|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
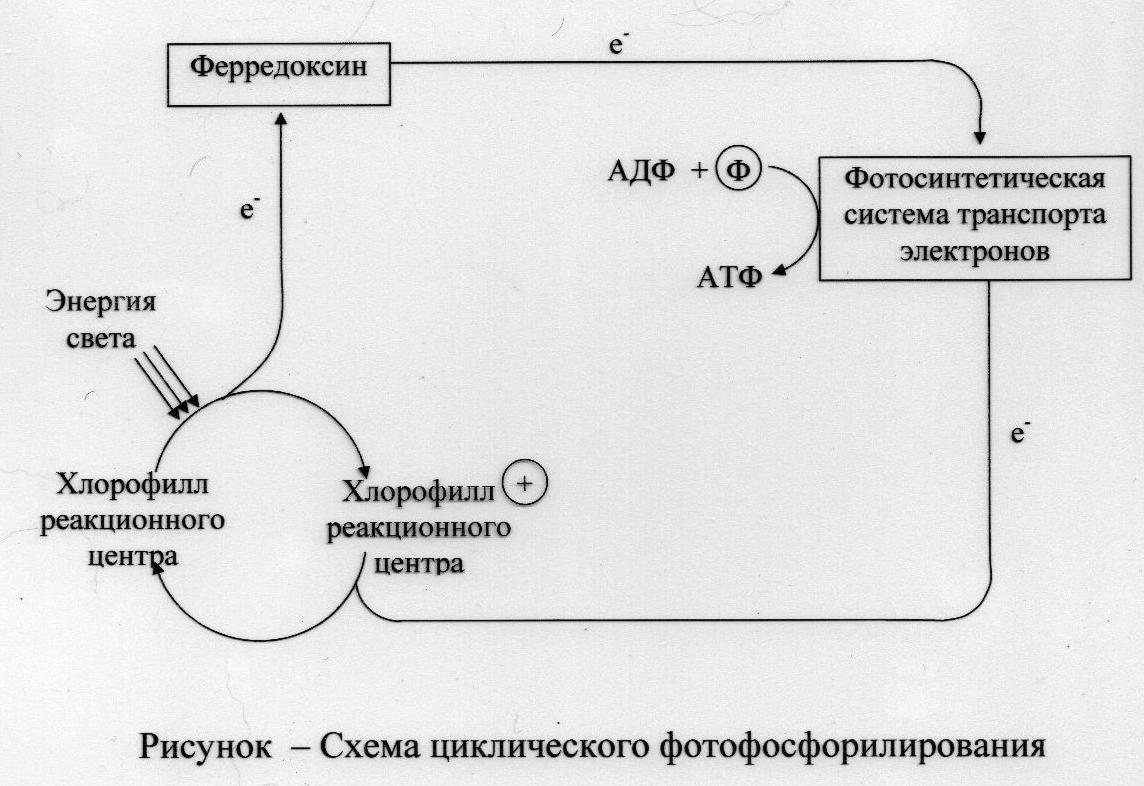
Механизмы фотофосфорилирования
Слайд №54. У всех фотосинтезирующих организмов энергия возбужденных электронов хлорофилла может быть использована для образования АТФ в процессах трёх типов: -циклическое фотофосфорилирование; - нециклическое фотофосфорилирование; -сопряжённое фотофосфорилирование (комбинация циклического и нециклического процессов.) Схема циклическогофотофосфорилирования приведена на рисунке.
причем последний компонент этой цепи восстанавливает хлорофилл; таким образом, электроны проходят по замкнутому пути, а их поток включается и выключается за счет поглощения энергии света. Часть энергии света улавливается цепью переноса благодаря синтезу АТФ. Слайд №56 Схема нециклическогофотофосфорилирования приведена на рисунке. Этот процесс представляет собой другой путь использования электронов, выброшенных хлорофиллом реакционного центра под действием света, — восстановление пиридиновых нуклеотидов. При этом поток электронов становится открытым, незамкнутым, так как катион хлорофилла+ должен быть восстановлен электронами подходящего донора через систему переноса электронов фотосинтезирующего аппарата. При таком переносе электронов также образуется АТФ. Вариант нециклического фотофосфорилирования при кислородном фотосинтезе осуществляют фотохимические реакционные центры, которые относят к типу II.
Данный вариант фотофосфорилирования используют растения, водоросли, цианобактерий и прохлорофиты в том случае, когда необходимо эффективно утилизировать термодинамически невыгодный донор электронов - воду. Фотосинтезирующий аппарат этих организмов содержит фотохимические реакционные центры двух типов (типа I –циклический и типа II-нециклический). Их можно отличить по разным ответам при воздействии на них света с определенной длиной волны и ингибиторов фотосинтеза. Реакционные центры одного типа (типа I) осуществляют и циклическое, и нециклическое фотофосфорилирование. Однако, поглощения света только этими реакционными центрами недостаточно для одновременного восстановления НАД Ф и окисления воды. Нециклическое фотофосфорилирование кислородного типа требует одновременного поглощения света реакционным центром другого типа (типа II), в котором и происходит фотохимическое окисление воды. Электроны, освободившиеся в ходе этого окисления, проходят через фотосинтетическую цепь переноса электронов и восстанавливают окисленный хлорофилл, присоединяясь к НАДФ в реакционных центрах типа I. Таким образом, фотосистема II-типа была достроена к фотосистеме I-типа для того, чтобы стало возможным использование воды в качестве донора электронов. Побочный продукт этого процесса — молекулярный кислород. Фотосинтез, осуществляемый при координированном функционировании двух фотосистем и сопровождающийся выделением кислорода (02 ) из воды, стал одним из основных типов энергетического метаболизма у высших форм жизни и занимает доминирующее положение в энергетической системе живого мира. Таким образом неспособность бактерий образовывать кислород при фотосинтезе обусловлена отсутствием реакционных центров типа II в аппарате бескислородного фотосинтеза. Фотохимические реакционные центры бескислородного фотосинтеза относятся исключительно к типу I и осуществляют циклическое фотофосфорилирование. С точки зрения фотохимии бескислородный фотосинтез — более простой процесс, чем кислородный фотосинтез.
8. Особенности конструктивного метаболизма у фотосинтезирующих бактерий (биосинтетические процессы)
Мы рассмотрели, каким образом энергия света, поглощенная пигментами фотосинтезирующего аппарата, используется для энергетических реакций- синтеза АТФ и синтеза восстановленных пиридиновых нуклеотидов. Эти реакции, сопровождающиеся превращением энергии света в химическую энергию, свойственны исключительно фотосинтезу. Вместе с тем конкретные механизмы использования этой химической энергии для биосинтетических реакций не только не являются какой-то особенностью фотосинтеза, но, строго говоря, не являются и частью процесса фотосинтеза. Последующие биосинтетические процессы включают так называемые темновые реакции, катализируемые ферментами в отсутствие света.
В этом цикле углекислота фиксируется в четырех ферментативных реакциях, две из которых идут при участии фотохимически восстановленного ферредоксина, а одна — таким же путем образованного НАД-Н2 (никатинадениндинуклеотид-Н2). В результате одного оборота цикла из 4 молекул углекислого газа, 10 молекул водорода с использованием энергии (3 молекулы АТФ) синтезируется молекула щавелевоуксусной кислоты — Все реакции, в которых происходит фиксация углекислого газа в цикле, функционируют как механизмы хемогетеротрофной фиксации углекислого газа или аналогичные им. Слайд №59 Восстановительный пентозофосфатныйцикл, или цикл Кальвина, являющийся основным путем фиксации углекислого газа у всех высших фотосинтезирующих организмов, функционирует в группе пурпурных бактерий, у цианобактерий и прохлорофит (для студентов - см. рис.77 у Гусева). Последовательность ферментативных реакций, приводящих к фиксации углекислоты и образованию из нее молекулы гексозы, была расшифрована М. Кальвином (М. Calvin) с сотрудниками в 50-х гг. В этом цикле новое - это химическая природа акцептора. Акцепторами двуокиси углерода во всех до сих пор описанных реакциях были органические кислоты в обычной или активированной форме. В этом цикле впервые акцептором двуокиси углерода выступает вещество углеводной природы — активированная молекула пентозы. Для восстановительного пентозофосфатного цикла уникальными являются два фермента, не участвующие в других метаболических путях: фосфорибулокиназа и рибулозо-дифосфат-карбоксилаза. Последовательные ферментативные превращения ведут к образованию молекулы глюкозы. Эти превращения включают реакции, известные в гликолитическом пути, но они идут теперь в обратном направлении. Такова биосинтетическая часть цикла, ведущая к фиксации углекислого газа и образованию из нее молекулы гексозы. Восстановительный пентозофосфатный цикл является основным механизмом автотрофной ассимиляции углекислоты. Последняя у большинства фотосинтезирующих бактерий восстанавливается с помощью фотохимически образованной " ассимиляционной силы" — АТФ и восстановителя. Однако и АТФ, и восстановитель - пиридиновые основания ( НАДФ-Н2 никатинадениндинуклеотидфосфат-Н2 или НАД-Н2 никатинадениндинуклеотид-Н2) образуются в разных метаболических путях. Поэтому нельзя рассматривать восстановительный пентозофосфатный цикл ассимиляции двуокиси углерода строго привязанным только к фотосинтезу. У большой группы хемоавтотрофных эубактерий этот путь фиксации двуокиси углерода сочетается с темновыми окислительными процессами получения энергии. Важно отметить только, что это основной путь ассимиляции углекислоты, если последняя служит единственным или главным источником углерода. |
Последнее изменение этой страницы: 2017-04-12; Просмотров: 1821; Нарушение авторского права страницы