|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Модуль 2. Микроорганизмы и окружающая среда. Действие биологических и химических факторовСтр 1 из 24Следующая ⇒
СПЕЦГЛАВЫ МИКРОБИОЛОГИИ конспект лекций Учебно-методическое пособие Киров 2011
Содержание Модуль 1. «Основные механизмы обмена веществ и преобразования энергии у аэробных и анаэробных микроорганизмов. Типы брожения. Фотосинтез» 1. Лекция №1.1 «Общие представления об обмене веществ у микроорганизмов. Понятия анаболизма, катаболизма и метаболизма. Термодинамические закономерности обменных процессов у прокариот и эукариот. Понятия аэробиоза и анаэробиоза»....................................................4
2. Лекция №1.2«Конструктивный и энергетический метаболизм микробной клетки. Общие принципы биосинтеза макромолекул у микроорганизмов. Пути метаболизма, приводящие к образованию макроэргов» …………………………………………………………………...22
3. Лекция №1.3 «Дыхание, брожение и фотосинтез микроорганизмов. Методы исследования обмена веществ у микроорганизмов»………………………………………………………….…57 Слайд №11 СН3СН2ОН + 02 -> СН3СООН + Н20 (∆ G = -494 кДж/моль) Окисление этилового спирта уксуснокислыми бактериями может идти и дальше — до появления диоксида углерода и воды, при этом освобождается большое количество энергии: СН3СН2ОН + 302 -> 2С02 + 3H20 (∆ G = -1366 кДж/моль)
Анаэробное дыхание микроорганизмов. Осуществляется без участия молекулярного кислорода. Различают собственно анаэробное дыхание (нитратное, сульфатное) и брожение. При анаэробном дыхании акцептором водорода являются окисленные неорганические соединения, которые легко отдают кислород и превращаются в более восстановленные формы. Так проходят денитрификация и десульфофикация (сульфатредукция). Слайд №12 Нитратное дыхание — восстановление нитратов до молекулярного азота — проходит по схеме:
5С6Н1206 + 24КN03 -> 24КНС03 + 18Н20 + 12N2 + 6С02 (∆ G = -1760 кДж/моль) Сульфатное дыхание — восстановление сульфатов до сероводорода, сопровождающееся выделением такого же количества энергии: С6н12о6 + 3K2S04 -> зк2со3 + зсо2 + зн2о + 3H2S (∆ G = -1760 кДж/моль) Процесс восстановления нитратов до молекулярного азота индуцируется молибденсодержащим ферментом- нитратредуктазой. Если в среде содержится кислород, функция нитратредуктазы подавляется. Поэтому восстановление нитратов происходит только в анаэробных условиях.
Брожение Это процесс расщепления органических углеродсодержащих соединений в анаэробных условиях. Оно характеризуется тем, что последним акцептором водорода служит молекула органического вещества с ненасыщенными связями. Вещество при этом разлагается только до промежуточных продуктов, представляющих собой сложные органические соединения (спирты, органические кислоты). Заключенная в них энергия не используется микробами, а образовавшаяся в небольших количествах энергия выделяется в окружающую среду. Слайд №13 При брожении молекулы глюкозы (гексозы) освобождается 166 кДж/моль, что примерно в 17 раз меньше, чем при аэробном окислении того же вещества: С6Н1206 -> 2С2Н5ОН + 2С02 (∆ G = -166 кДж/моль) При окислении глюкозы различают три фазы: 1) гликолиз (путь Эмбдена—Мейергофа—Парнаса); 2) окислительное декарбоксилирование (цикл Кребса); 3)окислительное фосфорилирование (дыхательная цепь, в которой происходит перенос водорода и электронов).
Гликолиз Слайд№ (без №) Это фаза, общая для анаэробного и аэробного дыхания, две другие фазы наблюдаются только в аэробных условиях. Гликолиз представляет собой совокупность последовательно протекающих окислительно-восстановительных реакций (около 10), в результате которых одна молекула глюкозы расщепляется на две молекулы пировиноградной кислоты. Процесс проходит в анаэробных условиях. При гликолизе чистый выход АТФ составляет две молекулы и освобождаются четыре атома водорода: С6Н1206 → 2С3Н4О3 + 4Н+ + 2АТФ Глюкоза Пировино- Чистый выход градная кислота При дальнейшем превращении продуктов расщепления глюкозы в аэробных условиях (цикл Кребса, окислительное фосфорилирование) образуется энергия, которая идет на синтез АТФ. При полном окислении одной молекулы глюкозы выход АТФ составляет 38 молекул. В аэробных условиях количество энергии, образующейся в виде АТФ, в 19 раз больше, чем при анаэробном брожении. Поэтому с энергетической точки зрения анаэробное дыхание по сравнению с аэробным считается малоэффективным. Все виды брожений до образования пировиноградной кислоты протекают одинаково. Дальнейшее превращение пировиноградной кислоты зависит от свойств микроба. Гомоферментативные молочнокислые бактерии превращают ее в молочную кислоту, дрожжи — в этиловый спирт ит.д.
4. Фотосинтез Слайд №15 Фотосинтез — процесс, при котором происходит превращение световой энергии в химическую. Специальные пигменты микроорганизмов (цианобактерии) и растений с помощью солнечной энергии из диоксида углерода (СО2 ) и воды (Н20) образуют органические вещества и кислород, благодаря чему поддерживается жизнь на Земле. Фотосинтетический аппарат представлен тремя компонентами: антенной, РЦ (реакционный центр) и ЭТЦ (электронно-транспортная цепь). В антенне сосредоточены светособираюшие пигменты: бактериохлорофиллы и каротиноиды. Реакционный центр (РЦ) воспринимает световую волну длиной 870—875нм и переносит ее на бактериохлорофилл с электронно-транспортной цепи (ЭТЦ), в которой содержится несколько цитохромов. Фотосинтез может быть оксигенным и аноксигенным. Оксигенный фотосинтез среди прокариот осуществляют только цианобактерии и прохлорофиты. При этом из воды образуется кислород, который выделяется во внешнюю среду. В аноксигенном фотосинтезе участвуют особые пигменты: бактериохлорофилл А и реже бактериохлорофилл В, а также каротиноиды.
5. Метаногенез
Метанобразующие бактерии принадлежат к археобактериям. По форме — это палочки и кокки. Спор в чистых культурах не образуют. В стенках таких бактерий отсутствует пептидогликан, поэтому установить окраску по Граму невозможно. Облигатные анаэробы широко распространены в природе. Встречаются в преджелудках жвачных животных, а также во многих водоемах: прудах, озерах, затопляемых рисовых полях, отдельных районах Мирового океана. Обычно в таких местах происходит разложение органических веществ с образованием Н2, СО2 и других соединений. Метановые бактерии завершают процесс разложения растительных и животных остатков. Используют водород (Н2) для восстановления двуокиси углерода (С02) до метана. Метан — горючий газ, который получают из органических отходов и используют для обогрева теплиц, животноводческих помещений, населенных пунктов. Так, в Китае работает более 7 млн биогазовых заводов, в Индии — 75 тысяч гобар-заводов, перерабатывающих навоз. В Японии при переработке отходов получают до 4, 5млрд м3 метана. Таким путем не только осуществляется очистка окружающей среды, но и происходит накопление сравнительно недорогого топлива, которое можно использовать в сельскохозяйственном производстве.
6. Роль ферментов в обмене веществ у микроорганизмов Обшая характеристика ферментов В каждом организме (животном, растительном или микробной клетке) все время происходят сложные процессы превращения веществ. Важное значение в этих процессах играют ферменты (энзимы) — биологические катализаторы белковой природы, причем не только белковой. Ферменты микробов делят на эндо- и экзоферменты. Эндоферменты прочно связаны с цитоплазмой, осуществляют дальнейшее расщепление поступающих питательных веществ и превращение их в составные части клетки. Экзоферменты выделяются в окружающую среду, где производят превращение питательных веществ в более простые соединения, которые проходят через оболочку микробной клетки и служат пластическим материалом. В 1898 г. ученик Луи Пастера Эмиль Дюкло предложил называть ферменты по веществу, на которое они действуют, с добавлением окончания «-аза». Например, фермент, действующий на крахмал, называют амилаза, на жиры — липаза, на белки — протеиназа. До настоящего времени сохранились старые названия ферментов. Так, фермент желудочного сока называют пепсином, фермент слюны — птиалином, фермент поджелудочной железы — трипсином и т.д
Свойства ферментов Слайд №17 Все процессы, которые протекают в микробной клетке, полностью зависят от активности ферментов. Ферменты растворимы в воде, растворах солей, кислот, щелочей. Они имеют большие молекулярную массу и электрический заряд. Ферменты — это в основном белковые комплексы. По форме — это кристаллы, которые могут выпадать из растворов. Существуют две группы ферментов: однокомпонентные, или белковые, в состав котороых входит белок-носитель (апофермент); простетические, с активной группой (коферменты). В отдельности белковая и простетическая группы не обладают ферментативной активностью и только после соединения приобретают активные свойства ферментов. Специфичность действия — одно из наиболее характерных свойств ферментов. Они способны реагировать только с определенными химическими веществами или группой родственных соединений. Например, фермент лактаза расщепляет только молочный сахар, уреаза гидролизует мочевину, каталаза разрушает пероксид водорода и т. д. Каталитическая активность проявляется в очень малых количествах. Один грамм амилазы может разложить 1т крахмала, 1г химозина — свернуть 12т молока, 1г пепсина способен расщепить 50кг коагулированного белка, одна молекула каталазы при 40°С в течение секунды разрушает 550тыс. молекул пероксида водорода. Таким образом, существует большое несоответствие между количеством фермента и действием, которое он оказывает на вещество. Ферменты термолабильны. Они легко инактивируются при нагревании. При 50—600С ферменты снижают свою активность, при 80°С происходит инактивация большинства из них, а при температуре 100оС наступает полное разрушение. Это объясняется прежде всего тем, что большинство ферментов имеет белковую природу. Оптимальная температура их действия 30—50°С, для ферментов же животного происхождения — 37—40°С. Ферменты действуют при определенном рН. Реакция среды, в которой ферменты проявляют свою активность, разная. Пепсин действует в кислой среде (рН 1, 5—2, 5), трипсин — в слабощелочной (рН 7, 8—8, 7), каталаза и уреаза — в нейтральной (рН 7). Ферменты не изменяются к концу реакции, не входят в состав конечных продуктов. Они нетоксичны. Это важное свойство имеет большое значение для многих отраслей народного хозяйства. Если бы ферменты сказались ядовитыми, невозможно было бы использовать многие продукты ферментации, особенно в пищевой промышленности и медицине.
Классификация ферментов В настоящее время известно более 2000 ферментов, поэтому возникла необходимость в научно обоснованной их классификации. Согласно классификации, разработанной специальной комиссией Международного биохимического союза (1961), все ферменты объединены в шесть классов: оксидоредуктазы, трансферазы, гидролазы, лиазы, изомеразы, лигазы, или синтетазы. 6.3.1 Оксидоредуктазы — окислительно-восстановительные ферменты. Они ускоряют процессы восстановления и окисления различных веществ, играют большую роль в процессах дыхания микробов. 6.3.2 Трансферазы — ферменты переноса. Они переносят отдельные группы, радикалы и атомы как между отдельными молекулами, так и внутри их (метильные, карбоксильные и другие группы; группы, содержащие азот, фосфор, серу, альдегидные и кетонные остатки и т. д.). Представители этого класса — аминотрансферазы, фосфаттрансферазы и др. Аминотрансферазы переносят аминогруппу с одной аминокислоты на другую. Фосфаттрансферазы переносят фосфатный остаток с АТФ на глюкозу и фруктозу. 6.3.3Гидролазы — ферменты, ускоряющие реакции гидролиза, т. е. процесса расщепления сложных веществ на более простые с присоединением молекулы воды. Они имеются у многих микроорганизмов. В эту группу входят: эстеразы, расщепляющие сложные эфиры, образованные органическими кислотами и спиртами; фосфатазы, гидролизующие сложные эфиры, образованные спиртами и фосфорной кислотой; глюкозидазы, расщепляющие глюкозидные связи в углеродах и их производных; пептидазы, ускоряющие гидролиз амидов, аминокислот и других соединений. 6.3.4 Лиазы — ферменты, отщепляющие от субстратов негидролитическим путем ту или иную группу (реакция между углеродом и кислородом, азотом, серой, галоидом).Наиболее важное значение из них имеют карбоксилаза, альдегидлиаза (альдолаза) и др. 6.3.5 Изомеразы — ферменты, ускоряющие перемещение внутри молекул водорода, фосфора и двойных связей, что имеет важное значение в обмене веществ. К этой группе относятся фосфогексоизомераза, триозофосфоизомераза и др. 6.3.6 Лигазы или синтетазы — ферменты, ускоряющие синтез сложных соединений из более простых за счет распада пирофосфорных связей (в АТФ или других богатых энергией пирофосфатах). Лигазы играют большую роль в синтезе белков, нуклеиновых кислот, жирных кислот и других соединений. Ферментативную активность бактерий и грибов широко используют в промышленности для приготовления уксусной, молочной, щавелевой, лимонной кислот; молочных продуктов (сыр, ацидофилин, кумыс); в виноделии, пивоварении и др. Лекция №1.2 Тема: «Конструктивный и энергетический метаболизм микробной клетки. Общие принципы биосинтеза макромолекул у микроорганизмов. Пути метаболизма, приводящие к образованию макроэргов» Вопросы: 1. Общие принципы биосинтеза макромолекул у микроорганизмов 1.1 Взаимосвязь конструктивного и энергетического обмена 1.2 Участие отдельных химических элементов в конструктивном метаболизме 1.3 Углеродные соединения, используемые в реакциях биосинтеза 1.4 Пировиноградная кислота - ключевой метаболит катаболических и анаболических (синтетических) процессов 2. Синтез структурных компонентов микробной клетки 2.1 Компонентный состав бактериальной клетки 2.2 Биосинтез аминокислот и белков 2.3 Биосинтез нуклеотидов и нуклеиновых кислот 2.4 Биосинтез олиго- и полисахаридов 2.5 Биосинтез жиров и липидов 3. Пути метаболизма, приводящие к образованию макроэргов. 3.1 Энергетические ресурсы микробной клетки 3.2 Общая характеристика энергетических процессов Слайд№34
НАД+ + 2Н ↔ НАД Н + Н+ (окислен форма) (восстановл. форма) НАДФ+ + 2Н ↔ НАДФ Н + Н+.
Функция пиридиновых нуклеотидов в метаболизме состоит в переносе восстановительного потенциала или в его образовании. Каждая форма клеточной энергии может быть использована для осуществления химической, механической и осмотической работы, между ними существует определенное " разделение труда". Например, большинство биосинтезов обеспечивается энергией АТФ, активный транспорт — энергией протонного градиента (∆ μ н+). Из этого следует, что клетке необходимо всегда иметь определенное количество энергии в той и другой легко мобилизуемой форме. Это может быть одной из причин существования в клетке двух взаимосвязанных энергетических пулов (резервуаров), между которыми при необходимости легко может осуществляться перекачка энергии (см. рисунок 3). Емкость обоих энергетических пулов невелика. Например, величина протонного градиента (∆ μ н+) поддерживается на уровне 200-250мВ. Внутриклеточная концентрация АТФ составляет около 2мМ. Это также указывает на каталитическую роль АТФ в клетке. Подсчитано, что для удвоения клеточной массы молекула АТФ должна около 10000 раз участвовать в реакциях гидролиза и синтеза.
5.3 Энергетические затраты микробной клетки В растущей бактериальной культуре потребление энергии в первую очередь связано с процессами биосинтеза веществ, из которых состоит клетка. Количество энергии, необходимое для биосинтетических целей, в большой степени зависит от состава среды культивирования. Теоретически рассчитано, что при выращивании культуры бактерий гетеротрофов в среде с минеральными солями и глюкозой в качестве единственного источника углерода 1 моль АТФ затрачивается для синтеза 27г вещества клеток. Если же единственным источником углерода служит двуокись углерода (хемотрофы), тогда использование того же количества АТФ приведет к синтезу только 5г вещества клеток. Кроме энергетических затрат на биосинтетические процессы, связанных с ростом, определенная часть клеточной энергии всегда тратится на процессы, не связанные непосредственно с ростом. Последние получили название процессов поддержания жизнедеятельности. К специфическим функциям поддержания жизнедеятельности относятся: обновление клеточного материала, осмотическая работа, обеспечивающая поддержание концентрационных градиентов между клеткой и внешней средой, подвижность клетки и др. Энергию, расходующуюся на осуществление перечисленных функций, обозначают как энергию поддержания жизнедеятельности.
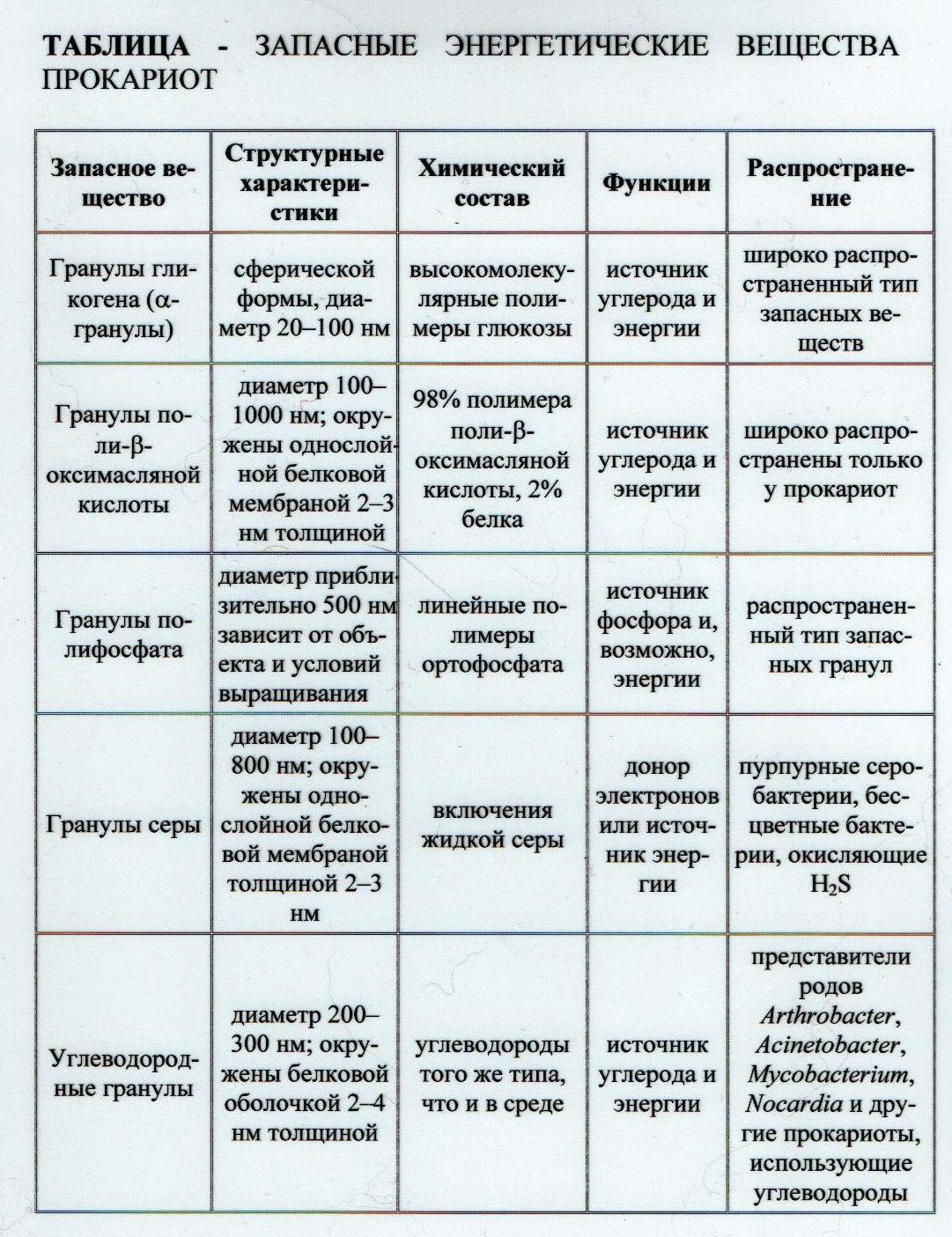
5.4 Консервирование энергии в микробной клетке Возможны такие условия, когда клетка запасает энергии больше, чем тратит. В этом случае она сталкивается с проблемой консервирования энергии. В молекулах АТФ энергия не хранится в течение длительного времени. Средняя продолжительность " жизни" молекул АТФ составляет около 1/3с. Энергия в форме протонного градиента (∆ μ н+) также не может накапливаться. Движение Н+ против градиента возможно только до достижения определенного уровня, после которого возникшая разность концентраций и электрических зарядов будет тормозить поступление ионов водорода против градиента. Таким образом, в молекулах АТФ и в виде протонного градиента (∆ μ н+) энергия находится в мобильной форме, призванной обеспечивать все идущие в настоящий момент энергозависимые процессы. Проблема консервирования энергии решена прокариотами путем синтеза восстановленных высокополимерных молекул. Эти запасные энергетические вещества представлены полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы (таблица 4). Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобкое вещество — гранулеза. Последняя -специфический запасной полисахарид анаэробных споровых бактерий группы клостридий. Названные полисахариды построены из остатков глюкозы. В неблагоприятных условиях они используются в качестве источника углерода и энергии.
Липиды накапливаются в виде гранул, резко преломляющих свет и поэтому хорошо различимых в световой микроскоп. Запасным веществом такого рода является полимер β -оксимасляной кислоты, накапливающийся в клетках многих прокариот. Другой широко распространенный тип запасных веществ многих прокариот — полифосфаты, содержащиеся в гранулах, называемых волютиновыми, или метахроматиновыми, зернами. Используются клетками как источник фосфора. Полифосфаты содержат макроэргические связи и, таким образом, являются депо энергии, хотя считается, что их роль как источника энергии незначительна. Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в клетках молекулярной серы. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды оказывается исчерпанным. Для аэробных тионовых бактерий, окисляющих сероводород (H2S), сера служит источником энергии, а для анаэробных фотосинтезирующих серобактерий она является донором электронов. Молекулы запасных энергетических веществ представлены в виде высокомолекулярных полимерных молекул, плотно упакованы в гранулах и часто окружены белковой оболочкой. В таком виде они находятся в осмотически неактивном состоянии, что очень важно для клетки, так как в противном случае сосредоточение в цитоплазме большого числа молекул осмотически активных веществ оказало бы на клетку отрицательное действие. Лекция №1.3
Тема: «Дыхание, брожение и фотосинтез микроорганизмов. Методы исследования обмена веществ у микроорганизмов»
Вопросы: Спиртовое брожение Этиловый спирт (этанол)-один из широко распространенных продуктов сбраживания сахаров микроорганизмами. Даже растения и многие грибы в анаэробных условиях накапливают этанол. Главные продуценты этанола-дрожжи, особенно штаммы Saccharomyces cerevisiae. Дрожжи, как и большинство других грибов, осуществляют аэробное дыхание, но без доступа воздуха они сбраживают углеводы до этанола и углекислого газа. У ряда анаэробных и факультативно-анаэробных бактерий этиловый спирт тоже является главным или побочным продуктом сбраживания гексоз или пентоз. Еще Гей-Люссак (в 1815 г.) дал уравнение, описывающее превращение глюкозы в этанол в той форме, как это принято и теперь: СбН1206 → 2С02 + 2С2Н5ОН
Превращение пирувата в этанол происходит в два этапа. Сначала пируват декарбоксилируется пируватдекарбоксилазой(а) при участии тиаминпирофосфата до ацетальдегида, а затем ацетальдегид восстанавливается алкогольдегидрогеназой (б) в этанол при участии NAD H2. Слайд №37. Процесс спиртового брожения суммарно можно выразить следующим уравнением: СбН12Об + 2ФН + 2АДФ → 2СН3-СН2ОН + 2С02 + 2АТФ +2Н20. Сырьем для производства спирта с использованием дрожжей служат углеводы растительного происхождения (картофель, злаки), отходы пищевой (мелассы) и целлюлозно-бумажной (щелока) промышленности, различные сельскохозяйственные отходы, а также гидролизаты древесины. Сбраживание дрожжами виноградного сока лежит в основе виноделия; сбраживание пивного сусла, приготовленного из проросших зерен ячменя, специальными пивными дрожжами — в основе пивоварения. 6. Спектральный состав солнечного света Электронов Хлорофилл играет двоякую роль в превращении энергии при фотосинтезе: он служит светопоглощающим пигментом и участвует в первичной фотохимической реакции. В то же время каротиноиды и фикобилипротеиды функционируют только как светопоглощающие пигменты, передавая энергию поглощенного ими света в реакционный центр (т.е. хлорофиллу). Энергия фотонов, поглощенных фотосинтезирующей пигментной системой, переносится в реакционные центры, где находятся в особом состоянии молекулы хлорофилла, тесно связанные с участвующими в фотосинтезе компонентами цепи переноса электронов. Слайд №51. Процесс фотохимического превращения энергии начинается в тот момент, когда молекула хлорофилла в реакционном центре поглощает энергию света. Молекула хлорофилла окисляется, освобождая электрон, который акцептируется ферредоксином: 1. Хлорофилл + Энергия света → Хлорофилл+ + е-; 2. Ферредоксин + е- → Восстановленный ферредоксин. Обратное окисление восстановленного ферредоксина дает примерно столько же энергии, сколько окисление молекулярного водорода. Эта энергия передается другим переносчикам электронов фотосинтетической цепи транспорта электронов и используется таким образом для образования АТФ. Слайд №52. При аэробном и анаэробном типах дыхания и при фотосинтезе АТФ образуется в результате прохождения электронов от первичного донора электронов через цепь переноса до конечного акцептора. Доноры и акцепторы электронов, участвующие в различных реакциях образования АТФ за счет транспорта электронов, перечислены в таблице 3.
При дыхании первичный донор электронов — окисляемый субстрат, а конечный акцептор электронов — неорганическое соединение: либо 02 при аэробном дыхании, либо N0-3, S02- и С02- при анаэробном дыхании. В процессе фотосинтеза молекулы хлорофилла реакционных центров типа I служат и донорами, и акцепторами электронов для циклического фотофосфорилирования. Вместе с тем, при нециклическом фосфорилировании в качестве первичного донора электронов используется вода или другое химическое соединение, а в качестве конечного акцептора электронов — окисленный хлорофилл (присоединение электронов к НАДФ- в реакционных центрах типа I). Слайд №53. Хотя сложность и набор компонентов цепей переноса электронов варьируют, все подобные цепи обладают некоторыми общими свойствами: во-первых, компоненты цепи — это переносчики, способные легко вступать в обратимые реакции окисления и восстановления; во-вторых, АТФ образуется в результате прохождения электронов по цепи. Цепь переноса электронов может также функционировать и в обратном направлении, когда образование восстановительного потенциала путем непосредственного переноса электронов от неорганического субстрата на НАД+ (никотинадениндинуклеотид-пиримидиновое основание) термодинамически невыгодно. Лекция 2.1 Типы взаимоотношений Нейтрализм – популяции не влияют друг на друга, если разделены в пространстве или во времени. .Положительные взаимодействия: Комменсализм – популяция комменсала получает пользу от объединения с популяцией хозяина, для которого это объединение безразлично. В некоторых случаях это метабиоз Метабиоз. В природе это явление распространено очень широко. При метабиозе продукты жизнедеятельности одного микроорганизма потребляются другими микроорганизмами в качестве питательного материала. Это почти всегда имеет место при последовательном использовании какого-либо одного сложного субстрата. Так, например, при использовании белковых веществ последовательно могут принимать участие в этом процессе аммонификаторы, нитрификаторы и денитрификаторы. Мутуализм (взаимовыгодный) – облигатное взаимодействие полезное для обеих популяций. В микробиологии этот тип взаимодействий обычно называют симбиозом. Это симбиоз гриба и водоросли в лишайниках, симбиоз клубеньковых бактерий с бобовыми, симбиоз папоротника азолла с цианобактериями. Кефирные зерна (дрожжи и молочнокислые бактерии), чайный гриб (уксуснокислые бактерии и дрожжи) Мутуализм иногда обозначают как синтрофные взаимодействия. Синтрофными отношениями называют пищевые связи в том случае, когда субстрат потребляется только смешанными популяциями, не способными к его использованию в чистых культурах. Наиболее широко распространенным механизмом, лежащим в основе синтрофного взаимодействия, является обмен факторами роста. Также часто встречаются синтрофные ассоциации, основанные на обмене субстратами роста или удалении токсических продуктов обмена. Mucor rammanianus и дрожжи Rhodotorula rubra нуждаются в тиамине (В1). Гриб при совместном росте выделяет пиримидиновый компонент, дрожжи – тиазоловый компонент молекулы тиамина, при этом удовлетворяются потребности обоих партнёров. Протокооперация – взаимодействие друг с другом полезно для обеих популяций, но не является облигатным. Взаимодействие целлюлозоразрушающих грибов, дающих бактериям глюкозу и бактерий, фиксирующих для грибов азот. Метаноокисляющие и метонолокисляющие бактерии часто встречаются вместе. Вокруг гетероцист цианобактерий накапливается много хемотрофных бактерий, в т.ч. водородокисляющих. В природе кооперация между организмами различных метаболических типов скорее правило, чем исключение. При использовании в биотехнология микроорганизмоа по отдельности кооперативную активность часто полностью упускают из вида. Отрицательные взаимодействия: В естественных условиях развития микробов довольно часто могут наблюдаться явления не только взаимно благоприятные, но и такие, при которых один вид микроорганизмов тем или иным способом угнетает или полностью подавляет рост и развитие других видов. Такая форма взаимоотношений называется антагонизмом. Это явление широко распространено у грибов, бактерий и актиномицетов. Конкуренция (непосредственное взаимодействие)– прямое взаимное подавление обоих видов. Микробы антагонисты образуют антибиотики, действующие друг против друга. Конкуренция из-за ресурсов – опосредованное подавление, возникающее, когда появляется недостаток в каком-либо факторе, используемом обоими видами. Это наиболее широко распространённый вид конкуренции: за органическое вещество, за источники азота, фосфора, калия, железа, витамины, воду, свет. Обычно побеждает тот организм, который быстрее растёт или который может усваивать вещество из рассеянного состояния. Правило Гаузе: два вида, занимающие одну и туже нишу, не могут существовать вместе. Аменсализм (несотрапезники) – одна популяция подавляет другую, но сама не испытывает отрицательного влияния (микроб-продуцент антибиотика и чувствительный микроорганизм). Паразитизм – паразит существует за счёт вещества (клетки) хозяина, иногда убивает его, иногда приносит вред, не убивая. В экологии как паразиты обычно рассматриваются патогенные для растений и животных микроорганизмы, фаги, Bdellovibrio. Бактерии-паразиты в своём эволюционном развитии утратили способность синтезировать многие вещества, они получают их в готовом виде за счёт своего хозяина. Хозяин же взамен от такого сожительства никакой пользы не получает. Бактерии, как правило, внеклеточные паразиты, а риккетсии, хламидии и вирусы – внутриклеточные. Риккетсии - вызывают у человека ряд опасных инфекционных заболеваний - сыпной тиф, лихорадку Скалистых гор. Хламидии являются также возбудителями ряда инфекционных заболеваний (орнитоз, трахома, и др.). Хищничество – уничтожение хищником жертвы, тоже одна из форм антагонизма. Процесс хищничества состоит в том, что некоторые микробы поглощают клетки других видов микроорганизмов и используют их в качестве питательного материала. Популяция хищника меньше популяции жертвы. Простейшие питаются бактериями и грибами, хищные грибы поедают нематод. К числу микроорганизмов-хищников относятся главным образом, миксоформы (миксобактерии, миксомицеты) Следует иметь ввиду, что рассмотренные типы взаимодействий, существующие в мире микроорганизмов, не могут исчерпать всё то разнообразие связей, которое имеет место среди микроорганизмов в природе. Два общих экологических принципа: в ходе эволюции и развития экосистемы существует тенденция к уменьшению роли отрицательных взаимодействий и увеличению роли положительных, способствующих выживанию взаимодействующих видов. В новых ассоциациях вероятность возникновения сильных отрицательных взаимодействий больше, чем в старых ассоциациях. Гипотеза симбиогенеза (К.С.Мережковский, 1910) и её значение. Симбиозы стали мощным фактором эволюции более 1-1.5 млрд.лет назад, начиная с образования эукариотической клетки. Это принципиальное усложнение организации клетки скорее всего является результатом симбиотического объединения древних прокариот. Сейчас она получила подтверждение на молекулярном уровне благодаря работам Л.Маргулис., хотя пока не признана теорией. Митохондрии произошли от аэробных органотрофных грамотрицательных бактерий, а пластиды от оксигенных цианобактерий. Высказано мнение, что прокариотические клетки, давшие начало пластидам, митохондриям и возможно ядру, поглощались путём фагоцитоза клеткой-предшественницей эукариотической. В результате такого комплексного эндосимбиоза произошёл глобальный ароморфоз (принципиальное усложнение организации), положивший начало развитию многоклеточности и образованию нового надцарства – эукариот. Многоклеточные организмы также вступали в симбиозы с микроорганизмами. Их совместная жизнь определяет и их совместную эволюцию (коэволюцию). Таким образом, концепция симбиоза имеет большое значение для понимания эволюции и филогенитических отношений между прокариотами и эукариотами. Для объяснения функционирования и эволюции микробных сообществ и биосферы в целом, Г.А.Заварзиным недавно был предложен, в качестве основополагающего, принцип аддитивности, предполагающий кооперацию между организмами, а не конкуренцию, как считали ранее, движущей силой процесса эволюции микробных сообществ. " Кооперация лежит в основе трофической пирамиды сообществ, а конкуренция служит механизмом тонкой регулировки в системе" (Заварзин, 1995). В практическом отношении для нас представляют более значительный интерес антагонистические взаимодействия микроорганизмов, изучение которых на протяжении последнего века послужили основой современной химиотерапии. Впервые обобщение отдельных фактов микробного антагонизма осуществил И.И.Мечников. Он же наметил пути его использования на практике. Его учение о преждевременном старении в связи с постоянной интоксикацией организма гнилостными бактериями кишечника и использование молочнокислых палочек простокваши для вытеснения этих гнилостных бактерий заложили научные основы современного учения об антагонизме микроорганизмов. Лекция 2.2 Излучения. Свет – фактор, необходимый для роста фотосинтезирующих микроорганизмов, например, цианобактерий, зелёных и пурпурных бактерий, которые имеют пигменты (в частности, каратиноидные), обеспечивающие поглощение энергии солнечного луча и превращать её в химическую. Для большинства других бактерий прямые солнечные лучи бесполезны или даже вредны. Губительное действие обусловлено негативным влиянием коротковолнового спектра – УФ. Все патогенные микробы являются фотофобами. Сапротрофные микробы и споры более устойчивы к действию света, особенно пигментированные. Установлено, что УФ лучи вызывают коагуляции белков, разложение некоторых сложных органических веществ, инактивируют ферменты. Поэтому Уф лампы используют для стерилизации воздуха, создании стерильности при массовых посевах культур в промышленности и тд. УФ-излучение с длиной волны 400-320 нм вызывает мутагенный и летальный эффект. Гибель клетки связана с повреждением ДНК и мембран. |
Последнее изменение этой страницы: 2017-04-12; Просмотров: 816; Нарушение авторского права страницы