|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Песчаные почвы не образуют агрегатов ⇐ ПредыдущаяСтр 5 из 5
Песчаные почвы, такие как эта, – пример зернистой структуры почвы. Крупинки песка относительно крупные и слабо объединяются между собой. Даже те песчаные почвы, где количество коллоидов достаточно большое, под действием давления легко распадаются. Песчаные типы почв с низким уровнем содержания глины требуют более глубокой культивации для создания плодородной структуры и посевного ложа. При низком содержании глины, падает возможный внутренний потенциал образования агрегатов. Глинистые почвы структурируются
Глинистые почвы, такие как эта часто агрегируются и образуют хорошую структуру. Даже при содержании 5%, глина сильно влияет на почву и определяет ее свойства.
Структурированность почвы – это результат ряда совместно протекающих в ней процессов образования агрегатов. Эти процессы формирования структур влияют на состав почвы в динамике с её обработкой. Агрегаты улучшают почву Когда частицы глины соединяются в агрегаты, свойства почвы почти всегда улучшаются. Важное результатом является упрощение агротехнических мероприятий. Также лучше становится аэрация, улучшая транспортировку кислорода и углекислого газа в корневой системе. Агрегированность также увеличивает проницаемость и водоудерживающую способность. Хорошая структурированность почвы увеличивает потенциал посевного ложа, т.к. обеспечены все условия, необходимые для растения и его корней. Структура = Почвенная структура определяется соотношением классов с разным диаметром, в частности относительные пропорции песка, ила и глины, в соответствии с Таблицей «Соотношение частиц по распространенности» Зернистая структура = в почвах, имеющих зернистую структуру, первичные частицы слабо агрегируются, либо этого не происходит вообще. Агрегированная структура = Когда первичные частицы из класса глины, они соединяются друг другом и могут становиться прочнее за счет органического вещества, извести и различных химических осадков. Коллоид = Коллоиды являются лучшими структурными единицами почвы, со средним диаметром менее 0, 0002 мм. Коллоиды включают в себя некоторые органические вещества и настоящую глину. Кислород = элемент, встречающийся в воздухе в форме О2 и концентрации 21% - жизненно необходимо для клеточного дыхания растений и их корней. Диоксид углерода = углекислый газ (СО2) клеточного дыхания корней, который является составной частью, наряду с водой, для образования сахара растением в процессе фотосинтеза. 11. Экологические группы растений по отношению к кислотности почв (ацидофилы, кальциофилы, ацидо-кальциофилы, индифферентные). Ацидофилы (от лат. acidus — кислый + др.-греч. φ ι λ έ ω — люблю) — тип экстремофилов, организмы, обитающие в условиях высокой кислотности. Некоторые жгутиковые и коловратки могут развиваться в массовых количествах в сфагновых болотах при рН воды до 3, 8 и при этом не встречаются в нейтральных и щелочных водоёмах. Типичными ацидофилами являются десмидиевые водоросли, обитающие преимущественно в болотах. К ацидофильным бактериям относятся уксуснокислые и молочнокислые бактерии. Archaea · Sulfolobales · Thermoplasmatales · Acidianus brierleyi, Acidianus infernus — термоацидофилы, факультативные анаэробы · Metallosphaera sedula — термоацидофилы Bacteria · Acidobacterium · Acidithiobacillales · Thiobacillus prosperus, Thiobacillus acidophilus, Thiobacillus organovorus, Thiobacillus cuprinus · Acetobacter aceti Ацидофильные растения[править | править вики-текст] Ацидофильные растения или ацидофиты - растения, произрастающие на кислых почвах. Растения, обитающие на наиболее кислых субстратах (рН 3, 5-4, 5), являются представителями флоры сфагновых болот: клюква, багульник, сфагновые мхи. На сильно кислых почвах могут расти вереск, белоус, щучка извилистая, щавелек малый. На среднекислых и слабокислых почвах (рН 4, 5-6, 5) произрастают полевица собачья, щучка дернистая, погремок большой.
Кальцефилы (кальцефиты) (от лат. calx, родительный падеж calcis — «известь» и др.-греч. φ ι λ έ ω (phileo) — «любить») — растения, обитающие преимущественно на почвах, богатых соединениями кальция, а также в местах выхода известняков, мергелей, мела и других пород. К кальцефилам относятся такие растения, как Пафиопедилум беглый, Пафиопедилюм замечательный[1], лишайники рода Psora: Psora decipiens, Psora globifera, Psora rubiformis и др. По отношению к кальцию различают факультативных (Dendranthema sinuatum, Chamaerhodos altaica, Echinops humilis и др.) и облигатных кальцефилов (Костенец постенный, Brachanthemum krylovianum, Молочай скальный и др.). По отношению к кислотности почвы различают ацидофилы (см. Ацидофильные растения), предпочитающие кислые почвы, нейтрофилы -- растения почв снейтральной реакцией (ясень обыкновенный, сныть обыкновенная), базифилы -- растения щелочных почв(люцерна хмелевидная, тимьян Маршалла, саксаул), индифферентные виды -- растения с широкой экологической амплитудой, живущие в широком диапазонекислотности (сосна обыкновенная). Некоторые растения проявляют положительную реакцию на высокое содержание солей кальция. Это - кальцефилы (лиственница сибирская, бук лесной). Противоположная группа растений -- кальцефобы, произрастающие на болотах и кислых почвах (вереск, водяника). Отношение растений к содержанию кальция в почве, как правило, обратно их отношению к pH почвенного раствора: на карбонатных почвах произрастают базифилы, большинство из которых относится к ацидофобным растениям, или ацидофобам. Напротив, ацидофильные растения являются кальцефобами. По отношению к общему богатству почвы различают следующие экологические группы: олиготрофные растения, или олиготрофы, довольствующиеся небольшим количеством питательных веществ, мезотрофные растения, или мезотрофы, приуроченные к почвам со средней обеспеченностью элементами минерального питания; эвтрофные растения, или эвтрофы, распространенные преимущественно на плодородных почвах. Растения, особо требовательные к повышенному содержанию азота в почве, называют нитрофилами (малина, крапива). От экологических групп следует отличать экологоценотические группы -- группы видов растений, сходных по отношению к совокупности экологических факторов, присущих биотопам того или иного типа. Напр., для лесов Восточной Европы выделены следующие эколого-ценотические группы: бореальная лесная (виды сомкнутых темнохвойных лесов); бореальная опушечная (виды, растущие в «окнах» и на опушках темнохвойных лесов); неморальная лесная (виды сомкнутых широколиственных лесов); неморальная опушечная (виды, приуроченные к «окнам» и опушкам широколиственных лесов); нитрофильная лесная (виды сомкнутых черноольховых лесов); нитрофильная опушечная (виды разреженных черноольховых лесов); боровая лесная (виды сомкнутых сосновых лесов северной части лесной зоны); боровая опушечная (виды разреженных сосновых лесов южной части лесного пояса) и др. 12. Экологическое значение доступного азота почвы. Группы растений по отноше-нию к азоту: нитрофилы и нитрофобы. Облигатные и факультативные нитрофи-лы. Экологическое значение доступного азота почвы Огромное влияние на растительный покров, на экологию и биологию растений оказывают соединения азота, содержащиеся в почве. Считают, что азот атмосферы первоначально находился в твердом веществе Земли в виде соединений аммония и нитридов, т. е. соединений азота с металлами и другими элементами. По мере разогревания Земли азот выделялся в атмосферу в составе аммиака. А когда атмосфера обогатилась кислородом за счет развивающегося фотосинтеза, аммиак стал окисляться до элементарного азота.
Однако концентрация нитратов в почвенном растворе обычно очень низка, в противоположность общему содержанию азота, которое выше в сотни раз. Нитраты быстро потребляются из почвенного раствора, но также быстро и пополняется их количество при минерализации. Азот возвращается в почву преимущественно с опадом листьев, плодов, отмирающими корнями, а также, вероятно, и с корневыми выделениями. Кроме того, он поступает в почву и при отмирании многочисленных почвенных организмов. Все эти азотсодержащие вещества могут накапливаться в почве и обычно включаются в минерализацию; тогда они расщепляются сапротрофами, которые используют в качестве источника энергии главным образом углеводы, а азот высвобождается в виде аммиака.
Газообразные соединения азота могут поступить в почву из атмосферы с осадками и пылью. С атмосферными осадками в почву поступают аммиак и окислы азота, которые появляются в атмосфере при извержении вулканов, грозовых разрядах, а также как следствие промышленного загрязнения атмосферы. Однако этот источник дает очень небольшое количество N. Правда, вблизи промышленных городов такая прибавка за год может составлять до 20 кг N на 1 га, но обычно она бывает не более 2—10.кг, что, возможно, имеет некоторое значение для сфагновых болот, крайне бедных доступным азотом.
Важнейшим способом пополнения почвы азотом является процесс его биологической фиксации из атмосферы, связанный с деятельностью микроорганизмов (прокариотов). По отмирании микроорганизмов азот возвращается в почву. Грибы (эукариоты), видимо, не способны к фиксации N. Биологическая фиксация N происходит с участием особого активирующего фермента — нитрогеназы, что дает ей возможность идти при обычных температурах и давлении; в искусственных же условиях фиксация N возможна лишь при очень высоком давлении и температуре в сотни градусов. Фиксация происходит двумя путями. Несимбиотическая фиксация. В этом случае N атмосферы включается в органические соединения свободно живущими микроорганизмами, которые используют для фиксации энергию, содержащуюся в органическом веществе почвы или в прижизненных выделениях авто-трофов, т. е. это сапро- или эккрисотрофы (Работиов, 1979). Упомянем важнейшие роды сапротрофных азот-фиксаторов: Azotobacier, Aerobacter, Beijerinckia, Mycobacterium, Methylobacieria, Apirillum, Bacillus, Entero-bacter, Klebsiella, Rhodopseudomonas, Pseudomonas, Clostridium, Mechanobacterium и др. Особое значение среди азотфиксаторов.имеют свободноживущие и эпифитные синезеленые водоросли (циааюбакте-рии). Азотобактер — строго аэробная бактерия культурных плодородных почв с зернистой структурой; в бесструктурных, холодных, кислых или уплотненных почвах он отсутствует. Азотобактер обычно связан с ризосферой определенного культивируемого растения: его довольно много в ризосфере клевера, люцерны, риса, но на тех же почвах его значительно меньше в ризосфере пшеницы, льна, хлопчатника. Видимо, это объясняется или отсутствием необходимых питательных веществ в ризосфере этих растений, или наличием каких-то бактерий-антагонистов. Азотобактер — основной фиксатор азота в тех сообществах, где почти нет бобовых (например в злаковниках).
Клостридий способен фиксировать N в основном в анаэробных условиях, иногда и в аэробных, но в присутствии некоторых других бактерий; этот организм широко распространен во всех почвах земного шара, но лучше развивается в почвах, близких к нейтральным, не ниже рН 5. Некоторые азотфиксаторы тропиков (Beijerinckia) могут фиксировать азот при рН 3, и существуют они как в почве, так и на листьях (в филло-сфере).
Синезеленые водоросли (Anabaena, Nostoc) замечательны тем, что они не только С-автотрофны, но и N-автотрофны; встречаются в водоемах, в почве, на листьях и хвое, в горячих источниках и, что особенно важно, на безжизненных горных породах, являясь пионерами их заселения. Фиксация N синезелеными водорослями может идти и в экстремальных условиях (пустыни, скалы), а это имело огромное значение на первых •порах появления растений на суше вообще. Есть взгляды, что помимо самостоятельной способности фиксировать N воздуха почвенные водоросли помогают азотобактеру, который в их присутствии энергичнее фиксирует N. По мере формирования растительного покрова роль синезеленых водорослей снижается и основное значение в фиксации азота приобретают те организмы, которые способны получать энергию от зеленых растений — автотрофов, т. е. организмы ризосферы, подстилки. Поскольку несимбиотическая фиксация азота атмосферы начинается с восстановительного расщепления молекулы N2, то она требует большого количества энергии, а поэтому обычно лимитируется недостатком питательных веществ для самих азотфиксаторов. В связи с этим несимбиотическая фиксация идет довольно плохо на почвах с грубым гумусом, а в арктических холодных областях она сильно тормозится низкими температурами. Оптимальные условия для несимбиотйческих фиксаторов (особенно для синезеленых водярослей) представляют теплые и постоянно влажные районы. Общее же значение свободноживущих азотфиксаторов для растительности нормально дренированных почв невелико. Большинство свободноживущих азотфиксаторов дают незначительные количества N, однако в биогеоценозе в совокупности они могут обеспечивать поступление 5—10 кг/га азота в год.
Симбиотическая фиксация, т. е. превращение N атмосферы в органический микроорганизмами, в первую очередь живущими в симбиозе с растениями, особенно в клубеньках бобовых. Здесь обычно обнаруживаются облигатные аэробы, представители рода Rhizobium — несколько видов, включающих специализированные расы, специфические для определенных видов бобовых и живущие в почве тех областей, где распространено растение-хозяин; вне ареалов растений-хозяев они не встречаются. Внедряясь в корень, бактерии вызывают ненормальное разрастание его тканей, в (результате чего и образуется клубенек. Внутри этого клубенька создаются благоприятные условия (рН, ^парциальное давление кислорода и т. д.) для функционирования бактерий. Корневые клубеньки соединены с растением-хозяином паренхвдлными клетками, которые облегчают поступление N в растение. В процессе симбиотической фиксации N определенное значение имеют синезеленые водоросли, которые могут вступать в симбиоз с некоторыми печеночными мхами, водными папоротниками (Azolla) или же играть роль эпифитов даже на мхах в условиях Арктики. Иногда, входя в состав напочвенных и элифитных лишайников, синезеленые водоросли могут фиксировать N в различных количествах, в зависимости от вида лишайника и условий произрастания (Работнов, 1978). Помимо синезеленых водорослей большую роль в фиксации N играют актиномицеты, симбиотичеоки связанные с высшими растениями более 150 видов (из семейств Betula-ceae, Rhamnaceae, Elaeagnaceae, Myricaceae, Casuarina-ceae, Coriaceae, Rosaceae).
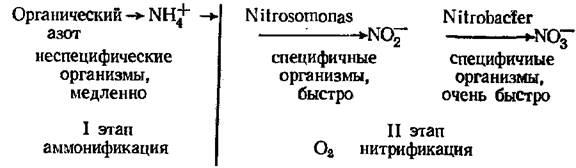
Поступление азота на поля за счет симбиотической фиксации во многом зависит от доли бобовых в общем посеве, их вида и урожайности. Наибольшую роль играют многолетние травы, особенно клевер и люцердоа. Однако нельзя забывать, что бобовые, фиксируя N цри помощи клубеньковых бактерий, одновременно могут получать его из почвы, а однолетние зерновые бобовые мало обогащают почву азотом, поскольку -почти весь фиксированный N удаляется с полей с урожаем зерна. Кроме того, интенсивная фиксация азота бобовыми настолько благоприятствует росту этих растений, что значительно усиливает их потребность в других элементах, особенно в К и Р. Фиксация N хорошо идет на почвах, богатых всеми минеральными соединениями; азотфик-саторы весьма нуждаются и в микроэлементах (особенно в Мо и Со). Для микроорганизмов, осуществляющих симбиотическую фиксацию, источником энергии являются клетки высших растений, а фиксация N требует много энергии: на 1 кг азота до 25 г углеводов. Вследствие этого сим биотическая фиксация значительно продуктивнее не симбиотической; выход N здесь может достигать 350 кг/га в год (Дельвич, 1972). Объем симбиотической фиксации, зависящей от поступления ассимилятов в клубеньки, связан с продуктивностью фитоценоза растения-хозяина: если повышается концентрация СО2 в воздухе (что стимулирует фотосинтез), то возрастает и фиксация N. Огромное значение в обеспечении растений минеральными формами азота имеет деятельность почвенных микроорганизмов, минерализующих органические вещества почвы, т. е. переводящих наиболее лабильные азотсодержащие вещества гумуса в минеральные соединения азота, потребляемые растениями. Схематически процесс минерализации можно свести к двум этапам: аммонификация, когда микроорганизмы почвы образуют из азотсодержащих веществ гумуса аммонийные соли, используя энергию окисления, а получающийся аммиак в почве немедленно соединяется с кислотами (угольной, органическими) и образует соли аммония; нитрификация — процесс образования из аммонийных солей, полученных на предыдущем этапе, усвояемых солей азотистой и азотной кислот, т. е. нитритов (N02-) и нитратов (N03-). Нитрификация происходит лишь при наличии аммония, степень обеспечения которым определяет ее интенсивность. В итоге весь процесс минерализации азота можно выразить следующей схемой;
(NO2-), но при этом выделяется достаточно энергии для существования Nitrosomonas. Nitrobacter извлекает из.нитритов ту энергию, которая осталась после деятельности Nitrosomonas, и этой энергии ему достаточно для существования. В природных условиях растения поглощают аммоний и нитраты в различном соотношении, что зависит от их содержания в почве и от других фактрров (Например конкуренции). Обычно считается, что чем беднее почва доступными формами азота, тем большее значение в питании растений имеет аммоний, и чем почва богаче доступным азотом, тем большее значение имеют нитраты. Последние не обнаруживаются в водных вытяжках кислых почв (лесов бореального типа, тундр, олиготрофных болот и др.). Таким образом, процесс аммонификации ведут многие неспециализированные организмы, идет он довольно медленно и в весьма широких пределах условий среды (рН почвы, температуры и т. д.). В случае плохо аэрируемых кислых почв (кустаркичковая тундра, болота, торфяники и т. д.) могут накапливаться продукты аммонификации. Аммонификация повышается с возрастанием температуры до 60—70°. А. Я. Орлов и д-р. (1974) отмечают, что содержание аммонифицирующих бактерий в лесной почве различно в разных, типах лесов. Например, для еловых лесов их больше всего найдено в подстилке ельника кислицевого, вдвое меньше в торфянистом слое хвощово-сфагнового типа и меньше всего в подстилке сосняка-черничника.
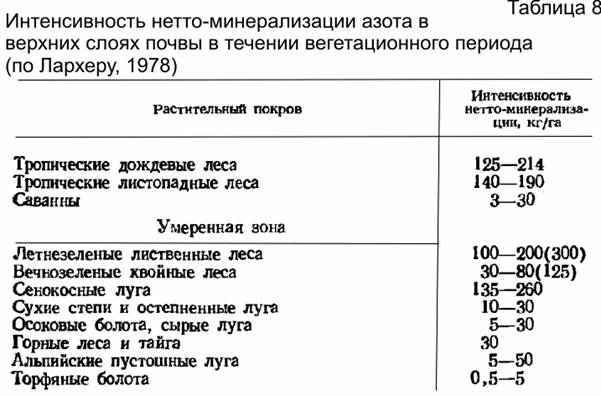
Нитрификация требует свободного кислорода, поэтому осуществляется, как правило, в самом верхнем слое почвы (в подстилке и в подстилающем ее горизонте), что во многом определяет и размещение корней растений. Обычно подчеркивается, что нитрификация идет на нейтральных или слабокислых почвах. Однако строгой положительной корреляции между нитрификацией и рН почвенного раствора не установлено, видимо, имеется некоторая корреляция с содержанием Р и особенно А1. Увеличение сухости почвы обычно снижает интенсивность образования нитратов непосредственно, а сильная влажность ингибирует этот процесс из-за снижения аэрации. Оптимальная температура нитрификации при- мерно 30°, а ниже 5 и выше 40° нитрификация резко снижается. Процесс минерализации N органических соединений почвы одновременно сопровождается процессами, в ходе которых часть нитратов и другие формы азота могут потребляться или вообще теряться для почвы. Такими процессами являются иммобилизация и денитрификация азота. Иммобилизация — превращение минерального азота в органическую форму, т.е. биологический процесс усвоения азота или самими микроорганизмами, или высшими растениями. В этом случае идет ассимиляция N, иначе говоря, фактическое изъятие его из абиотического компонента почвы. Денитрификация означает потерю азота почвой, удаление его в атмосферу. При этом из нитритов и нитратов образуются газообразный азот и его закись N20, а также газообразный аммиак. Следовательно, иммобилизация и денитрификация — это разные процессы. Нитраты частично потребляются самими микроорганизмами, но большую их часть потребляют растения. O2, и при этом образуется газообразный азот. Считают, что денитрификация не идет в аэробных условиях, однако она может происходить не только в условиях анаэро-биозиса и не обязательно в щелочной среде (Петербургский, 1979). К тому же имеется довольно много почв с анаэробными условиями, благоприятствующими денитрификации. В арктических тундрах, болотах и в других местах с ограниченным поступлением кислорода декитрификация идет и вблизи поверхности почвы. Косвенные расчеты нетто-минерализации (т. е. избытка минерального азота, остающегося после удовлетворения потребностей микроорганизмов) показала, что она зависит от очень многих факторов: температуры, влажности почвы, ее рН, наличия /питательных веществ; меняется она и по сезонам, в зависимости от смены комплекса условий. Лархер (1978) приводит сводную таблицу нетто-минерализации в верхних слоях почвы за вегетационный период (табл. 8). Мы видим, что в хвойных лесах минерализация идет почти вдвое медленнее, чем в лиственных.
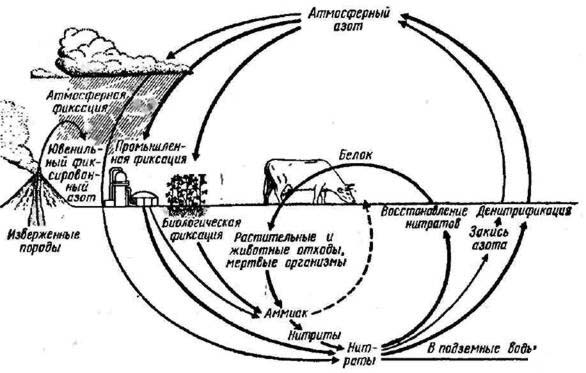
NO3- аммоний, нитраты. Но как раз эти формы, воспринимаемые растением, подвержены значительным колебаниям. Схематический цикл азота в целом представлен на рис. 64.
Рис. 64. Схема круговорота азота (по Дельвич, 1972) Надо, однако, помнить, что для растений важно как общее содержание N в данный момент, так и интенсивность его пополнения, оборота. В лесу оборот азота довольно высок, например насаждения сосны в течение года поглощают его столько же, сколько и сельскохозяйственные культуры, хотя потребность сосны в других питательных веществах не очень велика. В подстилке сосновых лесов содержание азота обычно колеблется в пределах 0, 5—2, 0% (Орлов и др., 1974). Для хвои сосны отмечены следующие показатели содержания азота: в сосняке кислицево-черничном — 1, 6%, на бедных песчаных почвах — 1, 3%, на сильно заболоченных почвах содержание азота падает до 1, 0—1, 1%. В хвое ели концентрация составляет в ельнике нелицевом — 1, 31% (что близко к оптимальному снабжению), в ельнике черничном — 1, 1%, в ельнике хвощово-сфатно-вом — 1, 2% (что указывает на крайний недостаток азота). Содержание N в хвое может сильно колебаться по годам в зависимости от погодных условий. Надо учесть, что стволы деревьев (древесина) накопляют около 13% азота. Установлена довольно четкая корреляция между бонитетом сосновых и еловых насаждений и количеством N, которое почва может дать лесу. Обеспеченность азотом обусловливает также и темпы разложения опада в лесу. Способность связывать N воздуха при помощи азотфиксирующих микроорганизмов имеет особенно большое значение для растений-пионеров, например видов ольхи, поселяющихся на бедных азотом почвах; она обогащает эти почвы азотом.
Для оценки азотного режима шлей, как и для оценки кислотности почв, Элленберг (предложил выделять группу видов по степени их нитрофильности. К группе видов, растущих только на почвах с очень хорошим обеспечением азотом, относятся также виды, произрастающие около навозных куч и на почвах, загрязненных органическими отбросами. Это так называемые рудеральные сорняки — особо витрофильные виды. Сюда входят хорошо известные синантропные виды — Urtiса dioica, Lamium album, виды рода Arctium. Поскольку в таких местах идет усиленная нитрификация, то эти растения воспринимают нитраты в таких количествах, что их можно обнаружить даже в клеточном соке. Отношение к потреблению N растениями, например лугов, может определять конкурентные взаимоотношения. При равном количестве доступного азота в надземных органах луговых злаков содержится его значительно меньше, чем у видов разнотравья (Работнов, 1979). Иначе говоря, используя одно и то же количество N, злаки развивают более мощную систему надземных органов, обеспечивая себе более высокую конкурентоспособность. Потребление N луговыми видами зависит от многих факторов (Работнов, 1974), но особенно велика роль конкуренции. Так, некоторые виды, обычно встречающиеся на лугах на почвах, бедных азотом (например душистый колосок), в опытах в сосудах, без конкуренции, довольно хорошо реагируют на азот (на его высокую обеспеченность). Кроме того, встает вопрос, совершенно ли необходимо для оптимального развития нитрофильных видов высокое содержание доступного N в почве (как обычно говорят) или они растут на таких почвах потому, что лучше переносят избыток N, отчего более конкурентоспособны в подобных местообитаниях. Опыты показали следующее: облигатные нитрофильные рудеральные виды предъявляют высокие требования к снабжению азотом и переносят гораздо большие количества нитратов, чем факультативные нитрофильные сорняки. Правда, последние при меньших количествах N (развиваются хуже. Интересно, что вне конкуренции крапива не предъявляет особых требований к нитратам. В естественных же условиях ее типичными местообитаниями являются влажные, богатые гумусом почвы с высокой интенсивностью нитрификации. Поэтому возможно, что крапива реагирует не столько на имеющиеся количества нитратов, сколько на постоянное их пополнение, приток. Хорошо нитрифицированные почвы под зарослями крапивы содержат к тому же довольно большие количества фосфора, от которого также во многом зависит ее рост. Оптимальное развитие нитрофильных видов при хорошем снабжении азотом указывает на то, что в конкурентной борьбе на почвах, богатых- азотом, они побеждают, а на почвах, бедных азотом, оказываются побежденными. Сильно нитрофильные виды можно обнаружить в разных местообитаниях, например на скалах и побережьях, где имеются птичьи базары и поверхность сильно удобрена. Кроме того, нитрофилы охотно -поселяются на местах стоянок скота и скоплений диких животных, на бывших стоянках человека (юрты), по берегам водоемов, где имеется вынос органической массы водорослей, и т. д. Весьма богаты нитрофилами лесные вырубки, пожарища и просеки (вспомним заросли крапивы и др.). Однако в этом случае действует фактор выключения корневой системы деревьев, усиление процесса нитрификации, чему способствует на вырубках большая инсоляция, увеличение нагрева почвы. В таких условиях накопление нитратов способствует разрастанию нитрофильных видов, типичных для вырубок: Atгора belladonna, Rubus idaeus, Sambucus racemosa и др. В буковых лесах на высокую нитрификацию указывает Anemone nemorosa; еще интенсивнее нитрификация идет в буковых и ольховых лесах с Mercurialis perennis. Число и распространение нитрофильных сообществ и их видовое богатство обычно увеличиваются с усилением аридности климата. В гумидных областях эти сообщества обеднены из-за значительного выщелачивания почвы и привязаны к более сухим местообитаниям. В аридных областях нитрофильные сообщества распространены шире, они более богаты видами. Таким образом, разные виды используют различные источники азотного снабжения. Виды кислых почв проявляют большую способность воспринимать аммоний, чем растения менее кислых почв, хотя многие из них неплохо растут при снабжении нитратами. Например, довольно ацидофильный вид Deschampsia flexuosa лучше растет на почвах, богатых аммонийными солями. Rumex acetosa хорошо растет при достаточном количестве нитратов (при менее кислом режиме), но его рост сильно ингибируется на почвах с преобладанием аммония и особенно при низких значениях рН. Виды, адаптированные к почвам, бедным азотом, способны расти на них за счет подавления в конкурентной борьбе менее приспособленных видов. И наоборот, распространение некоторых видов ограничено участками с хорошим азотным питанием именно потому, что они не способны существовать на почвах, бедных азотом. При определении отношения отдельных видов к доступному N почвы необходимо учитывать комплекс многих факторов. Еще больше усложняется дело тем, что недостаток азота обычно вызывает.некоторые морфологические изменения, имеющие сходство с KcepOMqp-фозом. Однако здесь речь идет о пейноморфозе, т. е. о морфологических изменениях, связанных с недостатком какого-то (в данном случае N) питательного элемента, так же, как это наблюдалось на кислых и холодных почвах. Недостаток N вызывает отдельные признаки ксероморфоза листьев у некоторых растений, особенно верховых болот (Lundkvist, 1956; и др.). Он может даже вызвать суккулентность лри неизменном снабжении водой, -поскольку нехватка N повышает восприятие солей. Но, как мы уже упоминали (глава о воде), у некоторых мезофитов (Chenopodium album, Urtica dioica) при снижении азотного питания усиливаются мезоморфные признаки, а при сухости почв (вместе с недостатком N) у крапивы увеличивается поверхность листьев и даже могут появиться признаки суккулентности. Интересно, что на сухих дюнах при довольно большом недостатке воды виды бобовых, имеющие азотфиксирующий симбиотрофный аппарат, не проявляют признаков ксероморфоза (Mothes, 1932). Недостаток N подавляет фотосинтез сильнее, чем недостаток других элементов; плохое обеспечение растения азотом обусловливает снижение нетто-ассимиляции единицы листовой поверхности и уменьшение -самой поверхности листа. В прямой зависимости от N находится и содержание хлорофилла в листьях (Лир и др., 1974); имеются сведения, что недостаток N снижает зимостойкость растений. Признаками недостатка азота можно считать мелколистность, тонкие стебли, уменьшение ветвистости. Листья на первых этапах голодания имеют желто-зеленую окраску, затем желтеют, краснеют или приобретают иную окраску; корни утончаются. С повышением обеспеченности растений азотом наблюдается снижение в них углеводов. Например, у сахарной свеклы максимальное содержание сахара получают при меньших количествах N, чем это необходимо для наивысшего урожая растения в целом. Лучшее обеспечение азотом увеличивает рост надземных частей в большей степени, чем подземных. Улучшение снабжения азотом вызывает определенные.изменения в морфологии, анатомии и химическом составе растений. Так, у травянистых растений увеличиваются высота и число надземных побегов, количество листьев, их размеры, возрастают суккулентность (увеличивается -количество воды), количество хлорофилла (с чем связано усиление теневыносливости); увеличивается отношение массы надземных частей к подземным. При достаточном обеспечении азотом растения обычно начинают вегетацию при более низких температурах и позднее заканчивают ее осенью. Длительное использование усвоенного N увеличивает продолжительность жизни листьев; вечнозеленость можно рассматривать как приспособление к произрастанию на бедных почвах. Кроме того, лучшая обеспеченность N ведет к более экономному расходованию воды на образование фитамассы (снижается транспирационный коэффициент). Хорошо обеспеченные азотом растения могут быть бесплодными, но развивать большую вегетативную массу. В случае некоторого уменьшения N идет сокращение вегетативной массы, увеличение плодоношения, а при недостатке N сокращаются и вегетативный рост и плодоношение. Избыток азота усиливает полегаемость, поскольку увеличивается масса надземных частей и уменьшается прочность нижних частей стебля. При изучении растений естественных сообществ иногда пренебрегают нехваткой доступного N, хотя его влияние на культурные растения изучено довольно широко. Признаки азотного голодания обнаруживают, например, сеянцы ели под пологом леса (Орлов, Кошельков, 1971), поскольку на их снабжение азотом сильно влияет конкуренция корней взрослых деревьев (Карпов, 1973). 13. Экологическое значение элементов минерального питания. Макро- и микроэле-менты, их значение. Олиго-, мезо- и эутрофные виды. Минеральное питание растений
Нормальный рост и развитие растений происходят в том случае, когда все макро- и микроэлементы в почве имеются в необходимом для растений количестве. Если содержание некоторых элементов чрезмерно высоко или низко, на листьях появляются признаки дефицита или токсикоза данного элемента.
Основные элементы, которые необходимы для роста и развития: углерод, водород, кислород, азот, фосфор, сера, калий, магний и железо, а также микроэлементы: бор, цинк, медь, марганец и др. Кислород, водород и углерод растения поглощают из воды и воздуха, а остальные элементы из почвенного раствора. Каждый из этих элементов играет определенную роль в жизнедеятельности растений.
Очень важно соблюдать нормы подкормок и не вносить удобрения по принципу " чем больше - тем лучше" или " на глазок". Клюква относится к тем растениям, которые лучше недокормить, чем перекормить. Лучше всего использовать сернокислые формы удобрений, так как они позволяют поддерживать кислую реакцию почвенной среды.
*АЗОТ. Азот имеет важное значение для роста. Он входит в состав аминокислот, белков и других веществ. Первый признак дефицита азота - покраснение и измельчание листьев; при сильном дефиците - цвет листьев становится желто-зеленым и над основной окраской образуется красно-бронзовый налет. Сначала признаки азотного дефицита появляются на старых листьях стелющихся побегов и только потом на листьях вертикальных побегов. При сильном азотном дефиците рост побегов очень слабый или прекращается совсем; развитие ягод также задерживается. При повышенном содержании азота в почве наблюдается сильный рост побегов.
*ФОСФОР. Этот элемент также имеет очень важное значение в жизнедеятельности растений. Он входит в состав нуклеиновых кислот, которые в соединении с белками образуют нуклеопротеиды - важную составную часть клеточного ядра и протоплазмы, а также в состав целого ряда других соединений. Фосфор концентрируется в молодых тканях растений, цветках, семенах. При фосфорном дефиците деление клеток замедляется, и рост растений прекращается, старые листья приобретают красный цвет, замедляется рост корней, а также образование боковых корней. Первым признаком дефицита фосфора является покраснение листьев, как на стелющихся, так и на прямостоячих побегах. При более сильном дефиците все листья приобретают пурпурную окраску, рост побегов прекращается, и через некоторое время они засыхают.
*КАЛИЙ. Из всех необходимых катионов калий нужен в наибольших количествах. В ионной форме он выполняет важную для жизнедеятельности растений функцию: этот элемент является компонентом ионных " насосов", ионных потоков и потенциалом действия - процесса единого ионного транспорта. Достаточное обеспечение калием способствует повышению морозо- и засухоустойчивости, биосинтезу и транспорту углеводов, в результате чего возрастает накопление углеводов в вегетативных и запасающих органах, т.е. в семенах и ягодах. При оптимальном калийном питании ягоды клюквы значительно крупнее и интенсивнее окрашены. При недостатке калия рост замедляется, а при сильном дефиците листья приобретают бронзово-красный цвет. В течение вегетационного периода листья разрушаются, окраска кончиков молодых листьев изменяется, как и при азотном дефиците. Стебли становятся тонкими и ломкими.
КАЛЬЦИЙ. Этот элемент присутствует во всех клеточных структурах организма и стабилизирует их функции. Особенно важное значение кальций имеет для нормального роста и деятельности корневой системы. При недостатке этого элемента задерживается рост корней и формирование корневой системы. |
Последнее изменение этой страницы: 2017-05-04; Просмотров: 99; Нарушение авторского права страницы