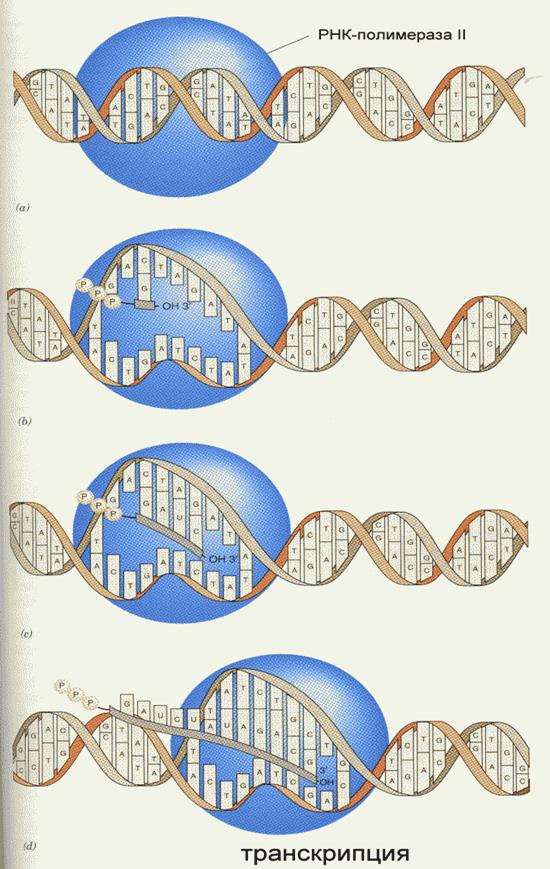
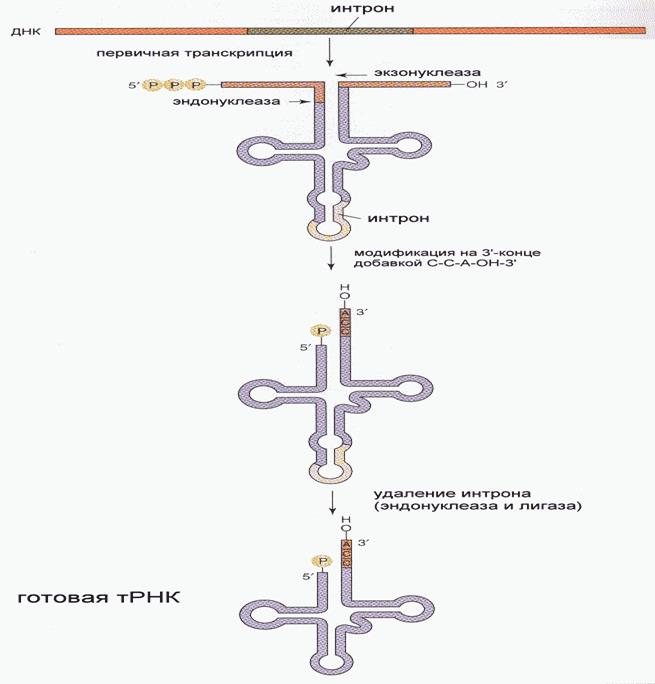
Транскрипция Транскрипция - синтез молекул РНК на основании информации, записанной в ДНК. Осуществляется в ядрах при участии ДНК-зависимых РНК-полимераз, существующих в типах I, II и III (в порядке выхода в гель-хроматографии). РНК-полимеразы I синтезируют рибосомальные РНК. РНК-полимеразы II синтезируют матричные и вирусные РНК. РНК-полимеразы III синтезируют транспортные РНК. В процессе транскрипции копируется не вся информация с ДНК, а только выборочная, часто отрезками. Сигналом для присоединения полимеразы служат так называемые промотеры, в районе которого (35 нуклеотидных пар до и 10 пар после него) и присоединяется РНК-полимераза. Происходит разделение цепей ДНК и начинается синтез молекулы РНК в направлении 5'-3', только на одной из цепей. При этом по месту тимидиновых нуклеотидов комплементарной цепи встают уридиловые нуклеотиды. Весь комплекс передвигается по молекуле ДНК, пока не будет закончен синтез требуемого участка РНК. ДНК с " отсканированной" информацией репарирует, ассоциируя в двухнитевые молекулы (смотри рисунок ниже). РНК-полимераза II очень чувствительна к некоторым соединениям, изменяющим ее активность. Так, сродство к альфа-аманитину (компонент грибного яда) составляет KL = 10-8–10-9M. Таким образом, аманитин является сильнейшим ингибитором РНК-полимеразы II, в результате при отравлении белой поганкой (достаточно 0, 5 г сырого гриба) вначале развивается расстройство желудочно-кишечного тракта, а через 48 часов наступает смерть в результате тяжелого поражения печени, вследствие прекращения синтеза требуемых белков (нет РНК). Терапия при этой патологии отсутствует, за исключением пересадки печени. Возбудитель туберкулеза Micobacterium tuberculosis (точнее, его РНК-полимераза) весьма чувствителен к антибиотику рифампицину, в то время как человеческая РНК-полимераза к нему мало чувствительна. На этом свойстве рифампицина основано его использование в терапии туберкулеза. Молекулы РНК очень часто претерпевают посттранскрипционную модификацию, заключающуюся в удалении участков построенной цепи. Наглядно это можно проследить на примере синтеза молекулы транспортной РНК:
Синтез белка Участие ДНК и РНК в синтезе белков– одна из основных функций нуклеиновых кислот. Белки – важнейшие компоненты каждого живого организма. Мышцы, внутренние органы, костная ткань, кожный и волосяной покров млекопитающих состоят из белков. Это полимерные соединения, которые собираются в живом организме из различных аминокислот. В такой сборке управляющую роль играют нуклеиновые кислоты, процесс проходит в две стадии, причем на каждой из них определяющий фактор – взаимоориентация азотсодержащих гетероциклов ДНК и РНК. Основная задача ДНК – хранить записанную информацию и предоставлять в тот момент, когда начинается синтез белков. В связи с этим понятна повышенная химическая устойчивость ДНК в сравнении с РНК. Природа позаботилась о том, чтобы сохранить по возможности основную информацию неприкосновенной. На первой стадии часть двойной спирали раскрывается, освободившиеся ветви расходятся, и на группах А, Т, Г и Ц, оказавшихся доступными, начинается синтез РНК, называемой матричной РНК, поскольку она как копия с матрицы точно воспроизводит информацию, записанную на раскрывшемся участке ДНК. Напротив группы А, принадлежащей молекуле ДНК, располагается фрагмент будущей матричной РНК, содержащий группу У, все остальные группы располагаются друг напротив друга в точном соответствии с тем, как это происходит при образовании двойной спирали ДНК.
По указанной схеме образуются полимерная молекула матричной РНК, содержащая несколько тысяч мономерных звеньев. На втором этапе матричная ДНК перемещается из ядра клетки в околоядерное пространство – цитоплазму. К полученной матричной РНК подходят так называемые транспортные РНК, которые несут с собой (транспортируют) различные аминокислоты. Каждая транспортная РНК, нагруженная определенной аминокислотой, приближается к строго обусловленному участку матричной РНК, нужное место обнаруживается с помощью все того же принципа взаимосоответствия групп А-У, и Г-Ц. В конечном итоге две аминокислоты, оказавшиеся рядом, взаимодействуют между собой, так начинается сборка будущей белковой молекулы.
Важная деталь состоит в том, что временное взаимодействие матричной и транспортной РНК проходит всего по трем группам, например, к триаде Ц-Ц-У матричной кислоты может подойти только соответствующая ей тройка Г-Г-А транспортной РНК, которая непременно несет с собой аминокислоту глицин. Точно также к триаде Г-А-У может приблизиться лишь набор Ц-У-А, транспортирующий только аминокислоту лейцин. Таким образом, последовательность групп в матричной РНК указывает, в каком порядке должны соединяться аминокислоты. Кроме того, система содержит в закодированном виде дополнительные регулирующие правила, некоторые последовательности из трех групп матричной РНК указывает на то, что в этом месте синтез белка должен остановиться, т.е. молекула достигла необходимой длины. Показанный на рис. 10 синтез белка проходит с участием еще одного – третьего вида РНК, они входят в состав рибосом и потому их называют рибосомными. Рибосома, представляющая собой ансамбль определенных белков рибосомных РНК, обеспечивает взаимодействие матричной и транспортной РНК, играя роль конвейерной ленты, которая передвигает матричную РНК на один шаг после того, как произошло соединение двух аминокислот. Основной смысл двух стадийной схемы состоит в том, что полимерная цепь белковой молекулы собирается из различных аминокислот в намеченном порядке и строго по тому плану, который был записан в закодированном виде на определенном участке ДНК. Таким образом, ДНК представляет собой отправную точку всего этого запрограммированного процесса. В процессе жизнедеятельности белки постоянно расходуются, и потому они регулярно воспроизводятся по описанной схеме, весь синтез белковой молекулы, состоящей из сотен аминокислот, проходит в живом организме приблизительно в течение одной минуты.
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 640; Нарушение авторского права страницы


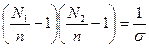
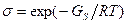 есть фактор кооперативности. Чем меньше σ, тем больше кооперативность. Равновесные значения N1, N2 и n находятся путем дифференцирования G по N1, причем N2 =N-N1, где N — константа. Приравняв ∂ G/∂ N1 нулю, получаем:
есть фактор кооперативности. Чем меньше σ, тем больше кооперативность. Равновесные значения N1, N2 и n находятся путем дифференцирования G по N1, причем N2 =N-N1, где N — константа. Приравняв ∂ G/∂ N1 нулю, получаем: 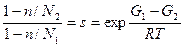
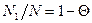 (Θ - степень спиральности) от T, идет тем круче, чем меньше σ.
(Θ - степень спиральности) от T, идет тем круче, чем меньше σ.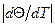 максимален и интервал определяется условием
максимален и интервал определяется условием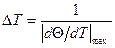 .
. ,
, 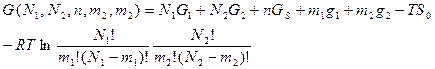
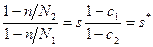 ,
, 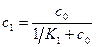 ,
, 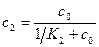 .
.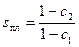 .
.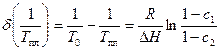 .
.
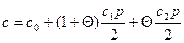 ,
, 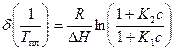 ,
,  .
.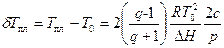 ,
, 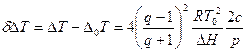
 ,
, 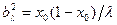 , x- содержание пар GC, x0- значение х, отвечающее максимуму распределения.
, x- содержание пар GC, x0- значение х, отвечающее максимуму распределения.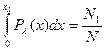
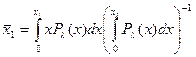

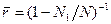 .
.
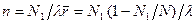

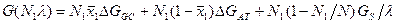

 .
.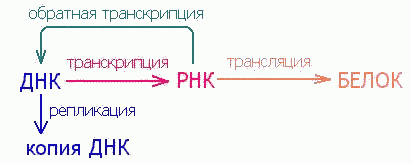 При жизни организма непрерывно происходят процессы обновления тканей, клеток и т.д., которые неизбежно включают процессы копирования и передачи информации, хранящейся в геноме. Направления передачи наследственной (генетической) информации выделяют в четыре группы:
При жизни организма непрерывно происходят процессы обновления тканей, клеток и т.д., которые неизбежно включают процессы копирования и передачи информации, хранящейся в геноме. Направления передачи наследственной (генетической) информации выделяют в четыре группы: