|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Структура и функции гена. Особенности структурной организации генов у про- и эукариот.
Теория гена Генетический код – система записи генетической информации в молекуле ДНК (РНК) в виде определенной последовательности нуклеотидов. Этот код служит ключом для перевода последовательности нуклеотидов в и-РНК в последовательность аминокислот полипептидной цепи при ее синтезе. Свойства генетического кода: 1. Триплетность – каждая аминокислота кодируется последовательностью из трех нуклеотидов (триплетом или кодоном) 2. Вырожденность – большинство аминокислот шифруется более чем одним кодоном. В ДНК или РНК имеется 4 различных нуклеотида, которые теоретически могут образовывать 64 разных триплета для кодирования 20 аминокислот, входящих в состав белков. Этим и объясняется вырожденность генетического кода. 3. Неперекрываемость – один и тот же нуклеотид не может входить одновременно в состав двух соседних триплетов. 4. Специфичность (однозначность) – каждый триплет кодирует только одну аминокислоту. 5.Код не имеет знаков препинания. Считывание информации с и-РНК при синтезе белка всегда идет в направлении 5, -- 3, в соответствии с последовательностью кодонов иРНК. Если произойдет выпадение одного нуклеотида, то при считывание его место займет ближайший нуклеотид из соседнего кода, из-за чего изменится аминокислотный состав в молекуле белка. 6.Код универсален для всех живых организмов и вирусов: одинаковые триплеты кодируют одинаковые аминокислоты. Универсальность генетического кода свидетельствует о единстве происхождения всех живых организмов. Среди триплетов универсального генетического кода три кодона не кодируют аминокислоты и определяют момент окончания синтеза данной полипептидной молекулы. Это так называемые «nonsens» кодоны (стоп-кодоны или терминаторы). К ним относятся: в ДНК – АТТ, АЦТ, АТЦ; в РНК – УАА, УГА, УАГ. Классификация генов Гены, как единицы функции наследственного материала, разделяются на структурные, регуляторные и гены-модуляторы. Структурные гены содержат информацию о структуре белка (полипептидов) и РНК (рРНК и тРНК), при этом генетическая информация реализуется в процессе транскрипции и трансляции или только транскрипции. Жизнедеятельность клеток обеспечивается небольшим набором функционирующих генов, среди них различают гены общеклеточных функций и гены специализированных функций. Регуляторные гены (ген-регулятор лактозного оперона), координируют активность структурных генов на уровне клетки, а также дерепрессию и репрессию генов на уровне организма. Гены-модуляторы усиливают или ослабляют действие структурных генов, изменяя их функциональную активность.
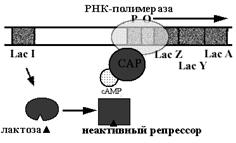
Структурная организация генов у прокариот Гены организованы в виде независимых генов, транскрипционных единиц и оперонов. Независимые гены состоят из непрерывной последовательности кодонов, они постоянно экспрессированы и не регулируются на уровне транскрипции (ген-регулятор лактозного оперона). Транскрипционные единицы - группы разных генов, которые связаны функционально и транскрибируются одновременно, что обеспечивает в последующем одинаковое количество синтезируемых продуктов. Обычно это гены белков или нуклеиновых кислот (у кишечной палочки в составе одного из транскриптонов находятся два гена т-РНК, три гена р-РНК). Оперон - это группа структурных генов, следующих друг за другом, находящихся под контролем оператора – определённого участка ДНК. Оперонная организация структурных генов бактерий Он состоит из: 1. Промотор - участок ДНК, с которым происходит связывание РНК-полимеразы и который определяет точку начала транскрипции. Типичный пример - TTGACA - 17 bp - TATAAT. В зависимости от типа промотора (а точнее, от типа распознающей его сигма - субъединицы РНК полимеразы) конкретная последовательность промотора варьирует. 2. Оператор - участок связывания регуляторного белка. Размер - около 20 bp. Располагается в непосредственной близости к промотору или же перекрывается с ним. В случае негативных регуляторов, или репрессоров, оператор, как правило, располагается непосредственно за промотором, или перекрывается с ним. В случае позитивных регуляторов (активаторов) оператор обычно располагается перед промотором. Связывание регуляторного белка с оператором меняет частоту инициации транскрипции. У многих (возможно, у большинства) оперонов имеется не один, а несколько сайтов связывания с регуляторными белками, которые не обязательно располагаются рядом и могут вообще находиться по разные стороны от промотора. В этих случаях термин " оператор" в классическом смысле становится неудобным, в связи с чем сейчас чаще просто говорят о сайтах связывания регуляторов. 3. Структурные гены. Кодируют белки, непосредственно производящие фенотипический эффект. Именно для контроля их экспрессии, собственно, и существуют оперонные структуры вместе со своими регуляторами. 4. Терминатор транскрипции. Здесь заканчивается синтез мРНК. Не входит в оперон, но является необходимой частью регуляторной системы ген-регулятор, кодирующий регуляторный белок, связывающийся с оператором. Ген-регулятор может находиться рядом с контролируемым им опероном, но часто располагается совсем в другом участке хромосомы. Почти всегда у гена-регулятора свой промотор и терминатор. В регуляции участвуют, как правило, и низкомолекулярные вещества-эффекторы, являющиеся либо индукторами, либо корепрессорами структурных генов. В зависимости от влияния на их работу низкомолекулярных молекул-эффекторов различают индуцибельные и репрессибельные (-руемые) опероны. В зависимости от эффекта связываниярегуляторного белка с оператором опероны могут иметь негативный или позитивный контроль. Т. о., можно выделить четыре типа оперонов. Lac оперон Относится к индуцибельным оперонам. Lac-оперон состоит из кодирующей области, представленной тремя структурными генами, ответственными за синтез ферментов: β -галактозидазы (lacZ - гидролиз до глюкозы и галактозы), β -галактозидпермеазы (lacY – мембранный транспортный белок, который переносит лактозу внутрь клетки) и β -галактозидтрансацетилазы (lacA – фермент, переносящий ацетильную группу от ацетил-КoA на бета-галактозиды); а также из промоторно-операторной области. Для катаболизма лактозы необходимы только продукты генов lacZ и lacY, роль продукта гена lacA не ясна. Возможно, что реакция ацетилирования дает бактериям преимущество при росте в присутствии определенных неметаболизируемых аналогов бета-галактозидов, поскольку эта модификация ведет к их детоксикации и выведению из клетки. Ген-регулятор лактозного оперона – lacI, – кодирует белок-репрессор. В активной форме это тетрамер. В основе индукции синтеза ферментов лактозного оперона лежит механизм негативной, или отрицательной, регуляции. В отсутствие лактозы молекула репрессора, активная в свободном состоянии, связывается с оператором, закрывая при этом промотор, что препятствует связыванию с ним РНК-полимеразы и началу транскрипции структурных генов. При наличии в среде внешнего индуктора лактоза транспортируется с помощью β -галактозидпермеазы внутрь клетки и с помощью фермента β -галактозидазы превращается в аллолактозу, которая действует как внутренний индуктор. Аллолактоза связывается с репрессором, который при этом претерпевает конформационное изменение, уменьшающее его сродство к ДНК оператора, и в результате репрессор отсоединяется от lac-оператора. Начинается транскрипция структурных генов, приводящая к синтезу ферментов катаболизма лактозы. При удалении из клетки индуктора репрессор снова переходит в активное свободное состояние, связывается с оператором, что приводит к прекращению синтеза соответствующих ферментов. Лактозный оперон подвержен также регуляции другого типа – позитивной, или положительной. Дело в том, что РНК-полимераза может связаться с промотором лишь тогда, когда к нему присоединен регуляторный белок БАК (белок, активирующий катаболизм), или САР (catabolite activator protein). Однако БАК может связаться с промотором только в том случае, если в клетке в достаточно высокой концентрации присутствует цАМФ, т. е. БАК связывается с промотором только в комплексе с цАМФ. Если в клетке цАМФ отсутствует, то БАК не способен взаимодействовать с промотором. Это было установлено с использованием феномена диауксического роста (или диауксии) – при наличии в среде глюкозы и лактозы клетки бактерий вначале используют глюкозу, а затем после ее полного израсходования начинают катаболизировать лактозу. Оказалось, что глюкоза репрессирует синтез β -галактозидазы. При наличии в среде глюкозы в клетке резко снижается количество цАМФ. Это явление называют катаболитной репрессией. Оно наблюдается и в тех случаях, когда вместо лактозы используется какой-то другой углевод. Катаболитная репрессия глюкозой может быть снята, если в среду добавить цАМФ. Образуется комплекс цАМФ с БАК, и РНК-полимераза присоединяется к промотору, а далее идет синтез ферментов катаболизма лактозы, даже в присутствии глюкозы.
Кроме индуцибельных оперонов, управляющих катаболизмом углеводов, у бактерий имеются и репрессибельные опероны. Это опероны, ответственные за синтез аминокислот аргинина, гистидина и триптофана. Максимальная транскрипция структурных генов этих оперонов достигается только при отсутствии в клетке конечных продуктов или эффекторов этих биосинтетических путей. Такие эффекторы, которыми являются конечные продукты, называют корепрессорами, а соответствующие регуляторные белки – апорепрессорами. Синтез ферментов репрессибельного оперона включается посредством дерепрессии структурных генов. Триптофановый оперон Оперон состоит их пяти структурных генов, ответственных за синтез пяти ферментов, участвующих в превращении хоризмовой кислоты в триптофан, а также из промоторно-операторной области. Ген-регулятор (trрR) расположен на хромосоме на некотором расстоянии от оперона. Он ответственен за синтез регуляторного белка – апорепрессора, который неактивен в свободном состоянии, не может связываться с оператором и неспособен, таким образом, препятствовать началу транскрипции. Когда конечный продукт метаболического пути – триптофан – накапливается выше определенного уровня, взаимодействует с апорепрессором и активирует его. Активированный апорепрессор (апорепрессор + корепрессор) присоединяется к оператору и подавляет транскрипцию структурных генов триптофанового оперона. Синтез триптофана прекращается. Установлено, что отсутствие активированного репрессора вызывает примерно 70-кратное увеличение актов инициации транскрипции. Но даже в условиях репрессии структурные гены сохраняют низкий уровень экспрессии. Для того чтобы понизить уровень транскрипции в присутствии триптофана в еще большей степени, в клетках бактерий E. coli имеется дополнительный механизм регуляции транскрипции, который называется аттенуацией, в осуществлении его принимает участие продукт гена trpL. В условиях избытка триптофана только одна из десяти молекул РНК-полимеразы, начавшая транскрибирование с промотора, взаимодействует со структурными генами и продолжает транскрипцию. Таким образом, действие аттенуатора проявляется в терминации транскрипции, а сам процесс аттенуации классифицируется как регулируемая терминация транскрипции. В отличие от репрессии, аттенуация зависит не от самой аминокислоты, а от образования триптофанил-тРНК, т. е. активированной аминокислоты, присоединенной к транспортной РНК. При уменьшении внутриклеточной концентрации триптофана сначала осуществляется дерепрессия. Это значит, что образуется возможность связывания молекул РНК-полимеразы с промотором Trp-оперона. При более глубоком голодании снижается уровень триптофанил-тРНК и возникают условия для преодоления аттенуатора. Регуляция триптофанового оперона: негативная (белок-репрессором) и аттенуация (регуляция транскрипции с помощью трансляции). Таким образом, и в случае индукции путем негативной регуляции (Lac-оперон), и в случае репрессии синтеза ферментов (Тrp-оперон) взаимодействие репрессора с оператором приводит к подавлению транскрипции соответствующих структурных генов. Различие заключается в том, что при индукции путем негативной регуляции эффектор (индуктор), взаимодействуя с репрессором, понижает сродство репрессора к оператору, а в случае репрессии эффектор (корепрессор) повышает это сродство.
|
Последнее изменение этой страницы: 2017-04-13; Просмотров: 2510; Нарушение авторского права страницы