
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Развитие диэнцефало-теленцефальной системы интеграции у земноводных
Переход от водного образа жизни к наземному повлек за собой целый ряд изменений в сфере организации сенсорных систем и центральных нервных образований. Так, например, у амфибий в связи с изменением типа локомоции в спинном мозге формируются два утолщения, соответствующие верхнему и нижнему поясам конечностей. В спинальных ганглиях вместо биполярных чувствительных нейронов, которые встречаются у рыб, сосредоточиваются униполярные клетки с Т-образно ветвящимся отростком, обеспечивающим более высокую скорость проведения возбуждения, без участия клеточного тела. Внутреннее строение спинного мозга амфибий также претерпевает определенные преобразования. Состоящее из нервных клеток серое вещество, сконцентрированное в окружности центрального канала, дифференцируется на дорсальные и хорошо выраженные вентральные рога, где; локализованы тела мотонейронов. Восходящие пути спинного мозга амфибий расположены в латеральных и дорсальных столбах белого вещества. В системе латеральных столбов выделяют дорсо- и вентролатеральные отделы. Волокна дорсолатерального отдела проецируются в продолговатый мозг, мозжечок и в крышу среднего мозга. Волокна вентролатерального отдела оканчиваются в базальных отделах продолговатого и среднего мозга. На базе данной системы в ходе филогенеза далее формируется спиноталамический тракт, проводящий информацию от тактильных, болевых и температурных рецепторов. Наибольшие преобразования наблюдаются в системе дорсальных столбов белого вещества. Они утолщаются, обособляются от латеральных столбов. В дорсомедиальном отделе продолговатого мозга дифференцируются зачаточные ядра дорсальных столбов, в том числе тонкого пучка Голля nucleus gracilis. Окончания восходящих волокон системы дорсальных столбов обнаружены в покрышке среднего мозга и в каудальной части таламуса. Нисходящие пути ретикулоспинальной системы амфибий по сравнению с таковыми у рыб претерпевают относительно небольшие изменения, это филогенетически консервативные элементы ЦНС. Изменение среды обитания накладывает отпечаток и на морфофункциональную организацию продолговатого мозга амфибий. С выходом животных на сушу орган боковой линии утрачивает свою сенсорную роль. У бесхвостых амфибий этот процесс сопровождается редукцией медиального ядра вестабуло-латеральной системы продолговатого мозга. На его месте формируется кохлеарное дорсальное крупноклеточное ядро, где осуществляется анализ информации, поступающей от примитивного органа слуха. От этого ядра идут пути в ретикулярную формацию заднего мозга к верхней оливе, а также восходящие пути в крышу среднего мозга. В ходе онтогенеза у бесхвостых амфибий изменяются функциональные особенности нейронов дыхательного центра. У головастиков основным раздражителем дыхательного центра выступает низкое содержание кислорода в водной среде, и в этом отношении они сходны с пресноводными рыбами. У взрослых лягушек ведущим фактором, влияющим на возбуждение инспираторных и экспираторных нейронов продолговатого мозга, является содержание углекислоты во внешней среде. Таким образом, переход к наземным условиям жизни отражается на функциях дыхательного центра и его элементов. Этот факт можно объяснить рекапитуляцией − повторением в эмбриогенезе современных организмов признаков, имевших место у взрослых предков. Выход на сушу сопряжен также со значительными преобразованиями в структуре и функциях мозжечка. У бесхвостых амфибий мозжечок меньше, чем у рыб (рис. 14). Это частично обусловлено уменьшением объема аурикулярных долей, куда у рыб проецируются афференты от органов боковой линии. Остаточные от аурикул образования имеют связи с афферентными системами иного происхождения, а именно с сенсорными путями, идущими от вестибулярного аппарата. Вместе с тем у бесхвостых амфибий значительного развития достигает тело мозжечка, анатомически связанное со спиноцеребеллярным трактом, который передает информацию от проприорецепторов мышц туловища, и конечностей. К афферентным связям мозжечка амфибий относятся также вестибуло-церебеллярный, бульбоцеребеллярный тракты, связи с ядрами седьмой и пятой пар черепномозговых нервов, с крышей среднего мозга и гипоталамусом.
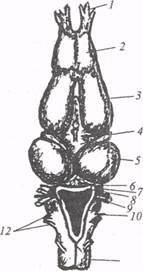
Рис. 14. Головной мозг лягушки Капаesculenta (по: Е.К.Сепп, 1959): 1 − обонятельный тракт, 2 − обонятельная доля, 3 − конечный мозг, 4 − промежуточный мозг, 5 − средний мозг (оптические доли), 6 − мозжечок, 7 − пятый черепномозговой нерв, 8 − седьмой черепномозговой нерв, 9 − восьмой черепномозговой нерв, 10 − продолговатый мозг, 11 − спинной мозг, 12 − девятый - одиннадцатый черепномозговые нервы Эфферентные проекции мозжечка формируют: церебелло-спинальный тракт, волокна, идущие к ядрам черепномозговых нервов, к среднему и промежуточному мозгу. По прямым и опосредованным связям в мозжечок бесхвостых амфибий может поступать информация от дистантных рецепторов, приобретающих особое значение в условиях жизни на суше. Так, например, с помощью электрофизиологических методов в мозжечке лягушек обнаружены нейроны, дающие четкие импульсные реакции в ответ на выключение и включение света. Таким образом, получая информацию от разнообразных рецепторов, мозжечок амфибий участвует в координации двигательной активности. Удаление мозжечка у лягушек вызывает отчетливые изменения моторики, которые проявляются в искаженной форме прыжков и в нарушении содружественных движений конечностей при плавании. Оценивая в целом филогенетические преобразования в мозжечке амфибий, следует отметить, что они сводятся к качественному изменению связей и к вовлечению этой части мозга в более тонкие регуляторные процессы. Средний мозг у амфибий не претерпевает коренных перестроек по сравнению со средним мозгом нижестоящих по уровню эволюционного развития позвоночных животных. Крыша среднего мозга у амфибий, так же как и у рыб, построена по экранному принципу. В верхних слоях тектума обнаружено широкое представительство зрительных волокон, проекции которых имеют топическую организацию, т. е. каждый квадрант сетчатки посылает свои афференты в тектум, «точка в точку». Нейроны тектума лягушки и жабы приспособлены для детекции специальных видов движения и пространственных отношений. Животные могут автоматически классифицировать внешний стимул и отфильтровать все его частные особенности. Так, например, некоторые из нейронов настроены на восприятие мелких темных движущихся предметов («детекторы жуков»), объектов охотничьего поведения. Функция нейронов-детекторов сводится к запуску соответствующей поведенческой реакции. После удаления тектума лягушки и жабы не становятся абсолютно слепыми, как полагали раньше, но у них нарушаются пищевые реакции. Животные перестают реагировать хватанием на движущиеся пищевые объекты, однако у оперированных амфибий сохраняется способность огибать неподвижные предметы и спасаться от угрожающих стимулов. У бесхвостых амфибий − животных с развитой акустической коммуникацией, в среднем мозге локализован слуховой центр − полукружный торус, torus semicircularis. Он представляет собой парный (как и у рыб) вырост тегментума и состоит из основного крупноклеточного ядра и окружающей его зоны реже расположенных клеток. По данным Н.Г. Бибикова (1974), в расположении клеток основного ядра отмечается определенная упорядоченность, т.е. в этом ядре, возможно, существует тонотопическая организация. Для подтверждения этой гипотезы необходимы дополнительные данные о локализации нейронов с определенной частотной настройкой. При внутриклеточной регистрации ответов одиночных нейронов полукружного торуса лягушки на акустическую стимуляцию выявлены клетки, реагирующие на интенсивность и длительность звука в сравнительно узком диапазоне. Значения воспринимаемых частот для нейронов полукружного торуса у лягушки-быка составляют 100-5000 Гц, причем максимальная чувствительность обнаружена в области частот 200-1500 Гц. Между нейронами слухового центра лягушки описаны сложные возбудительно-тормозные взаимодействия, в значительной мере сходные с межнейронными взаимодействиями в задних холмах среднего мозга млекопитающих. Восходящие пути, идущие от слухового центра среднего мозга в конечный, у бесхвостых амфибий изучены недостаточно, хотя предполагается связь полукружного торуса лягушек с центральным таламическим ядром и далее с ядрами стриатума и медиального паллиума в конечном мозге. Таким образом, средний мозг амфибий, как и у других позвоночных, служит первичным координационным центром, возникшим как надстройка над системами дистантной рецепции. Этот центр представляет собой субстрат для реализации врожденных поведенческих реакций. Вместе с тем пластичные формы поведения обеспечиваются с участием филогенетически более молодого координационного центра в конечном мозге амфибии, где наблюдается ряд существенных морфофункциональных преобразований. Прогрессивное развитие конечного мозга в ходе филогенеза обусловлено дифференциацией релейных центров промежуточного мозга, через которые афферентная информация поступает в конечный мозг. Промежуточный мозг амфибий состоит из эпиталамуса, таламуса и гипоталамуса. Эпиталамус представлен габенулярными ядрами, которые, как и у рыб, получают многочисленные тракты от конечного мозга. Таламус амфибий состоит из двух основных отделов − филогенетически древнего вентрального и более молодого дорсального. У высших бесхвостых амфибий дорсальный отдел преобладает над вентральным, относительно мало дифференцированным. У бесхвостых амфибий по сравнению с нижестоящими позвоночными в дорсальном таламусе наблюдаются более интенсивная миграция нейронов из перивентрикулярного положения в нейропильные поля и образование истинных ядер, которые весьма различны по структуре и могут иметь как нейропильную, так и клеточную организацию. Согласно представлениям Дж. Херрика, эту стадию развития таламуса можно рассматривать как зауропсидную. Законченную форму эта стадия приобретает у рептилий. Дорсальный таламус бесхвостых амфибий имеет обширные двусторонние связи с тектумом среднего мозга. В средних отделах дорсального таламуса переключается ретинотектоталамотеленцефальный тракт, передающий зрительную информацию от сетчатки и формирующий проекции в базальных ганглиях (стриатуме) конечного мозга. Этот канал связи полимодальный; возможно, он сопоставим с неспецифическими таламическими входами высших позвоночных. В пользу этой точки зрения свидетельствуют следующие факты. Зоны таламического представительства зрительной и соматосенсорной афферентных систем у лягушек в значительной степени перекрываются. Практически во всех структурах таламуса, где зарегистрированы соматические вызванные потенциалы, обнаружены электрические реакции на зрительное раздражение. Исключение составляют такие нейропильные образования дорсального таламуса амфибий, как наружное коленчатое тело и ядро Беллончи, где сосредоточено наибольшее количество нейронов, реагирующих только на зрительное раздражение. Представляет интерес тот факт, что электрические реакции, возникающие в конечном мозге при раздражении соматическими и зрительными стимулами, сохраняются после разрушения тектума. Следовательно, у амфибий на подступах к конечному мозгу существуют две афферентные системы: филогенетически древний путь с переключением в крыше среднего мозга (ретинотектотала-мотеленцефальный) и другой, более прямой с переключением в таламусе (ретиноталамотеленцефальный). В результате нейроморфологических и электрофизиологических экспериментов, которые проводили Л.М. Номоконова, Н.П. Веселкин, М.Г. Белехова, А.Л. Агаян и др. (1971), установлено, что ретиноталамотеленцефальный путь у амфибий переключается в переднем ядре дорсального таламуса. Это ядро помимо прямых ретинальных проекций получает гипоталамические проекции и афференты соматической модальности. Через медиальный пучок переднего мозга переднее ядро дорсального таламуса проецируется в зачаточный гиппокамп, который принято рассматривать как гомолог гиппокампальной формации млекопитающих. Таким образом, у амфибий формируются два канала для проведения зрительной информации к конечному мозгу − опосредованный (через тектум) и прямой (через таламус), причем принцип двойной иннервации характерен и для соматической афферентной системы. Вопрос о соотношении числа древних и новых каналов, о корреляции их функций пока остается одним из наиболее сложных в эволюционной нейрофизиологии. Гипоталамус у амфибий, исключая преоптическую область, разделен на две части: дорсальную, тесно связанную с малодифференцированным вентральным таламусом, и вентральную. Отметим, что вентральная часть сравнима с серым бугром млекопитающих и образует функциональное единство с нейросекреторным преоптическим ядром. Гипоталамус амфибий имеет анатомические связи с таламусом, стриатумом, конечным, средним и продолговатым мозгом; это один из афферентных источников для корковых формаций конечного мозга. При локальном раздражении переднего и заднего гипоталамуса вызванные потенциалы (ВП) регистрируются в палеокортексе (зачаточной пириформной коре) и архикортексе (зачаточном гиппокампе). В нейросекреторном преоптическом ядре бесхвостых амфибий намечаемся обособление двух групп клеток − дорсальной и вентральной. Из этих групп клеток у высших позвоночных формируются паравентрикулярное и супраоптическое ядра. Отростки секреторных клеток преоптического ядра образуют преоптикогипофи-арный тракт, направляющийся в гипофиз. У амфибий впервые в ряду позвоночных отмечается компактный нейрогипофиз, в котором различают срединное возвышение, инфундибулярную часть и главную заднюю часть. Конечный мозг амфибий морфологически и функционально связан в единую систему с промежуточным мозгом. Филогенетическое развитие этих образований шло параллельно и привело к возникновению новой таламотеленцефальной системы интеграции. В качестве ведущего фактора в формировании теленцефального интеграгивного комплекса выделяют афферентное снабжение, которое имеет обонятельные, гипоталамические и таламические источники. Эти источники играют существенную роль в обособлении зачаточных корковых формаций конечного мозга: палео-, архи- и неопаллиума. Зачаток древней, эбонятельной коры − палеопаллиум, у амфибий занимает дорсолатеральную часть полушария (рис. 15) и состоит из мультиполярных звездчатых клеток и грушевидных нейронов. дендриты которых тянутся дорсально к клеткам дорсального паллиума и латерально к волокнам дорсального ольфакторного тракта. Аксоны грушевидных нейронов либо синаптируют с нейронами соседних зон плаща, либо идут в вентральном направлении и, пересекая стриарную область, входят в нисходящие пути. Вентральная часть палеопаллиума граничит с полосатым телом (стриатумом). При электрическом раздражении латеральной части обонятельной луковицы в палеопаллиуме лягушки регистрируются медленные поверхностно негативные вызванные потенциалы (ВП), свидетельствующие о наличии обонятельных прюекций в этой области.
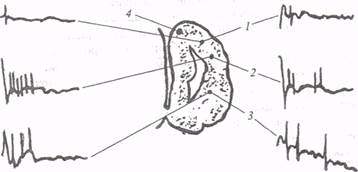
Рис. 15. Зачаточные корковые формации конечного мозга лягушки (поперечный срез) (по: А. И. Карамян. 1975): 1 − дорсальный участок плаща (зачаточная новая кора), 2 − зачаток древней, обонятельной коры (палеопаллиум), 3 − полосатое тело (стриатум), 4 − зачаточная архикортикальная формация (гиппокампальный паллиум); слева − нейрональные реакции формаций конечного мозга при раздражении переднего гипоталамуса; справа − то же при раздражении заднего гипоталамуса: частота раздражения 1 стимул в 15 с. Вместе с тем при локальном раздражении центральных зон обонятельной луковицы ВП появлялись и в дорсомедиальной части полушария, где расположен гиппокампальный паллиум. Эта зачаточная корковая формация амфибий наиболее развита, она состоит из звездчатых, биполярных и довольно крупных пирамидоподобных клеток. Аксоны пирамидоподобных клеток могут распространяться вентрально вдоль стенки желудочка, к мозговой перегородке или идти дорсально к дорсальному паллиуму. Как отмечалось выше, гиппокампальный паллиум амфибий отличает афферентация различной модальности. Сюда через гипоталамус поступает информация от висцеральных рецепторов (рис.15), через таламус − от экстерорецепторов. Наконец, в зачаточном гиппокампе обнаружены обонятельные проекции. Таким образом, в гиппокампальном паллиуме (архипаллиуме) осуществляется интеграция висцеральной и экстероцептивнои сенсорной информации, и в этом плане его функция сопоставима с функцией лимбической системы высших позвоночных, а именно с оценкой биологической важности сенсорной информации и запуском соответствующих соматовегетативных реакций. Дорсальный участок плаща (дорсальный паллиум) амфибий, который гомологизируется с неокортексом высших позвоночных, представляет собой наименее дифференцированную область зачаточных корковых формаций. Звездчатые и грушевидные клетки этой области расположены перивентрикулярно. Их отростки направляются либо медиально, в сторону гиппокампального паллиума, либо латерально, образуя синаптические контакты с нейронами палеопаллиума. Границы дорсального паллиума установить трудно, так как он представляет собой переходную зону между палео- и архипаллиумом. Вместе с тем на основании структурной организации этой зачаточной зоны можно допустить ее участие в ассоциативной функции, в интеграции импульсов, поступающих из соседних формаций архи- и палеопаллиума. Казалось бы, тенденция к обособлению трех зачаточных формаций коры головного мозга у амфибий должна сочетаться с большей пластичностью поведения. Однако результаты исследования обучения у амфибий свидетельствуют о том, что оборонительные условные рефлексы у этих животных вырабатываются с большим трудом.Оборонительный условный рефлекс на световой или звуковой раздражитель у травяных лягушек образуется после 15-30 сочетаний, но он весьма нестойкий и не стабилизируется даже после 10-15 дней опытов. В некоторых случаях трудности, которые встречаются при исследовании приобретенных форм поведения земноводных, обусловлены несовершенством методики, в частности использованием мало адекватных видов раздражения. Так, например, при выработке у бесхвостых амфибий условнорефлекторных реакций на звук болевая стимуляция вызывает затаивание и не приводит к активной реакции избегания. Эффективные методы поощрения пока тоже не выявлены. Рекордсменами в области научения выступают серые жабы, у них удается выработать ассоциацию звукового сигнала с несъедобностью корма, используемого как безусловный раздражитель. Тем не менее, изложенные выше факты не дают достаточного основания считать, что конечный мозг земноводных не принимает участия в осуществлении приспособительных реакций. После удаления полушарий конечного мозга у лягушек и жаб утрачиваются индивидуальные защитные реакции, способность к самостоятельному питанию и запоминанию местонахождения пищи. Оперированные животные не могут приспособиться к жизни в естественных условиях, и это свидетельствует о существенном вкладе конечного мозга в общую стратегию и организацию поведения. Итак, в связи с переходом от водного образа жизни к наземному в ЦНС амфибий имеет место целый ряд преобразований, направленных на совершенствование новых систем интеграции. К числу таких преобразований можно отнести: развитие ядерной структуры дорсального таламуса, гипоталамуса и базальных ганглиев, совершенствование морфологических связей между ядерными образованиями таламуса и конечным мозгом, дальнейшую дифференциацию зачаточных формаций коры головного мозга. Все эти: факторы способствуют становлению таламотеленцефальной системы интеграции, которая получает дальнейшее развитие в головном мозге рептилий. Зауропсидный этап развития Внутренняя структура спинного мозга рептилий претерпевает дальнейшую дифференциацию. Этот процесс выражается в обособлении дорсальных и вентральных рогов серого вещества, сужении просвета центрального канала, стенки которого значительно утолщаются. Белое вещество спинного мозга подразделяется на передние, задние и латеральные продольные столбы. Часть длинных восходящих волокон дорсальных столбов проходит до продолговатого мозга, где они оканчиваются в ядрах, идентичных ядрам Голля и Бурдаха млекопитающих. Проекции восходящих волокон дорсальных столбов имеют топическую организацию, и по этому признаку они также весьма близки к соответствующим трактам млекопитающих. Наиболее дифференцированы дорсальные столбы и их ядра у аллигаторов, в меньшей степени у черепах и змей. Длинные восходящие волокна латеральных столбов оканчиваются на разных уровнях мозгового ствола рептилий. Их проекции обнаружены в ретикулярной формации продолговатого мозга, в мозжечке, крыше среднего мозга, в перивентрикулярном сером веществе. В составе латеральных столбов, у рептилий проходит палеоспиноталамический тракт, который проводит информацию о тактильной чувствительности и обусловливает, дальнейшее совершенствование локомоции. Параллельно эволюции восходящих путей происходит развитие нисходящих трактов спинного мозга. В среднем мозге рептилий концентрируется группа крупных клеток, образующих красное ядро (n. ruber). От этого ядра начинается нисходящий руброспиналъный тракт, проходящий в латеральных столбах спинного мозга. Руброспинальный тракт наряду сцеребелло-спинальным, ретикулоспинальным, вестибуло-спинальным трактами составляет одно из завершающих звеньев экстрапирамидной системы. С помощью этой системы влияния конечного мозга, ретикулярной формации ствола, мозжечка, ядер вестибулярного комплекса передаются на мотонейроны спинного мозга. Таким образом, экстрапирамидная система участвует в управлении движением. У рептилий как истинно наземных животных возрастает роль акустической информации, возникает необходимость ее сопоставления со зрительной, обонятельной и вкусовой. Новые экологические факторы стимулируют преобразования как в структуре акустических рецепторов, так и в системе их центрального представительства. В продолговатом мозге рептилий в дополнение к дорсальному крупноклеточному ядру формируются угловое (n. angularis) и пластинчатое (n. laminaris) ядра, имеющие непосредственное отношение к слуховой функции. Все три ядра занимают довольно протяженную статоакустическую область продолговатого мозга. Аксоны нейронов углового ядра идут к ипсилатеральной верхней оливе и в составе латеральной петли к полукружному торусу среднего мозга. Оттуда путь продолжается в дорсальную область таламуса и далее в конечный мозг. Мозжечок у рептилий, в общем, развит сильнее, чем у амфибий (рис. 16), однако его структурная организация обладает видовой спецификой. Наиболее дифференцирован мозжечок у крокодилов. Мозжечок этих животных продольными бороздами поделен на переднюю, среднюю и заднюю доли. Намечается увеличение тела мозжечка за счет разрастания его боковых долей (flocculi), которые в ходе дальнейшей эволюции превращаются в полушария. У рептилий имеются собственные ядра мозжечка: крупноклеточное медиальное и мелкоклеточное латеральное. Первое гомологично фастигиальному, а второе − промежуточному ядру мозжечка млекопитающих. Мозжечок рептилий имеет афферентные и эфферентные связи со средним, продолговатым и спинным мозгом. Степень развития мозжечка разных рептилий находится, по мнению ряда исследователей, в зависимости от способа локомоции. У змей, которые лишены конечностей, тело мозжечка значительно меньше, чем у крокодилов и черепах.
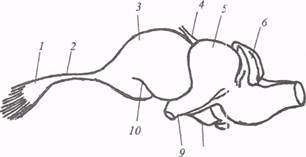
Рис. 16. Мозг ящерицы Varanus griseus (по: Е.К.Сепп, 1959): 1 − обонятельная луковица, 2 − обонятельный тракт, 5 − полушарие конечного мозга, 4 − эпифиз, 5 − крыша среднего мозга, 6 − мозжечок, 7 − гипофиз, 8 − воронка (infundibulum), 9 − хиазма, 10 − fovea limbica С помощью электрофизиологических методов установлено, что в мозжечке рептилий имеются не только вестибулярные и соматические проекции, но и представительство дистантных рецепторов, в частности зрительных. По данным А.И. Карамяна, в коремозжечка серого варана (Varanus griseus)некоторые нейроны в ответ на раздражение зрительного нерва реагируют учащением импульсных разрядов. К сожалению, природа данных нейрональных реакций окончательно не выявлена, хотя априори они свидетельствуют о важном вкладе экстерорецепторов в процессы сенсомоторной интеграции. Удаление мозжечка у ящериц приводит к нарушению ритмичных движений конечностей, однако общие последствия данной операции не соответствуют триаде Лючиани: атония, астения, астазия, характерной для млекопитающих. Дело в том, что у рептилий экстирпация мозжечка вызывает не только атонию (гипотонию), но и гипертонию мышц локомоторного аппарата. У рептилий, так же как у амфибий, средний мозг сохраняет свое значение как интегративный стволовый центр, который получает обширные проекции зрительной, слуховой, соматической сенсорных систем и посылает эфферентные пути к нижележащим моторным центрам. У крокодилов к передним буграм четверохолмия присоединяются полностью сформированные задние, куда подходят афференты слухового пути. Самая мощная афферентная система крыши среднего мозга (тектума) — зрительный тракт заканчивается в поверхностных слоях тектума. Структурная дифференцировка тектума у рептилий более выражена, чем у амфибий. Он состоит из 14 слоев, которые обычно объединяют в шесть, как и у всех позвоночных. Зрительные волокна оканчиваются преимущественно в верхних слоях, причем проекции зрительных афферентов имеют ретинотопическую организацию, наиболее велика их плотность для центральных отделов сетчатки. Нейроны тектума у рептилий участвуют в анализе зрительной информации и обладают избирательной чувствительностью к направлению движущихся объектов. Тектальные нейроны черепах организованы в виде функциональных колонок и участвуют в анализе оптических сигналов по интенсивности и длине волны. В глубоких слоях тектума рептилий обнаружены проекции соматических афферентов, идущих из спинного мозга. Средний мозг рептилий выполняет функции координационного центра, кроме того, это важная «станция» на пути восходящих сенсорных систем к дорсальному таламусу, играющему существенную роль в становлении таламотеленцефальной системы интеграции. Зауропсидная стадия развития таламуса у рептилий характеризуется дальнейшей миграцией клеток из перивентрикулярного слоя и смещением некоторых структур вентрального отдела таламуса в дорсальном направлении. Наряду с этим увеличивается число дифференцированных ядер с диффузной клеточной организацией. Особенно высокого уровня развития достигает дорсальный отдел таламуса рептилий, где можно выделить переднюю, медиальную, вентральную и заднюю группы ядер, сходные с соответствующими ядерными группами таламуса млекопитающих. Проекции различных видов чувствительности концентрируются в определенных зонах таламуса рептилий, например представительство зрительной системы в латеральной зоне. Электрофизиологическими и морфологическими методами М.Г. Белеховой и А.А. Косаревой установлено, что главное ядро этой зоны − латеральное коленчатое тело получает прямые афференты от сетчатки, без синаптического переключения в среднем мозге. Соматическая афферентная система проецируется в вентрокаудальную зону таламуса рептилий, куда приходит бульботаламический тракт. Вместе с тем соматические проекции в этой зоне перекрываются со слуховыми и зрительными. Существует точка зрения, что n. ventralis является предшественником релейных соматических ядер млекопитающих. Другое ядро вентрокаудальной зоны таламуса рептилий − n. reuniens, в ходе эволюции принимает функцию слухового реле млекопитающих. Наконец, расположенное в медиоцентральной зоне таламуса круглое ядро − n. rotundus, лишено прямого ретинального притока, на нейронах этого ядра переключаются зрительные импульсы, идущие из крыши среднего мозга в конечный мозг. Таким образом, зрительная информация в конечный мозг рептилий может поступать по двум каналам: ретиноталамотеленцефальному и ретинотектоталамотеленцефальному. филогенетически более древнему, прослеживающемуся почти у всех позвоночных начиная с хрящевых рыб. Второй путь, опосредованный через тектум, в конечном мозге имеет представительство в подкорковом отделе полушария − гиперстриатуме (дорсальный вентрикулярный край). Несмотря на субпаллиальное положение данная структура в настоящее время по своему эмбриогенезу, гистохимическим характеристикам считается сопоставимой с новой корой млекопитающих. Ретиноталамотеленцефальный зрительный канал, проецирующийся в дорсальную (новую) кору, − филогенетически более молодой тракт. Его нет у низших позвоночных ( круглоротых, рыб), он формируется у рептилий терапсидной линии (современные представители − черепахи). У рептилий второй дивергентной линии − зауропсидной (современные представители − ящерицы), ретиноталамотеленцефальный канал проецируется не в дорсальную кору, а в ростральный отдел дорсального вентрикулярного края. Функционально два описанных выше канала проведения зрительной информации не дублируют друг друга, а обеспечивают различные стороны зрительного восприятия (зрительное внимание и идентификация свойств зрительных объектов). В целом обе системы взаимодействуют и создают предпосылки для целостного зрительного восприятия. Вместе с тем у рептилий имеется еще один путь, через который зрительный и другие сенсорные импульсы передаются в конечный мозг. По данным М.Г. Белеховой, этот путь переключается в переднем дорсолатеральном ядре таламуса и проецируется в гиппокампальную (дорсомедиальную) кору. Этот канал обнаружен у ящериц, есть данные об эквивалентном пути у черепах. Данный путь переключения экстерорецептивной информации гомологичен лимбическому каналу у амфибий и служит для запуска сомато-вегетативных поведенческих реакций. Приведенные выше факты позволяют по-новому подойти к вопросу об источниках афферентного снабжения гиппокампальной коры у рептилий, так как ранее считалось, что основные (висцеровкусовые) проекции имеют место только в гипоталамусе и следуют в гиппокампальную кору через медиальный пучок конечного мозга. В гипоталамусе рептилий, так же как и в таламусе, происходит целый ряд прогрессивных изменений. Они затрагивают в первую очередь структуры заднего гипоталамуса, развитые у рептилий. При разрушении заднего и среднего гипоталамуса дегенеративные изменения нервных волокон наблюдаются в гиппокампальной и пириформной (обонятельной) формациях коры конечного мозга, что свидетельствует о более тесных связях гиппокампальной коры с задним гипоталамусом. В передней области гипоталамуса у рептилий впервые среди позвоночных формируются парные супраоптические и паравентрикулярные нейросекреторные ядра − гомологи преоптического ядра низших позвоночных. Оба ядра у рептилий характеризуются компактным расположением клеточных элементов и богатой васкуляризацией. Аксоны, идущие от ядер, формируют паравентрикуло- и супраоптико-гипофизарные тракты, занимающие, по данным А.Л. Поленова, обширную площадь вентрального гипоталамуса. В пределах серого бугра оба тракта сливаются и образуют единый гипоталамо-гипофизарный тракт, направляющийся в нейрогипофиз. Таким образом, гипоталамус рептилий характеризуется тенденцией к формированию новых ядер, к расширению связей с конечным мозгом и в целом по структуре приближается к гипоталамусу млекопитающих. Конечный мозг рептилий характеризуется переходом от нейропильного типа строения к более дифференцированному с четким структурным разделением паллиальных и субпаллиальных структур и разрастанием латеральной стенки полушария. Субпаллиальные (стриарные) образования рептилий развиваются по ядерному типу и, согласно мнению большинства исследователей, состоят из трех отделов: палеостриатума, неостриатума и гиперстриатума. Палеостриатум занимает в пределах конечного мозга вентральное положение и состоит из двух частей − медиальной и латеральной (рис. 17). Медиальная часть иннервируется волокнами медиального пучка конечного мозга и носит название paleostriatumaugment atum («увеличивающийся древний стриатум»). Эта зона является гомологом медиальной, более древней части головки хвостатого ядра у млекопитающих. Латеральная часть палеостриатума (paleostriatum primitivum) включает в себя бледный шар (globus pallidus), сформированный клетками, аксоны которых направляются к моторным центрам стволовой части мозга. Бледный шар − филогенетически древнее образование − прослеживается у всех позвоночных, начиная с круглоротых. Гомология данной структуры с помощью гистохимических методов установлена у рептилий, птиц и млекопитающих.
А − черепаха Podochemys unifilis, Б − ящерица Jguana iguana, В − аллигатор Alligator jissisipiensis, Г − ящерица Jguana iguana; 1 − дорсальная (общая) кора, 2− паллиальное утолщение, 3 − дорсальный вентрикулярный край, 4 − неостриатум, 5 − палеостриатум, 6− медиодорсальная (гиппокампапьная) кора, 7 − дорсомедиальный отдел общей коры, 8 − дорсолатеральный отдел общей коры, 9− латеральная (пириформная) кора Наиболее дорсальное положение среди субпаллиальных образований занимает гиперстриатум. Идентифицировать его у рептилий различных видов весьма сложно, что объясняет существование противоречивых трактовок. Это проявляется в обилии названий (дорсальный вентрикулярный край, дорсальный стриатум, гиперстриатум и т.д.), которые используют различные авторы для обозначения данной структуры. Согласно одной из наиболее распространенных гипотез, развиваемой такими нейроморфологами, как Б. Келлен, У. Наута, Г. Норткатт и др., дорсальный вентрикулярный край (ДВК) паллиального происхождения и, таким образом, относится к неокортикальной формации. В пользу этой точки зрения приводится целый ряд доказательств. Так, результаты эмбриогенетических исследований свидетельствуют о том, что ДВК рептилий и птиц формируется из гипопаллиальной эпендимы, нейробласты которой разрастаются в структуру ядерного типа. Гипопаллиальная эпендима − это один из зачатков неокортекса, а значит, ДВК имеет паллиальное происхождение. Гистохимическими методами установлено, что ДВК у рептилий, в отличие от палеостриатума и неостриатума, имеет слабую моноаминергическую и холинэстеразную активность. На основании этих данных можно говорить о своеобразии ДВК и исключить данную структуру из стриопаллидарной системы рептилий, имеющей непаллиальное происхождение и характеризующейся нейрохимическим единством. Третья группа фактов, приводимых как аргументы в пользу новых представлений о происхождении ДВК у рептилий, включает сведения о связях этого отдела мозга. Электрофизиологическими и морфологическими методами в ДВК черепах, ящериц и крокодилов обнаружены относительно дискретные проекции зрительной, слуховой и соматической сенсорных систем. Зрительные импульсы, идущие из кры |
Последнее изменение этой страницы: 2017-05-05; Просмотров: 660; Нарушение авторского права страницы
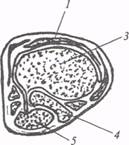 Неостриатум расположен дорсальнее и отделяется от палеостриатума бесклеточной зоной (рис. 17). Согласно эмбриогенетическим и гистохимическим данным, неостриатум рептилий гомологичен комплексу хвостатое ядро − скорлупа в базальных ганглиях млекопитающих. У рептилий, птиц и млекопитающих клетки неостриатума отличаются значительным содержанием моноаминов и холинэстеразы.
Неостриатум расположен дорсальнее и отделяется от палеостриатума бесклеточной зоной (рис. 17). Согласно эмбриогенетическим и гистохимическим данным, неостриатум рептилий гомологичен комплексу хвостатое ядро − скорлупа в базальных ганглиях млекопитающих. У рептилий, птиц и млекопитающих клетки неостриатума отличаются значительным содержанием моноаминов и холинэстеразы.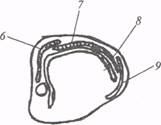
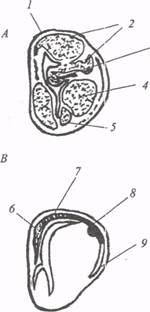 Рис. 17. Подкорковые и корковые отделы конечного мозга у рептилий (по: М.Г. Белехова, 1997):
Рис. 17. Подкорковые и корковые отделы конечного мозга у рептилий (по: М.Г. Белехова, 1997):