|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Цепочечная нервная система аннелид
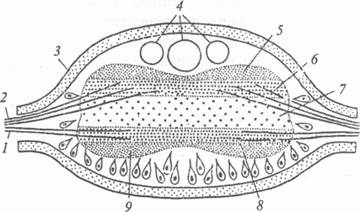
Для типа кольчатых червей характерны вторичная полость тела (целом), метамерное строение мышечного аппарата, а также сегментация кровеносной, выделительной и нервной систем. В типичном случае нервная система кольчецов состоит из окологлоточного нервного кольца (оно включает парные надглоточный, подглоточный ганглии и окологлоточные коннективы) и брюшной нервной цепочки. Брюшная цепочка формируется при слиянии парных брюшных стволов в результате укорочения поперечных комиссур. У большинства аннелид в каждом сегменте тела находится по одной паре ганглиев, соединенных продольными коннективами с парами ганглиев в соседних сегментах. Нервные элементы у кольчецов концентрируются вследствие сближения парных ганглиев одного сегмента, укорочения продольных коннектив, ведущего к слиянию последовательных ганглиев различных сегментов. Так, например, подглоточный ганглий дождевого червя образуется при слиянии двух парных ганглиев брюшной нервной цепочки; у пиявки семь задних ганглиев сливаются в ганглиозную массу задней присоски. Централизация нервного аппарата у аннелид не исключает существования субэпителиального нервного сплетения, где локализовано много рецепторных нейронов, аксоны которых идут в сегментарные ганглии или теряются на периферии среди мышечных волокон. Структура сегментарных ганглиев малощетинковых червей и пиявок детально описана в монографических исследованиях ГА. Невмываки (1966) и Ю.П. Лагутенко (1931). Здесь мы остановимся только на основных принципах организации этих анатомических образований. Брюшная нервная цепочка дождевого червя снаружи одета соединительнотканной оболочкой, под которой проходит слой продольных мышечных волокон. От каждого ганглия отходят три пары боковых периферических нервов: передняя и задняя пары - преимущественно двигательные, средняя - чувствительная. Двигательные нервы соединяются между собой на спинной стороне, образуя кольцевые нервы, иннервирующие мускулатуру каждого сегмента. Парные ганглии и периферические нервы формируют сегментарный невросомит. У многощетинковых кольчецов в состав невросомита могут входить еще и параподиальные ганглии, расположенные у основания параподий и связанные с чувствительными нервами средней пары. Приходящие с периферии чувствительные: волокна вступают в ганглий через вентральные корешки трех пар боковых нервов, и их разветвления всегда располагаются в вентральной части ганглия. Двигательные волокна выходят через дорсальные корешки, и их коллатерали сосредоточены в дорсальной части ганглия. Среднюю часть ганглия занимает мозговое вещество - нейропиль, где переплетаются клеточные отростки и находятся глиальные элементы. В нейропиле сконцентрирована основная масса синаптических контактов. Таким образом, ганглий на поперечном срезе имеет четко выраженную дорсовентральную слоистость (рис. 3). Наиболее дорсально расположена область дорсальных ассоциативных волокон. Кроме тонких ассоциативных волокон здесь проходят гигантские аксоны, которые тянутся практически по всей длине брюшной нервной цепочки. У земляного червя, как почти у всех олигохет, имеются три гигантских нервных волокна (два латеральных и одно медиальное). Максимальный диаметр медиального волокна 75 мкм, латеральных 50 мкм. Скорость проведения возбуждения коррелирует с диаметром гигантских аксонов и составляет 15-45 м/с для медиалъного и 7-15 м/с для латеральных волокон. Волна возбуждения сопряжена с потенциалом действия, средняя амплитуда которого достигает 100-120 мВ.
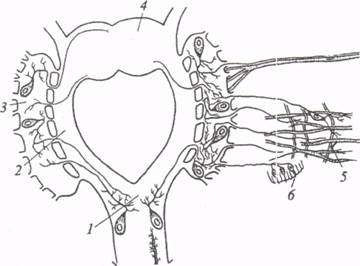
Рис. 5. Поперечный разрез брюшного ганглия дождевого червя (по: Г.А. Невмывака, 1966): 1 - чувствительные (вентральные) корешки, 2 - двигательные (дорсальные) корешки, 3 - оболочка ганглия, 4 - гигантские волокна, 5 - дорсальная ассоциативная область, 6 - двигательная область, 7 - нейропиль, 8 - чувствительная область, 9 - вентральная ассоциативная область Гигантские аксоны разделяются поперечными перегородками септами) на сегменты. Каждая септа состоит из двух трехслойных мембран, разделенных гомогенным пространством (10 нм). Формируемый таким образом септальный синапс не поляризован и проводит возбуждение практически без синаптической задержки. Септальные синапсы электрические. Однако по обеим сторонам септальных мембран обнаружены синаптические везикулы, поэтому некоторые авторы предполагают включение химического звена при проведении через септы. Между двумя латеральными гигантскими аксонами дождевого червя также обнаружена электротоническая связь через комиссуральные ветви. Здесь находится неполяризованный электрический синапс, который проводит возбуждение с задержкой порядка 0, 5 мс. В пределах каждого сегментарного ганглия гигантские аксоны имеют афферентные и эфферентные связи, превращаясь, таким образом, в сквозные проводящие пути, которые компенсируют расчлененность цепочечной нервной системы. Относительно высокая скорость проведения обеспечивает участие гигантских аксонов в реализации быстрых рефлекторных реакций. Вентральнее под зоной дорсальных ассоциативных волокон в сегментарном ганглии расположен моторный нейропиль, который формируется из отростков двигательных нейронов. Тела моторных клеток обычно находятся на периферии ганглия, с его дорсальной и вентральной сторон. Форма клеточных тел грушевидная, причем большинство нейронов униполярные с единственным интегративным отростком, который на некотором расстоянии от сомы формирует боковые дендритные ответвления и аксон, идущий в сегментарные нервы. Часть клеток двигательной зоны имеет биполярную и реже мультиполярную форму. Прежде чем попасть в боковые нервы ганглия, отростки клеток моторного нейропиля проделывают значительный путь и часто выходят из ганглия через нервы противоположной стороны, образуя по средней линии перекрест с отростками симметричных клеток. По пути аксоны таких клеток дают множественные боковые ответвления. Возможно, данные нейронывыполняют не только двигательные функции, но частично и ассоциативные, так как могут получать импульсы с обеих сторон ганглия. Полифункциональные ассоциативно-двигательные нейроны обнаружены у олигохет и полихет. В ходе филогенеза число поливалентных клеток убывает, и у высших беспозвоночных нейроны приобретают более узкую специализацию. Центральную часть сегментарного ганглия у дождевого червя занимает основной нейропиль. Его главная функция состоит в образовании связей между двигательными и чувствительными элементами ганглия. Еще вентральнее под основным нейропилем расположен слой чувствительного нейропиля ганглия. Он состоит из множества афферентных волокон, поступающих с периферии от механо-, хемо-, термо- и светочувствительных клеток, расположенных в эктодермальном эпителии и в толще кожно-мускульного мешка. В пределах ганглия чувствительные волокна могут концентрироваться в пучки и образуют коллатерали с тончайшими терминальными разветвлениями. Таким образом, нейропиль сегментарного ганглия представляет собой дифференцированную структуру, гомологичную по функции спинному мозгу позвоночных животных. На уровне сегментарных ганглиев осуществляются координация и регуляция двигательной активности кольчатых червей. Например, локомоция дождевых червей связана с координированной работой кольцевых и продольных мышц, поочередное сокращение которых вызывает распространение перистальтической волны вдоль тела животного. В данном случае кольцевые и продольные мышцы выступают в роли антагонистов, когда сокращение первых совпадает по времени с расслаблением вторых и наоборот. Механизм реципрокного торможения мышц-антагонистов у аннелид, по-видимому, имеет центральную природу и возникает на уровне сегментарного ганглия. Вместе с тем, результаты экспериментов О.Ф. Давида (1990) свидетельствуют о том, что у кольчатых червей различных классов могут существовать специфические формы нервного контроля перистальтических движений. У олигохет (дождевой червь) периодические спонтанные сокращения кожно-мускульного мешка не обусловлены пусковыми влияниями центральной нервной системы, так как они сохраняются и после удаления из препарата брюшной нервной цепочки. Очевидно, у этих животных существует периферический механизм запуска перистальтических сокращений, который приурочен к афферентным и эфферентным элементам субэпителиальной нервной сети. Выключение периферических чувствительных элементов различными анестезирующими веществами приводит к полному подавлению периодических спонтанных сокращений мышц земляного червя. Этот факт свидетельствует о нейрогенной природе механизма запуска, который реализуется в форме своеобразного периферического рефлекса, замыкающегося при переходе возбуждения с рецепторов прямо на сократительные элементы. Центральная нервная система в данном случае оказывает на периферический аппарат регулирующие воздействия, которые ограничиваются способностью усиливать или, наоборот, ослаблять мышечную активность. Естественно, что такое заключение относится к наиболее простым видам локомоции, местным двигательным реакциям. Реализация же быстрых рефлекторных реакций в ответ на раздражение рецепторов головной или хвостовой части обеспечивается у олигохет участием системы гигантских нервных волокон и связанных с ними мотонейронов. Указанная система позволяет животному мгновенно реагировать на раздражение сокращением продольных мышц и, как следствие, удалением переднего (или заднего) конца тела от раздражителя. В данном случае имеют место пусковые влияния центральной нервной системы на локомоторный аппарат. В отличие от олигохет у пиявок и полихет деятельность соматической мускулатуры в большей мере определяется центральными механизмами, в которых задействованы командные триггерные нейроны. Удаление брюшной нервной цепочки у этих животных нарушает координированное плавание, и ритмическая активность мышц выключается. Таким образом, экологические особенности полихет и пиявок сочетаются с более совершенными и быстрыми механизмами запуска и контроля плавательных движений. Вопрос о функциях нейронных цепей в сегментарных ганглиях кольчецов связан с проблемой межнейронного взаимодействия. Как упоминалось ранее, в брюшной нервной цепочке олигохет идентифицированы электрические синапсы. Однако наши представления о процессах интеграции в ЦНС кольчатых червей были бы односторонними, если не остановиться на характеристике другой категории синаптических контактов - синапсов с химическим механизмом передачи. С помощью современных гистохимических и микроэлектрофизиологических методов удалось выделить набор биологически активных веществ, выполняющих медиаторную функцию в ЦНС кольчатых червей. Из числа биогенных моноаминов в брюшной нервной цепочке аннелид обнаружен серотонин в значительной концентрации. Синтез серотонина осуществляется ограниченным числом ганглионарныхнейронов, среди которых следует отметить клетки Ретциуса пиявки. Они представляют собой гигантские (диаметром 74-80 мкм) нейросекреторные клетки, расположенные на вентральной стороне каждого сегментарного ганглия, и относятся к эффекторным элементам, которые участвуют в регуляции секреции слизи из кожных желез. В соме одной клетки Ретциуса содержится 1, 3-4, 1 ммоль серотонина. Ионофоретическая аппликация серотонина (1× 10-5 моль/л) на иннервируемые этими клетками эффекторные органы (слизистые железы) сопровождается выраженным потенцирующим эффектом. Гигантские клетки Ретциуса - весьма популярные у нейрофармакологовтест-объекты. На мембране этих клеток обнаружены рецепторы, чувствительные к катехоламинам: дофамину, норадреналину.Под действием норадреналина возникают деполяризация усиление импульсной активности, генерируемой клетками Ретциуса. Микроаппликация дофамина вызывает полярный эффект и тормозит разряды этих клеток. Гистохимическими методами (методом Фалька-Хилларпа) в сегментарных ганглиях кольчецов были идентифицированы катехоламинергические нейроны по специфическому зелёному свечению при просмотре под люминесцентным микроскопом. Кроме биогенных аминов в ЦНС кольчатых червей обнаружены медиаторные вещества холинергической природы. В мотонейронах аннелид используется главным образом ацетилхолин в качестве возбуждающего медиатора. Микроионофоретическая аппликация ацетилхолина на мембрану клеток Ретциуса вызывает деполяризацию и усиление импульсной активности, генерируемой этими нейронами. Фармакологический анализ данной реакции с использованием различных холинолитиков и холиномиметиков свидетельствует о том, что на мембране клеток Ретциуса имеются холинорецепторы двух видов - никотиновые и мускариновые. Следует отметить, что особый интерес к гигантским клеткам Ретциуса обусловлен также злектротонической связью между этими парными нейронами сегментарного ганглия. Связь определяется электрическим синапсом между аксонами клеток Ретциуса. Данный синапс не обладает выпрямляющими свойствами и создает предпосылки для синхронных разрядов обеих клеток при одновременной регистрации их электрической активности. Этот факт представляет интерес в эволюционном плане, так как указывает на сосуществование различных механизмов транссинаптической передачи на ранних этапах филогенетического развития животного мира. В гомогенатах нервной ткани аннелид выявлено довольно высокое содержание L-глютаминовой и g-аминомасляной (ГАМК) кислот. Имеются физиологические данные о возможном участии этих аминокислот в транссинаптических процессах. Так, например, L-глютаминовая кислота вызывает деполяризацию и заметное усиление активности клеток Ретциуса и механосенсорных нейронов пиявки. ГАМК обладает выраженным действием на возбуждающие и тормозные мотонейроны, иннервирующие соматическую мускулатуру пиявок. Анализируя приведенные выше факты, следует подчеркнуть, что набор медиаторных веществ (которые, к сожалению, не всегда соответствуют всем критериям идентификации синаптических передатчиков) у аннелид весьма обширен и по спектру приближается к набору синаптических посредников у других групп беспозвоночных и позвоночных животных. Это говорит об относительном консерватизме физиологической основы транссинаптической передачи, возникающей на ранних этапах эволюции. Процесс цефализации у кольчатых червей выражен в большей степени, чем у плоских червей. У олигохет надглоточный ганглий расположен над дорсальной поверхностью глотки и состоит из двух слившихся симметричных половин (рис. 4). Окологлоточными коннективами надглоточный ганглий (НГ) связан с подглоточным (ПГ), и в целом эти образования формируют окологлоточное нервное кольцо. Через передние нервы в НГ дождевого червя поступают афферентные волокна - отростки чувствительных клеток, тела которых расположены в эпителии переднего конца тела. В числе этих волокон есть афференты от примитивных светочувствительных органов. Кроме афферентных волокон в составе передних нервов есть и эфферентные. Они проходят по коннективам из подглоточного ганглия через надглоточный. Окологлоточные коннективы содержат нервные тракты, связующие НГ с ПГ. Посредством шести-семи перемычек окологлоточные коннективы соединены с симметричными окологлоточными ганглиями, в которых находятся центры вегетативной нервной системы.
Рис. 4. Строение головных ганглиев дождевого червя (по: Г. А. Невмывака, 1966): 1 - подглоточный ганглий, 2 - коннектив, 3 - окологлоточные ганглии, 4 - надглоточный ганглий, 5 - мышцы глотки, 6 - чувствительные нервные клетки в эпителии глотки Внутренняя структура НГ дождевого червя характеризуется слабой дифференциацией. Его наружный слой образован мелкими веретенообразными клетками, число которых достигает 500. Все эти клетки ассоциативные и связывают различные зоны НГ, образуя его внутренний аппарат. Моторных клеток в НГ не обнаружено. Центральная часть НГ занята нейропилем - переплетением отростков клеток ганглия, а также афферентных волокон из передних нервов. Помимо мелких клеток в НГ дождевого червя содержатся примерно 50 крупных (до 30 км), отростки которых выходят из ганглия через коннективы или через перемычки окологлоточного ганглия. Иногда аксоны крупных клеток проходят через ганглий и попадают в коннективу противоположной по отношению к клеточному телу стороны. Вероятно, эти нейроны относятся к сложным ассоциативным нейронам поливалентного типа. У полихет НГ более дифференцирован, чем у олигохет, и этот процесс сопряжен с развитием органов чувств на головном конце тела. Дифференциация проявляется в разделении НГ полихет на три функциональных отдела. Передний отдел представлен сенсорной зоной нейропиля, куда проецируются афференты от сенсорных нейронов пальп. К среднему отделу подходят оптические нервы от глаз. Зрительные афференты оканчиваются в нейропиле среднего отдела НГ. Наконец, задний отдел НГ полихет получает проекции от затылочного (нухального) органа, выполняющего хеморецепторную функцию. Наиболее существенная особенность в строении НГ полихет состоит в появлении стебельчатых, или грибовидных, тел, которые выполняют функцию ассоциативного центра. Грибовидные тела (ГТ) представлены компактной массой мелких клеточных тел и состоящим из клеточных отростков стебельком. Отростки с большим числом боковых ветвей заканчиваются в стебельке с той же стороны, что и клеточное тело. Однако имеются и проекционные отростки, которые проходят к различным отделам НГ и могут собирать информацию от сенсорных зон нейропиля этих отделов. У сидячих полихет (без активного поиска пищи) ГТ нет и строение мозга значительно упрощено. Удаление НГ у аннелид существенно нарушает приспособительный характер их реакций. Земляные черви без НГ роют норку медленно и неловко. Нарушается нормальный контакт вентральной стороны тела с субстратом, животные становятся беспокойными и более подвижными. У пиявок после удаления НГ наблюдается усиление моторной активности и рефлекса присасывания. У медицинской пиявки увеличивается частота импульсов кольцевых нервов после экстирпации НГ. Все эти факты позволили прийти к заключению, что НГ у аннелид участвует в коррекции приспособительных реакций, а также играет регулирующую роль по отношению к сегментарным моторным центрам. Тормозные нисходящие воздействия НГ снижают возбудимость сегментарных мотонейронов. Удаление НГ снимает эти воздействия, вызывает увеличение частоты импульсных разрядов в кольцевых нервах и, как следствие, усиление локомоторной активности. К важным функциям НГ относится нейросекреторная. У олигохет часть нейросекреторных клеток НГ вырабатывают гормон, тормозящий развитие половых желез и регулирующий процесс размножения. Другая группа нейросекреторных клеток НГ продуцирует гормон, необходимый для нормального течения процесса регенерации. Как центр нервной и гуморальной интеграции НГ аннелид имеет отношение к процессам обучения у этих животных. В этом плане представляет интерес сопоставление сложности поведенческих реакций с дифференциацией НГ у представителей различных классов кольчатых червей. Земляные черви в условиях лабиринтной методики обучаются правильно выбирать рукав Т-образного лабиринта при сочетании неправильного выбора с ударом электрического тока. Выработанная реакция отличается слабой стойкостью, и процент правильных решений зависит от силы безусловного раздражителя (при токе 5 В 54% положительных реакций, а при токе 18 В 85% соответственно). Существенно, что удаление НГ не отражается на динамике обучения. В то же время у полихет Nereis pelagica можно выработать довольно стойкий пищевой условный рефлекс при использовании в качестве условного раздражителя вибрации. По данным Л.Г. Воронина и др. (1972), после 30-50 сочетаний условного раздражителя с пищевым подкреплением вибрация приобретала сигнальное значение, и животные проявляли четкую поисковую реакцию, выходя из стеклянной трубки. При этом наблюдался высокий максимальный процент положительных реакций (80-100%) и длительность их сохранения достигала 6-15 дней. Стабильность условной рефлекторной реакции у полихет коррелирует с появлением в их НГ грибовидных тел, которых нет в слабо дифференцированном НГ земляных червей. Таким образом, отмечается соответствие между структурной дифференциацией НГ и возрастанием его интегрирующей роли в процессах обучения. |
Последнее изменение этой страницы: 2017-05-05; Просмотров: 999; Нарушение авторского права страницы