
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
УЧЕБНО-МЕТОДИЧЕСКИЙ КОМПЛЕКС ПО ДИСЦИПЛИНЕСтр 1 из 49Следующая ⇒
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ Пензенский государственный педагогический университет имени В.Г. Белинского
Кафедра анатомии, физиологии и гигиены человека УЧЕБНО-МЕТОДИЧЕСКИЙ КОМПЛЕКС ПО ДИСЦИПЛИНЕ Генетика Химия Биология Пенза - 2007 г. Тексты лекций
Тема 1. ГЕНЕТИКА – НАУКА О НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ. ЗАСЛУГИ Г. МЕНДЕЛЯ Задачи: 1. Познакомить с историей возникновения генетики как науки, заслугами Г.Менделя, его гибридологическим методом исследования, с основными генетическими понятиями и терминами. 2. Научить правильно раскрывать сущность основных понятий генетики. 3. Сформулировать убежденность в том, что знание основных понятий необходимо для понимания важных биологических закономерностей. Общие закономерности наследования признаков Для всех эукариот характерны общие закономерности наследования признаков. Эти закономерности сформулированы Г. Менделем. Главный постулат Г. Менделя, который он доказал в своих известных экспериментах на горохе, состоит в том, что каждый признак определяется парой наследственных задатков, которые позже были названы аллельными генами. С развитием хромосомной теории наследственности было показано, что аллельные гены находятся в гомологичных хромосомах в одинаковых локусах и кодируют один и тот же признак. Пара аллельных генов может быть одинакова (АА или аа), тогда говорят, что особь гомозиготна по данному признаку. Если пара аллельных генов альтернативна (Аа), то особь гетерозиготна по данному признаку. Совокупность генов данного организма называется генотипом. Часто под генотипом понимают одну или несколько пар аллельных генов, которые отвечают за один и тот же признак. Совокупность признаков данного организма называют фенотипом. Фенотип развивается в результате взаимодействия генотипа с внешней средой. Г. Мендель ввел понятия доминантных и рецессивных генов. Аллель, который определяет фенотип гетерозиготы, он назвал доминантным. Например, ген А в гетерозиготе Аа. Другой аллель, который не проявляется в гетерозиготном состоянии, он назвал рецессивным. В нашем примере это ген а. Основные закономерности наследования признаков по Менделю (закон единообразия гибридов первого поколения, закон расщепления на фенотипические классы гибридов второго поколения и закон независимого комбинирования генов в ди- и полигибридных скрещиваниях) реализуются благодаря существованию закона чистоты гамет. Суть этого закона состоит в том, что пара аллельных генов, определяющая тот или иной признак, никогда не смешивается, и в процессе образования гамет расходится в разные гаметы, т. е. в каждую гамету попадает один ген из аллельной пары. Цитологически это объясняется мейозом. Генетика человека опирается на общие принципы, полученные первоначально в исследованиях на растениях и животных. У человека также имеются менделирующие признаки, которые наследуются по законам, установленным Г. Менделем. Г. Менделем разработан и основной метод генетики — гибридологический. Он основан на скрещивании особей одного вида с альтернативными признаками и количественном анализе особей в полученных фенотипических классах. В настоящее время гибридологический анализ является частью генетического анализа. Гибридологический метод в генетике человека не применяется. Т ема 6. ГЕНЕТИКА ПОЛА Задачи :
Хромосомный механизм, определяющий пол. В кариотипе человека 46 хромосом (23 пары), из них 22 пары хромосом одинаковы у мужчин и женщин (аутосомы), а по одной паре хромосом, называемой половой, мужчины и женщины отличаются. Половые хромосомы женщины — XX, половые хромосомы мужчины — XY (см. главу 6). Половые хромосомы представлены в каждой соматической клетке индивида. При образовании гамет во время мейоза гомологичные половые хромосомы расходятся в разные половые клетки. Следовательно, каждая яйцеклетка имеет 22 аутосомы и 1 половую хромосому Х (гаплоидный набор хромосом равен 23). Каждый сперматозоид также имеет гаплоидный набор хромосом (22 аутосомы и 1 половая). Половина сперматозоидов содержит Х-хромосому, другая половина — Y-хромосому. Поскольку у женщин половые хромосомы одинаковы и все яйцеклетки несут Х-хромосому, то женский пол у человека называют гомогаметным. У мужчин сперматозоиды различаются по наличию половых хромосом (X или Y), следовательно, мужской пол называют гетерогаметным. Пол человека определяется в момент оплодотворения. Женщина имеет один тип гамет (X) мужчина — два типа гамет (X и Y), причем они образуются в равной пропорции согласно законам мейоза. При оплодотворении хромосомные наборы гамет объединяются. В зиготе 22 пары аутосом и одна пара половых хромосом. Если яйцеклетка оплодотворена сперматозоидом с Х-хромосомой, то в зиготе пара половых хромосом XX. Из такой зиготы развивается девочка. Если яйцеклетка оплодотворена сперматозоидом с Y-xpo-мосомой, то набор половых хромосом в зиготе XY. Такая зигота даст начало мужскому организму. Таким образом, у человека пол будущего ребенка определяет мужчина, так как он гетерогаметен по половым хромосомам. Соотношение полов при рождении по данным статистики соответствует примерно 1:1. Наследование признаков, сцепленных с полом. Хромосомы Х и Y гомологичны, так как у них есть общие гомологичные участки, где локализованы аллельные гены. Однако эти хромосомы, несмотря на гомологию отдельных локусов, различаются по морфологии. Помимо общих участков Х- и Y-хромосомы имеют большой набор генов, по которым они различаются. В Х-хромосоме имеются гены, которые отсутствуют в Y-хромосоме, в свою очередь, в Y-хромосо-ме имеются гены, которых нет в Х-хромосоме. Таким образом, у мужчин в половых хромосомах имеются гены, которые не имеют второго аллеля в гомологичной хромосоме. В этом случае признак определяется не парой аллельных генов, как обычно менделирую-щий признак, а только одним аллелем. Такое состояние гена называется гемизиготным. Признаки, развитие которых обусловлено одним аллелем, расположенным в одной из альтернативных половых хромосом, называются сцепленными с полом. Они развиваются преимущественно у одного из двух полов. Эти признаки наследуются по-разному у лиц мужского и женского пола. Признаки, сцепленные с Х-хромосомой, могут быть и рецессивными, и доминантными. Рецессивные признаки — гемофилия, дальтонизм, атрофия зрительного нерва и миопатия Дюшена. Доминантные — рахит, не поддающийся лечению витамином D, и темная эмаль зубов. Рассмотрим сцепленное с Х-хромосомой наследование на примере рецессивного гена гемофилии. У мужчины половые хромосомы XY. Ген гемофилии локализован в Х-хромосоме и не имеет аллеля в Y-хромосоме, т. е. находится в гемизиготном состоянии. Следовательно, несмотря на то, что признак рецессивный, у мужчин он проявляется. У женщин половые хромосомы XX. Признак определяется парой аллельных генов, поэтому гемофилия проявляется только в гомозиготном состоянии: Основные формальные характеристики Х-сцепленного рецессивного наследования следующие. Обычно поражаются мужчины. Все их фенотипически здоровые дочери являются гетерозиготными носительницами, так как от отца в процессе оплодотворения получают X-хромосому:
Среди сыновей гетерозиготных матерей соотношение пораженных и непораженных 1:1.
Такое наследование получило название крисс-кросс (крест-накрест), т. е. сыновья наследуют фенотипический признак матери, а дочери — отца. Законы передачи признаков, сцепленных с Х-хромосомами, были впервые изучены Т. Морганом. Помимо Х-сцепленных признаков, у мужчин имеются Y-сцепленные признаки, называемые голандрическими. Гены, определяющие голандрические признаки, локализованы в тех районах Y-xpo-мосом, которые не имеют аналогичных участков в Х-хромосоме. Эти признаки также определяются только одним аллелем. Так как голандрические признаки определяются генами, находящимися только в Y-хромосоме, то выявляются они у мужчин и передаются от отца всем сыновьям. Голандрические признаки: волосатость ушей, перепонки между пальцами ног, ихтиоз (кожа имеет глубокую исчерченность и напоминает рыбью чешую). Гомологичные районы Х- и Y-хромосом содержат аллельные гены. Они с равной вероятностью встречаются у лиц мужского и женского пола. К числу таких признаков относятся общая цветовая слепота (отсутствие цветового зрения) и пигментная ксеродерма — заболевание, при котором под влиянием ультрафиолетовых лучей на открытых частях тела образуются опухоли. Оба признака являются рецессивными. Признаки, которые определяются аллель-ными генами, находящимися в Х- и Y-хромосомах, наследуются как классические менделирующие признаки. Наследование ограниченное и контролируемое полом. Признаки человека, наследование которых каким-то образом связано с полом, подразделяется на несколько категорий. Одна из категорий — признаки, ограниченные полом. Развитие этих признаков обусловлено генами, расположенными в аутосомах обоих полов, но проявляющимися только у одного пола. Например, гены, определяющие ширину таза женщины, локализованы в аутосомах, наследуются и от отца, и от матери, но проявляются только у женщин. То же касается возраста полового созревания женских индивидов. Среди мужских признаков, ограниченных полом, можно назвать количество и распределение волосяного покрова на теле. К иной категории относятся признаки, контролируемые полом или зависимые от пола. Развитие этих соматических признаков обусловлено генами, расположенными в аутосомах. Проявляются ониу мужчин и женщин, но по-разному. Например, у мужчин раннее облысение — признак доминантный, он проявляется у доминантных гомозигот (АА) и у гетерозигот (Аа). У женщин этот признак рецессивный и проявляется только у рецессивных гомозигот (аа). Для признаков, контролируемых полом, выраженность обусловлена в большой степени половыми гормонами. Свойства гена Как единица функционирования ген имеет ряд свойств: · аллельное состояние (2, 3 и более альтернативных варианта). В зависимости от сочетания аллелей может наблюдаться гомозиготность или гетерозиготность; · дискретность – развитие разных признаков контролируется разными генами. Менделирующие признаки: 1 ген – 1 фен; · дозированность – ген обусловливает развитие признака до определенного количественного предела (дозы). Например: размер ушной раковины, длина спинки носа; · специфичность – определенный ген обусловливает развитие определенного (специфического) признака или их группы (плейотропное действие гена); · стабильность – при отсутствии мутации ген передается в ряду поколений в неизменном виде. Гены подразделяются на аллельные и неаллельные, в связи, с чем выделяют и типы их взаимодействия. Проект « Геном человека» Цель проекта: определение полной нуклеотидной последовательности молекул ДНК человека и идентификация всех генов. Исследования были начаты сотрудниками Института геномных исследований (США) в 1992г. На базе института в 1998г. была создана коммерческая компания «Селера джиномикс», в финансировании которой приняли участие частные фирмы. Именно эта компания первой объявила о завершении работ по расшифровке ДНК человека. Согласно результатам, геном человека составляет около 32тыс генов и вряд ли превышает 35тыс. Уровень точности можно считать удовлетворительным: 1 ошибка на 104 нуклеотидов для 80% генома и 1 ошибка на 10 6 нуклеотидов - для 20% генома. Лучше всего расшифрована нуклеотидная последовательность двух самых маленьких хромосом человека - 21-22-й. Интересным является тот факт, что коммерческие компании, по сравнению с научными коллективами, не обязаны предавать гласности все свои результаты. Заявление сделано, но сами последовательности в полном объеме в международный банк данных переданы не были. В настоящее время готовятся заявки на патенты, т.к. результаты принесут прибыль фармацевтическим, медицинским и сельскохозяйственным компаниям. Правительства США, Японии, Великобритании, Германии, Франции стали выделять на работы по проекту «Геном человека» миллиарды долларов. Россия в начале создания проекта участвовала на равных условиях, но затем фактически приостановила свои вклады, и в будущем будет в экономической зависимости от этих стран, хотя многие специалисты, получив образование на родине, мигрировали в США и Западную Европу.
Свойства генетического кода 1. Генетический код триплетен. Триплет (кодон) — последовательность 3-х нуклеотидов, кодирующая 1 аминокислоту. 2. Генетический код вырожден (причем неравномерно). Из 64 триплетов 3 кодона УУА, УАГ, УГА не кодируют аминокислот. Первоначально их назвали нонсенс-кодонами, а позже - терминирующими кодонами или стоп-сигналами, т. к. они останавливают биосинтез белка. Оказалось, что 1 аминокислота может кодироваться несколькими триплетами (2-мя, 3-мя, 4-мя). Исключение составляют: мепшонин (АУГ) и триптофан (УГГ) - они кодируются только одним триплетом. Если бы из 64 кодонов - 20 кодировали аминокислоты, а 44 - ничего не значили, т. е. 1 кодон = 1 аминокислоте, код считался бы невырожденным. Следует помнить: если триплет АУГ стоит в начале цепи ДНК, он не кодирует аминокислоту метионин, а выполняет функцию инициирования считывания информации. 3. Однозначность - каждому кодону свойственна только одна определенная аминокислота. АГГ – ТАЦ – ААА – ТЦГ 4. Генетический код не перекрываем - синтез идет непрерывно, начавшись на определенном кодоне, процесс трансляции идет без пропусков, без каких-либо вставок вплоть до стоп-сигнала. / Инициирующий триплет - триплет - триплет - триплет - стоп-сигнал /. Последовательность триплетов определяет рибосома. 5. Генетический код универсален - он един для всех организмов, обладающих разным уровнем организации (кошка, мышь, человек, слон, бабочка, растение). Информация в ядерных генах кодируется одинаково. Аминокислоты не имеют видовых различий. Главный вывод: Расшифровка генетического кода позволила: • Понять механизм синтеза белка • Связать между собой дефекты синтеза определенных белков человека и наследственные заболевания • Создала необходимые предпосылки для диагностики и лечения заболеваний. В последние десятилетия было доказано, что при считывании информации очень важное значение имеет положение 1-го кодона (триплета). Оно определяется так называемой рамкой считывания. При сдвиге рамки считывания (выпадении или добавлении нуклеотидов) при последующей сборке белка в нем будет нарушена последовательность аминокислот. Это называется мутацией со сдвигом рамки. • Ученые считают, что единство генетического кода подтверждает тот факт, что все современные клетки являются потомками некогда 1 примитивной линии клеток, разработавшей биосинтез белка. Последовательность триплетов м-РНК, определяющей порядок расположения аминокислот в молекуле белка, соответствует комплиментарным кодонам гена, локализованного в структуре ДНК. Это свойство названо коллинеарностью, что означает соответствие последовательности аминокислот белка кодонам, расположенным в гене. Комбинативная изменчивость Этот тип изменчивости возникает в результате комбинирования генов родителей. По этой причине у детей могут появляться новые признаки. Причины комбинативной изменчивости различные: мейотический кроссинговер, свободное сочетание хромосом и генов, случайное слияние гамет с различным набором генов при оплодотворении. Комбинативная изменчивость у людей может быть связана с миграционными процессами. В последнее время все чаще отмечаются браки между супругами, родившимися в различных регионах одной страны или разных стран, межнациональные браки. Это может приводить к оплодотворению гамет, различных по генному набору, и др. Комбинативная изменчивость делает организмы более пластичными; они лучше приспосабливаются к меняющимся условиям окружающей среды, способствуя выживаемости вида. Примером комбинативной изменьчивости у человека может служить развитие групп крови системы АВО. Если мать имеет группу крови III (BO), а отец гетерозиготен по группе крови II (AO), то у детей возможна IV (AB) или I (OO) группы крови, отсутствующие у их родителей. Мутационная изменчивость Мутационная изменчивость по характеру изменения генотипа делится на генные (точковые) мутации, хромосомные и геномные. Отдельно выделяют цитоплазматические мутации, причиной которых является изменчивость определенных органоидов цитоплазмы (пластид, плазмид, митохондрий и др.), содержащих ДНК или РНК. Генные мутации Генные мутации — это изменения, происходящие в пределах одного гена. В результате генных мутаций у человека возникает большое число болезней обмена веществ (галактоземия, фенилкетонурия, алкаптонурия и др.). В настоящее время их число превышает 3,5 тыс. Известно, что один и тот же ген может мутировать в несколько аллельных состояний, образуя серию множественных аллелей. По этому типу наследуются такие признаки, как группы крови системы АВО у человека, окраска шерсти у кроликов, наличие светлых пятен на листьях у клевера и др. Поскольку у диплоидных организмов в одной хромосоме не более двух аллелей одного гена, то наследование будет происходить в соответствии с законами Г. Менделя. Генные (точковые) мутации Ген представляет собой определенный участок ДНК. ДНК, в свою очередь, полимер, состоящий из мономеров (нуклеотидов 4 типов, отличающихся азотистым основанием). Азотистое основание может быть 4-х типов: А - аденин; Г - гуанин; Т - тимин; Ц - цитозин. Последовательность нуклеотидов - видоспецифична. Генные мутации не случайно назвали точковыми, так как связаны с изменением 1-го (реже нескольких нуклеотидов) в молекуле ДНК или РНК. Выделяют несколько групп генных мутаций: 1. Транзиции (один пурин ошибочно заменен другим пурином или один пиримидин ошибочно заменен другим пиримидином). Замена нуклеотидов не изменяет ориентацию молекулы ДНК в пределах пары. Возможно 4 типа ошибок. а) А – Т, б) Г – Ц, в) Т – А, г) Ц – Г. 2. Трансверсии (мутации, связанные с ошибочной заменой пурина на пиримидин). Возможно 8 типов ошибок, в общей сумме они составляют >20% спонтанных мутаций. Замена пуринового основания на пиримидиновое, происходит как спонтанно, так и под действием мутагенов. К числу пуринов относятся: А, Г; к числу пиримидинов: Т и Ц. 3. Мутации со сдвигом рамки считывания (фреймшифт). Сдвиг рамки считывания вправо или влево происходит за счет: вставки лишней пары нуклеотидов (инсерция), удвоения (дупликация); выпадения пары нуклиотидов (делеция). В результате меняется триплетность кода, а, следовательно, и последовательность аминокислот в молекуле синтезируемого белка. В результате сдвига рамки считывания может возникнуть сутуация, связанная с образованием мутантного нонсенс-кодона - УАА, УАГ, УГА – который является стоп-сигналом. Синтез белковой молекулы остановится в неположенном месте, она будет незаконченной, следовательно, нарушатся и основная функции белка. Такие мутации называются нонсенс–мутациями, они связаны с появлением кодонов-терминаторов. Чем ближе к 5 концу (к началу транскрипции), тем короче будет белковая молекула. Наибольшее повреждающее действие оказывают именно нонсенс-мутации. Мутации возникают либо в момент репарации (т.е. самовостановления; при внешнем повреждении), либо как побочный продукт нормального процесса клеточной физиологии, в момент репликации (удвоения ДНК) и рекомбинации. Такая концепция получила название «ошибки трех Р», т.е. репарации, репликации, рекомбинации. Ошибка может происходить спонтанно (когда мы переписываем текст с учебника в тетрадь – то часто допускаем орфографические ошибки – вместо о - а, вместо е - и), и индуцированно (т.е. под влиянием мутагенов внешней среды). Пример. У человека известна наследственная патология – серповидно-клеточная анемия. Формирование серповидной формы эритроцитов - результат точечной мутации. Сдвига рамки не произошло, но один из триплетов кодирует иную аминокислоту. Последствия роковые: эритроциты деформируются, приобретают серповидную форму и не способны нормально транспортировать газы. Не все точковые мутации имеют фенотипическое проявление. Существует группа «молчащих» мутаций, для них характерно образование синонимического кодона. Дело в том, что в связи с вырожденностью генетического кода, ошибочная замена нуклеотита в 3-ей позиции триплета не оказывает влияния на изменение аминокислоты по данному триплету. Следовательно, частота произошедших мутаций и фенотипически выявленных при этом мутантов может не совпасть. Пример: аминокислота валин кодируется 4-мя триплетами: ГУУ, ГУЦ, ГУА, ГУГ. Замена нуклеотида в третьей позиции не приведет к замене аминокислоты валин. Последовательность аминокислот в кодируемом белке не нарушится. Главный вывод: Значимость нуклеотидных мутаций внутри кодона неравнозначна – замена первого и второго нуклитидов всегда ведет к изменению (замене) аминокислоты. Миссенс–мутация предполагает полную или частичную потерю функциональной активности белка. 3-й нуклеотид в большинстве случаев является синонимичным. Среди наследственных заболеваний, вызванных генными мутациями, следует назвать следующие патологии: а) аутосомно-доминантного типа – арахнодактилия, нейрофиброматоз; б) аутосомно-рецессивного типа – амавротическая идиотия, альбинизм, галактоземия; в) сцепленные с полом: гемофилия, дальтонизм, некоторые формы аллергических реакций. В настоящее время общее число наследственных заболеваний составляет, по разным источникам, от 2,5-4 до 7-9 тыс. Они вызывают аномалии обмена в-в, нарушения конституции, психические заболевания. Причина – генные мутации, хромосомные аберрации. Генные мутации нередко приводят к явлению, названному множественным аллелизмом. В этом случае, в популяции ген может быть представлен 3-мя или даже серией альтернативных аллелей. Генные мутации вызывают изменение признака в разных направлениях: морфологических, биохимических, физиологических. Причем, изменения эти могут быть сильно и слабо выражены. По функциональному значению генные мутации делятся на 3 класса:
Известный генетик Ф.Г. Добжанский в свое время писал: «Если мы сохраняем слабых и генетически больных детей и даем им с возрастом возможность продолжения рода, мы можем опасаться заката генетического. Но если мы дадим им умереть или страдать, в то время как можем помочь, мы, несомненно, предвидим закат моральный. По мнению генетиков в XXI веке метод генотерапии станет общедоступным. Межхромосомные перестройки Межхромосомные перестройки — транслокации — характеризуются перемещением участка одной хромосомы на другую (негомологичную) хромосому. Различают реципрокные и нереципрокные транслокации (транспозиции). В первом случае происходит взаимный обмен участками негомологичных хромосом, во втором — участок хромосомы изменяет свое положение или включается в другую хромосому без взаимного обмена. Робертсоновские транслокации образуются при слиянии двух центромер акроцентрических хромосом. В результате возникает одна мета- или субметацентрическая хромосома и число хромосом в клетке уменьшается на одну. 1) Транслокации – перемещение части 1 хромосомы на другую, не гомологичную (те не парную) ей. В результате группы сцепления генов нарушаются (зачастую приводя к летальному исходу). Выделяют 2 типа транслокаций: а) реципрокные – взаимный обмен участками негомологичных хромосом (меняется характер сцепления генов, коньюгация, образуется фигура креста). б) нереципрокные – участок 1 хромосомы включается в другую, без взаимного обмена.
К эволюционным преобразованиям приводят дицентрические слияния, когда 2 и более фрагмента негомологичных хромосом, несущих участки с центромерами, соединяются в единую структуру (центромеры при этом объединяются). Наиболее показательными, в этом плане, являются центрические (робертсоновские) – слияние двух центромер негомологичных акроцентрических хромосом с образованием 1 мета- или субметацентрической хромосомы.
У человека 2n = 46 хромосом. У шимпанзе 2n = 48 хромосом. Как оказалось, вторая хромосома человека содержит большую часть материала, гомологичного дополнительной паре хромосом шимпанзе. Помимо слияний, возможно и центрическое разделение – 1 хромосома делится на 2, с образованием новой центромеры (участок хромосомы без центромеры утрачивается, а с центромерой может реплицироваться). 2. Транспозиции - перемещение небольших участков генетического материала между разными хромосомами или в пределах 1 и той же при участии особыхмигрирующих генетических элементов – (транспозонов, IS–элементов). Хромосомные аберрации представляют собой разные варианты перемещений генов, нередко приводящих к изменению и самой структуры хромосомы. Геномные мутации Геномные мутации — полиплоидия и гетероплоидия — обусловлены изменением числа хромосом в кариотипе. - полиплоидные (полиплоидия от греч. poli – много, ploidia – одиночный, вид); - анеуплодные. Полиполоидия – изменение числа хромосом, кратное гаплоидному набору. Полиплоидные формы можно получать искусственно: действием колхицина, температурного шока, наркотических веществ и др.). У человека 1n = 23 хромосомы – гаплойдный набор, 2n = 46 хромосом – диплойдный, 3n = 69 хромосом – триплоидия, 4n = 92 хромосомы – тетраплоидия. Среди животных организмов полипоидия встречается крайне редко. При половом размножении процессы мейоза нарушены. Вместе с тем, активно синтезирующие клетки нередко содержат дополнительный набор хромосом (в клетках печени человека нередко насчитывается 69, 92 хромосомы и это нормально). Среди процессов, ведущих к полиплоидному состоянию клетки, следует назвать эндомитоз – удвоение хромосом с последующим делением центромер, но без расхождения по разным клеткам. У человека эндомитоз часто наблюдается в соматических тканях после воздействия мутагенов. Примечательно, что в эмбриогенезе человека такая аномалия хромосомного набора, как триплоидия (3n), встречается с частотой около 2% от всех хромосомных нарушений. Триплойдные зародыши погибают в начале 2-го месяца внутриутробного развития, а до возраста 6 – 7 месяцев (эмбриогенез) доживают не более 1 %. Синдром триплоидии (69, XXY) у новорожденных вперые был обнаружен в 60-х годах ХХ столетия. К настоящему времени списано свыше 60 случаев триплоидии у детей, максимальная продолжительность жизни которых составила 7 дней. Она сопровождалась многочисленными пороками развития: пороки головного мозга, сердца, желудочно-кишечного тракта и др. органов. Тетраплоидия (4n) встречается еще реже – из всех зародышей с хромосомными аномалиями лишь у 5–6 % . Для них характерны серьезные проки развития, зародыш редко вступает в плодный период, погибая на 2-м месяце. Всего описано 5 случаев рождения детей с тетраплоидией, которые вскоре погибли. Помимо полиплоидии, у человека возможен мозаицизм — наличие генетически разнородных клеток в организме. Это связано с нарушениями в период соматического развития. В результате внутри одного фенотипа появляются участки нового фенотипа. Известно, что интерсексьг часто содержат клетки с разным набором половых хромосом, например, 45,ХО; 46,ХХ, тогда как гермафродиты могут иметь 46,ХХ; 46,XY. Причины их образования различны: оплодотворение разными спермиями, слияние двух оплодотворенных яйцеклеток, ошибки в период первого дробления и др. Растения могут размножаться вегетативно, поэтому среду них полиплоидия представлена достаточно широко. Следует помнить, что растительные полиплоиды с нечетным набором хромосом 3n, 5n, 7n, - стерильны, тогда как 4n, 6n, 8n вполне плодовиты (сбалансированные). Большенство с/х растений относятся к разряду полиплоидов: крупная листовая пластинка, крупные цветки, крупный плод, повышенная сахаристость, повышенное содержание витаминов. Полиплоидные формы очень широко представлены в северных широтах, высоко в горах – быстрая вегетация (в связи с коротким летом), на фоне ограниченной плодовитости. В селекции используются полиплоиды двух видов: авто- и аллополиплоиды. Если мы имеем дело с кратным увеличением хромосом 1-го вида – это автополиплоиды. Организм, содержащий хромосомы разных видов называется аллополиплоидом (капустно-редичный гибрид, полученный при отдаленной гибридизации). Наряду с полиплоидами существуют моноплоидные организмы (трутни пчел) – развиваются из неоплодотворенного яйца. В природе есть виды с чередованием гапло- и диплофазы. Анеуплоидия (гетероплоидия) – изменение числа хромосом, некратное гаплоидному набору. Хромосома той или иной пары может полностью отсутствовать или, наоборот, быть повтореной трижды, четырежды и более раз: 2n–2 - «нуллисомик» - нет пары гомологичных хромосом. 2n–1 - «моносомик» - нет одной хромосомы в паре. 2n+1 - «трисомик» - в паре 1 лишняя хромосома. 2n+2 – в паре 2 лишние (дополнительные хромосомы). Анеуплоидия чаще всего возникает в результате неправильного расхождения хромосом в мейозе (анафаза–I, анафаза–II). В результате возникают аномальные по количеству хромосом гаметы, а после оплодотворения – гетероплоидные зиготы. Плазмиды В самых первых лекциях говорилось о том, что генетики научились внедрять в бактерии гены, контролирующие образование функционально активных белков человека. Вся процедура именуется клонированием генов или амплифицированием. Белки, которые раньше получали в ничтожно малом количестве, теперь нарабатывают в достаточно большом. В основе этой технологии лежит необычный принцип: Бактерии (E.coli), помимо своей собственной кольцевой хромосомы, часто содержат дополнительные маленькие кольцевые молекулы двухцепочечной ДНК, называемые плазмидами. Мутации, связанные с плазмидами позволяют бактериям перейти на новый вид питания. Плазмиды реплицируются автономно, и сами могут содержать гены, определяющие устойчивость бактерий к антибиотикам или/и гены, контролирующие синтез веществ, убивающих другие бактерии (например, колицины). Плазмидную ДНК можно выделить, затем подходящей рестриктазой расщепить в 1-ом сайте, превратив тем самым кольцевую молекулу в линейную с липкими концами. Фрагменты любой чужеродной ДНК с липкими концами, уже при помощи лигаз можно «сшить» с плазмидной ДНК в общее кольцо. Рекомбинантную конструкцию вводят в бактерию, где она реплицируется. Откуда взят фрагмент экзогенной ДНК – значения не имеет. Можно вшивать даже искусственно синтезированные гены. Если мы внедрили плазмиду с фактором устойчивости к антибиотику, то E.coli будет давать колонию на субстрате с антибиотиком. Нетрансформированные E.coli при этом расти не будут. Организм человека является средой обитания для сотен видов бактерий и вирусов. С биологической точки зрения организм человека представляет собой целую систему сосуществующих организмов-симбионтов. Далеко не все из симбионтов патогенные. Без некоторых видов бактерий человек просто не способен существовать, их утрата или снижение количества является причиной развития ряда тяжелых заболеваний. Расшифровка геномов многих болезнетворных микроорганизмов с идентификацией всех белков поможет разработать методы предупреждения и лечения инфекционных болезней. Оказалось, что геномы многих паразитирующих микроорганизмов обладают огромной пластичностью, имеются значительные структурные различия между штаммами бактерий, обитающих в разных частях одного организма. Для развития инфекционного процесса большое значение имеет генетический статус самого хозяина. Например, отдельные индивидуумы являются носителями вируса иммунодефицита, но СПИДом не болеют. У этих лиц имеется мутация в гене, кодирующем поверхностный белок, ответственный за попадание вируса внутрь лимфоидных клеток. Плотность белка на поверхности клеток снижена, вирус удерживается, но внутрь не попадает. Частота гомозигот по этой мутации среди жителей Европы составляет около 1%, они имеют выраженную устойчивость к ВИЧ-инфекциям. Более устойчивыми оказываются и гетерозиготные носители мутации, в российской популяции их частота достигает 13%. Общий вывод: митохондрии животных и растений, пластиды растительных клеток, плазмиды бактерий содержат собственную автономную ДНК, которая способна реплицироваться, транскрибировать рРНК, тРНК с последующим их участием в белковом синтезе. Органоиды цитоплазмы (хлоропласты, митохондрии), обладающие собственной автономной ДНК, образуют совокупность наследственных задатков, именуемую плазмоном. Кроме ядерной наследственности существует и цитоплазматическая.
Типы деления клеток Митоз - тип деления клеток, при котором из одной диплоидной клетки (2п=46) образуются две диплоидные, генетически равнозначные клетки. Процесс митоза в соматических клетках человека идет стандартно. К концу профазы хромосомы становятся отчетливо видимыми, каждая состоит из двух хроматид. Обе сестринские хроматиды прилежат одна к другой. Центромера в каждой хромосоме удерживает две сестринские хроматиды вместе. Ядрышко исчезает, ядерная оболочка распадается на фрагменты. Хромосомы располагаются в цитоплазме в центральной части клетки, оттесняя все органоиды к периферии. Во время метафазы центромеры всех хромосом располагаются в экваториальной плоскости между двумя полюсами. Хроматиды каждой хромосомы начинают отделяться одна от другой, оставаясь соединенными только в центромерной области. В районе центро-мер с противоположных сторон прикреплены нити веретена деления. Их количество может достигать нескольких десятков в районе каждой центромеры. Анафаза начинается с одновременного разделения всех центромер и расхождения сестринских хроматид каждой хромосомы к противоположным полюсам. Утрата синхронности процесса может привести к неправильному расхождению хромосом. Центромеры с помощью нитей веретена деления увлекают за собой дочерние хроматиды к противоположным полюсам. Анафаза заканчивается с прекращением движения хроматид, которые становятся хромосомами. У каждого полюса клетки должно оказаться по 46 хромосом, состоящих из одной хроматиды. Телофаза связана с образованием ядерных оболочек вокруг хромосом на двух полюсах клетки и началом перехода хромосом в состояние хроматина. Завершается телофаза образованием перетяжки в центральной части делящейся клетки, которая завершает деление клетки надвое. Известно, что у эукариот прохождение митоза может быть заблокировано физиологически или экспериментально, что приводит к развитию полиплоидных клеток. Системная полиплоидия не характерна для человека. Полиплоидные эмбрионы погибают на ранних стадиях развития, что является одной из причин спонтанного прерывания беременности на ранних сроках. В то же время нали чие полиплоидных клеток в некоторых органах человека не является патологией. Например, полиплоидные клетки встречаются в сердечной мышце, особенно в предсердиях, в печени и некоторых железах. Они могут быть одно- и двуядерными, уровень полиплоидии невысок и обычно находится в пределах 4—8п.
Мейоз - особый тип клеточного деления, возникновение которого связано с появлением полового размножения. При половом размножении два родителя — отец и мать — дают начало новому организму. При оплодотворении сливаются ядра половых клеток родителей, что увеличивает вдвое количество хромосом в зиготе. Следовательно, образование половых клеток должно быть связано с уменьшением количества хромосом в два раза, но таким образом, чтобы совокупность генетического материала обеспечивала преемственность поколений. Закономерное чередование репликации ДНК и, соответственно, xromocom, митозов и мейозов обеспечивает сохранение видоспецифического кариоти-па как в индивидуальном развитии — онтогенезе, так и в череде поколений организмов. В процессе мейоза из одной диплоидной клетки (2п=46) образуются 4 гаплоидные клетки (п=23). Кроме того, в мейозе происходит два вида перегруппировки генетического материала хромосом, т. е. два вида генетической рекомбинации: 1) независимое распределение гомологичных хромосом из разных пар к полюсам деления; 2) кроссинговер — обмен участками между гомологичными хромосомами. Эти процессы обеспечивают широчайший спектр наследственной изменчивости, генетическую неповторимость индивидов даже среди потомков одной пары родителей. Мейотическое деление у человека не имеет каких-то коренных отличий от мейоза других эукариот. Оно состоит из двух, следующих друг за другом делений, между которыми не происходит удвоения ДНК, а следовательно, и хромосом. Перед мейозом обязательно проходит интерфаза, в S-периоде которой ДНК реплицируется. Следовательно, в профазе первого мейотического деления выявляющиеся нитевидные хромосомы состоят из двух хроматид. Каждое из двух делений мейоза состоит из про-, мета-, ана- и телофазы с индексами I или II. Первое мейотическое деление протекает значительно дольше, чем второе. Самой длительной фазой первого мейотического деления является профаза, так как именно в этой фазе происходят такие сложные процессы, как образование бивалентов из гомологичных хромосом и кроссинговер. Профаза мейоза I подразделяется на 5 подфаз: лептотена, зиго-тена, пахитена, диплотена, диакинез. Лептотена — стадия тонких нитей. Ядра клеток, вступающих в мейоз, значительно крупнее других. В этих ядрах вместо хроматина выявляются очень тонкие и длинные нитевидные хромосомы. Они настолько тонкие и длинные, что их тру дно проследить по всей длине. Для лептотены характерно также появление на тонких хромосомах особых, напоминающих бусины структур — хромомеров. Это участки более сильно конденсированного хроматина. Число, размер и расположение хромомерных участков специфичны для каждой хромосомы. Выявление хромомерных структур отражает постепенный процесс конденсации хромосом из хроматина. Каждая хромосома состоит из двух сестринских хроматид, но их далеко не всегда удается различить под световым микроскопом, настолько близко они прилегают друг к другу. Зиготена - стадия сливающихся нитей. На этой стадии гомологичные хромосомы находят друг друга и сливаются (конъюгируют) с образованием бивалента. У человека 46 хромосом (23 пары гомологичных хромосом), следовательно, количество бивалентов равно 23 и соответствует гаплоидному набору хромосом. Бивалент - это стабильная структура, состоящая из двух гомологичных хромосом, или 4-х хроматид. Стабильность этой структуры поддерживается с помощью специфических белков синалто-немного комплекса. Объединение гомологов чаще всего начинается на концах хромосом (в теломерах), а также в центромерных районах. Позднее по длине соединяющихся хромосом (внутри бивалента) между хромосомами формируются белковые тяжи синап-тонемного комплекса, сближающие хромосомы.
В настоящее время показана специфичность хромомерного строения индивидуальных бивалентов человека в мужском и женском мейозе, т.е. по рисунку хромомер можно определить, какая пара хромосом образует тот или иной бивалент, даже если по размеру и общей морфологии биваленты одинаковы. Конъюгация гомологичных хромосом с образованием бивалентов является обязательным процессом для всех хромосом человека, включая короткие и половые хромосомы. Показано, что конъюгация происходит не только между половыми хромосомами Х и X, но также между Х- и Y-хромосомами, несмотря на большую разницу в их морфологии (размерах). В процессе сперматогенеза половой бивалент из Х- и Y-хромосом начинает конъюгировать раньше других хромосом. Конъюгируют между собой часть района короткого плеча Х- и короткое плечо Y-хромосомы. Эксперименты по гибридизации ДНК показали, что эти районы гомологичны между собой. Негомологичные участки хромосом Х и Y остаются свободными. Зиготена заканчивается образованием 23 бивалентов. Пахитена — стадия прохождения кроссинговера. В пахитене хромосомы выявляются в виде толстых нитей, так как они представлены бивалентами. Именно в бивалентах происходит кроссинговер — взаимный обмен идентичными участками по длине гомологичных хромосом. Генетическим следствием кроссинговера является рекомбинация сцепленных генов, что обеспечивает широкую генетическую изменчивость гамет. Морфологически этот процесс в пахитене уловить нельзя. Для умозрительного восприятия его можно изобразить на схеме (пахитена и диплотена). Кроссинговер — это процесс, происходящий со сложными пространственными изменениями суперспирализованных участков молекул ДНК несестринских хроматид с использованием целого комплекса ферментов, возможно объединенных вместе в специализированную структуру. Кроссинговер является обязательным процессом в каждом биваленте. Не исключено, что отсутствие кроссинговера в каком-то биваленте может быть запирающим фактором мейоза. В дальнейшем, в диплотене, когда начинают расходиться гомологичные хромосомы, в бивалентах, выявляются те места, где происходил процесс кроссинговера. Это места длительного соединения гомологичных хромосом во время разрушения бивалентов. Поскольку морфологически они напоминают греческую букву «X», их называют хиазмами. В зоне хиазм видно, что в перекрест вовлекаются только две хроматиды из четырех (по одной от каждого гомолога).
Диплотена – стадия двойных нитей. На этой стадии синаптонемный комплекс разрушается, и гомологи отталкиваются друг от друга, оставаясь соединенными только в районе хиазм, где по-прежнему сохраняется структура синаптонемного комплекса. Так как количество хиазм соответствует одному событию кроссинговера, в котором участвуют две несестринские хроматиды, то по количеству хиазм можно судить об интенсивности процесса кроссинговера. Общее число хиазм на хромосомный набор человека колеблется от 35 до 66. Некоторые биваленты могут содержать несколько хиазм. Среднее количество хиазм на бивалент около двух. Диакинез — стадия, завершающая профазу мейоза I. Она является переходной к метафазе. Число хиазм уменьшается, биваленты укорачиваются, разрушается ядро, начинает формироваться веретено деления.
Метафаза I — биваленты выстраиваются в экваториальной плоскости в цитоплазме. Центромеры хромосом расположены на экваторе, к ним прикреплены нити веретена деления. Число выстроенных бивалентов соответствует гаплоидному набору хромосом и для человека равно 23. Анафаза I — расхождение гомологичных хромосом к противоположным полюсам клетки. Каждая хромосома состоит из двух сестринских хроматид. Телофаза I. В этой фазе происходит образование двух дочерних ядер, каждое из которых содержит гаплоидное число хромосом, равное 23. Каждая хромосома состоит из двух сестринских хроматид. Промежуток между двумя последующими делениями мейоза очень небольшой. Почти сразу начинается второе мейотическое деление. Оно идет по схеме митоза: 23 хромосомы, состоящие из парных сестринских хроматид, связанных в центромерных участках, в каждом из двух образованных ядер проходят профазу и мета-фазу. В анафазе они разъединяются, и сестринские хроматиды каждой хромосомы расходятся к противоположным полюсам, в результате чего образуется четыре гаплоидных ядра. Рекомбинация генетического материала в мейозе происходит не только за счет процесса кроссинговера. В анафазе первого мейоти-ческого деления происходит случайное распределение по отношению к полюсам клетки гомологичных хромосом из каждого бивалента. Это приводит к большому числу возможных комбинаций отцовских и материнских хромосом в гаметах. Рассмотрим процесс подробнее на простом примере.
Проанализируем распределение первой и второй пары гомологичных хромосом в анафазе I. Известно, что в каждой паре гомологичных хромосом в процессе оплодотворения одна хромосома приходит из гаметы отца, другая - из гаметы матери. Обозначим заглавными буквами хромосомы отца, а строчными - хромосомы матери. Таким образом, А и а — первая пара хромосом, В и b - вторая пара. В профазе I образуются биваленты. В метафазе I они выстраиваются в экваториальной плоскости: А//а, В// b. В анафазе I гомологичные хромосомы из бивалентов расходятся к противоположным полюсам: к одному полюсу пойдут хромосомы А и В, т.е. отцовские, а к другому, а и b, т.е. материнские. Но у этого события может быть и другой исход, когда расположение хромосом в метафазе на экваторе будет другим: А//а, b//В. Тогда к одному полюсу пойдут хромосомы А и b, а к другому - а и В, т.е. сочетание хромосом на полюсах будет содержать одну отцовскую и одну материнскую. Наличие двух пар гомологичных хромосом обеспечивает, как мы видим, образование четырех типов гамет, качественно отличающихся друг от друга сочетанием отцовских и материнских хромосом. У человека 23 пары хромосом. Разнообразие гамет оценивается как 223. Это примерно 10 миллионов вариантов разнообразных сочетаний отцовских и материнских хромосом из каждой пары гомологов. При оплодотворении практически равновероятна встреча любого из сперматозоидов с овулировавшей яйцеклеткой. Это увеличивает число возможных генотипов детей (223). Частота генетической рекомбинации в результате независимого распределения разных пар гомологов выше, чем частота рекомбинации в результате кроссинговера. Оплодотворение и развитие Оплодотворение - сложный процесс, приводящий к слиянию яйцеклетки и сперматозоида, объединению их ядер, образованию диплоидной зиготы, из которой впоследствии будет развиваться новый организм. Генетическое значение процесса оплодотворения состоит в объединении гаплоидных наборов хромосом отцовских и материнских гамет, отличающихся высокой степенью генетического разнообразия.
Несмотря на количественное постоянство хромосомного набора, это обеспечивает генетическое разнообразие особей. В результате процесса оплодотворения восстанавливается диплоидность зиготы, что само по себе является толчком к последующим делениям митозом. У человека, как и у других млекопитающих, процесс оплодотворения, который начинается с проникновения сперматозоида в ооцит второго порядка, запускает механизм созревания яйцеклетки. В ооците второго порядка второе мейотическое деление останавливается на стадии метафазы. Завершение развития яйцеклетки наблюдается только после оплодотворения. Оплодотворение состоит из трех последовательных этапов. Первый этап характеризуется сближением сперматозоида и яйцеклетки до их контакта. Второй этап начинается с момента прикрепления сперматозоида к поверхности яйцеклетки и осуществления контактных взаимодействий между ними. Третий период начинается после проникновения сперматозоида в яйцеклетку и завершается объединением их ядер. Оплодотворение происходит в верхних концах фаллопиевых труб. Сперматозоиды сохраняют жизнеспособность в женских половых путях в течение 1-3 суток, но остаются высокофертильными лишь 12-24 часа. Сперматозоид способен оплодотворить ооцит только после того, как он проведет в половых путях несколько часов. Спермии скапливаются вокруг ооцита второго порядка. В головке одного из спермиев происходит очень быстрая акросомальная реакция. Она состоит в том, что разрываются мембраны головки спермия и ферменты акросомы, особой клеточной структуры сперматозоида, «переваривают» клеточные слои, окружающие ооцит. Затем спермйй проникает внутрь ооцита. У человека сперматозоид входит в ооцит целиком. Время акросомной реакции не превышает 20 секунд. После проникновения в яйцеклетку одного из спермиев разрываются многочисленные вакуоли, расположенные под мембраной яйцеклетки, и из них освобождается вещество, под действием которого zona pellucida утолщается и образует непроницаемую преграду поверх яйцеклетки — оболочку оплодотворения. Она препятствует проникновению в ооцит других спермиев. У человека проникновение сперматозоида в ооцит является сигналом для завершения второго мейотического деления, которое приводит к образованию гаплоидной яйцеклетки и сопутствующих клеток (полярных телец), которые дегенерируют. В это же время-в цитоплазме ооцита рассасывается хвост сперматозоида. Следующий этап процесса оплодотворения связан с объединением генетического материала яйцеклетки и сперматозоида в одно диплоидное ядро. Высококондеисированное ядро сперматозоида начинает набухать, хроматин разрыхляется, и ядро превращается в структуру, называемую мужским пронуклеусом. Аналогичные изменения происходят в женском ядре, что приводит к образованию женского про-нуклеуса. В процессе формирования пронуклеусов происходит синтез ДНК и в мужском, и вженском ядре (In 2C). В результате каждая хромосома удваивается и состоит уже из двух сестринских хроматид. После того, как репликация ДНК закончена, пронуклеусы перемещаются к центру яйцеклетки и их мембраны разрушаются. Отцовские и материнские хромосомы, каждая из которых имеет две хроматиды, прикрепляются к нитям сформировавшегося веретена деления. Затем хромосомы выстраиваются по экватору веретена. В это время наблюдается картина, характерная для метафазы митоза. Объединение отцовских и материнских хромосом в общем митозе переводит оплодотворенную яйцеклетку в качественно новое состояние — зиготу» Зигота проходит стадии анафазы и телофазы и завершает свое первое митотическое деление. Образуются две дочерние диплоидные клетки - бластомеры. Генетический материал бластомеров представляет собой совокупность хромосом отца и матери. В каждой паре гомологичных хромосом одна хромосома отцовская, а другая материнская. Процесс индивидуального развития начинается с момента оплодотворения. Зигота делится митотически, затем проходят этапы пренатального развития: бластула, гаструла, нейруляция, имплантация в стенку матки, закладка и формирование органов и систем органов, рост плода. Пренатальный период сменяется перинатальным, который начинается с 28-й недели беременности, включает роды и заканчивается через 7 дней после родов. После этого начинается постнатальный период, включающий в себя всю жизнь человека от рождения до смерти. Процесс индивидуального развития имеет ряд критических периодов, когда организм наиболее уязвим к различным факторам окружающей среды. Нормальное развитие особи в течение эмбриогенеза сопряжено с активным делением клеток, морфогенетическим движением зародышевых листков, с процессами дифференцировки и органогенеза. Тератогенные факторы могут изменять способность клеток к размножению и перемещению из одного места в другое, нарушать процесс приобретения клетками определенной специализации и в конечном итоге вызвать развитие уродств. К основным механизмам развития пороков на тканевом уровне относятся: гибель отдельных клеточных масс, замедление распада и рассасывания отмирающих клеток, нарушение взаимодействия тканей между собой. В результате тот или иной орган йедоразвивается или развивается неправильно. Например, если смещается устье аорты, то развивается врожденный порок сердца. На ранних стадиях эмбриогенеза выделяют так называемые критические периоды, во время которых развивающийся орган-особо чувствителен к факторам Окружающей среды. Если женщина заболеет краснухой между 3-й И 9-й неделями беременности, то возникает риск развития у плода таких пороков, как порок сердца, катаракта, глухота. В другие сроки краснуха не вызывает пороков развития плода. Сходное действие предполагается у других вирусных инфекций (гриппа, оспы, паротита). Поскольку тератогенные факторы не нарушают генетических структур, такие пороки развития не наследуются, т. е. риск повторного рождения ребенка с аналогичным пороком предельно мал. К препаратам, способным вызвать образование врожденных пороков развития, относятся транквилизаторы, противосудорожные, гормональные и противоопухолевые средства. Стероидные гормоны, входящие в состав многих контрацептивов, которые женщины могут принимать, не зная о наличии беременности, относятся к числу факторов риска развития врожденных пороков. Широко применяемые в промышленности и сельском хозяйстве бензол, фенолы, хлоропрен, формальдегид, ядохимикаты, а также свинец и пары ртути обладают токсическим действием. Их воздействие может вызвать самопроизвольный аборт, внутриутробную смерть плода или рождение ослабленного ребенка.
Биопсия хориона и плаценты К числу перспективных методов пренатальной диагностики относится биопсия плода и плаценты. Плацентобиопсия проводится на более ранних, чем УЗИ, сроках беременности (7-9 недель). Ворсинки хориона берут особым шприцом с помощью гибкого катетера через шейку матки (рис. 1, 2). Затем их подвергают лабораторной диагностике с помощью цитологических, биохимических, молекулярно-генетических методов. В случае выявления наследственных заболеваний у плода беременность прерывают. Одним из осложнений хорионбиопсии является относительно высокая частота спонтанных абортов (выкидышей). Общие потери плода после хорионбиопсии в среднем составляют 2,5-3%, а в некоторых случаях и выше, что ограничивает применение этого метода. Каких-либо нарушений плаценты, роста плода, появлений пороков развития и увеличения перинатальной смертности после хорионбиопсии не отмечается. Однако, существуют данные, что ранняя хорионбиопсия может вызывать поперечные врожденные апмутации конечностей (редукционные пороки). По этой причине в последнее время хорионбиопсию не рекомендуется проводить ранее 8-й недели, а плацентобиопсию — 12-й недели беременности. Амниоцентез Амниоцентез — прокол плодного пузыря для взятия 8-10 мл околоплодной жидкости с находящимися в ней слущенными клетками амниона и плода. Последние являются основным субстратом для цитологических и биохимических исследований. Этот метод является наиболее распространенным и доступным. Проводится на 15-18-й неделе беременности. Риск осложнения ее течения незначителен (0,2%). Амниоцентез делают чрезбрюшинно под контролем УЗИ, чтобы не повредить плаценту. Чрезвлагалищный амниоцентез применяется редко. С помощью этого метода диагностируют многие хромосомные нарушения, болезни, сцепленные с полом, болезни обмена веществ (болезнь Тея-Сакса, мукополисахаридозы, гликогенозы, фенилкетонурия и др.). Кордоцентез В этом случае берутся образцы крови (лейкоциты) из пуповинных сосудов плода, для цитогенетического, молекулярно-генетического и биохимического анализов крови. Кровь более удобна для исследования, чем клетки амниотической жидкости, т.к. лимфоциты быстрее и надежнее культивируются. Кордоцентез проводится под контролем УЗИ на 18-22-й неделе беременности. Его используют для диагностики хромосомных болезней, таких, как: гемоглобинопатии, энзимопатии, различные иммунодефицитные аномалии, синдром фрагильной (ломкой) Х-хромосомы и др. Фетоскопия Данный метод основан на прямом рассмотрении плода через специальный прибор — фетоскоп (тонкий эластичный зонд со специальной оптической системой). Зонд вводится в плоскость амниона через брюшную систему. Проводится на 18-23-й неделе беременности. Фетоскопия используется редко и только при особых показаниях, поскольку вхождение зонда в амниотическую жидкость может вызвать осложнение беременности. Выкидыши отмечаются в 7-8% случаев фетоскопии. Проще и безопаснее использовать УЗИ, способное выявить большинство пороков развития. Методы пренатальной диагностики постоянно совершенствуются и все шире применяются в профилактике наследственных заболеваний. Дальнейшее совершенствование и расширение применения этих методов — одна из задач современной медицинской генетики. Вопросы для самоконтроля: 1. Основная цель и задачи медико-генетического консультирования (МГК). 2. Перечислите показания для направления на медико-генетическое консультирование; основные этапы МГК. 3. Что такое прогноз потомства? 4. Что имеется в виду под термином "пренатальная диагностика"? Назовите основные показатели для проведения пренатальной диагностики. 5. Перечислите важнейшие методы дородовой диагностики. 6. Выберите два правильных ответа. Состояния, диагностируемые у плода с помощью УЗИ: 1) синдром Марфана; 2) редукционные пороки конечностей; 3) фенилкетонурия; 4) нарушения в развитии мозга. 7. Какой из указанных методов дает наибольший процент осложнений: а) фетоскопия; б) хорионбиопсия; в) кордиоцентез; г) УЗИ. Клеточная инженерия: · Культивируют клеточные культуры. Нарабатываются биологически активные вещества, размножаются редкие и ценные растения. · Гибридизация протопластов соматических клеток от · Создание гибридом: гибридизация лимфоцитов, образующих антитела, с раковыми клетками.
· Метод пересадки ядер соматических клеток в яйцеклетки. Возможно клонирование животных. Хромосомная инженерия: · — Введение в генотип или замещение одной пары гомологичных хромосом на другую. Создание «идеального сорта». · — Получение полиплоидных растений. Введение гена из одного организма в другой. Излюбленный объект — кишечная палочка. Уже получают гормон роста, инсулин, интерферон, помогающий справиться с вирусной инфекцией. Селекция животных Основные методы селекции животных. Создание пород домашних животных началось вслед за их приручением и одомашниванием, вероятно, 20—30 тыс. лет назад. Первым этапом было приручение животных. В результате бессознательной формы искусственного отбора происходило одомашнивание и изменение генофонда прирученных животных. Менялся их внешний вид, продуктивность, характер. Сохранялись те животные, которые могли размножаться в неволе, не проявляли агрессивности по отношению к человеку. Методический отбор привел к созданию всего многообразия пород домашних животных от небольшого числа видов диких предков. В селекции животных, по сравнению с селекцией растений, есть ряд особенностей. Во-первых, для животных характерно в основном половое размножение, поэтому любая порода является сложной гетерозиготной системой. Во-вторых, у них часто поздняя половозрелость, смена поколений происходит через несколько лет. В-третьих, потомство немногочисленное. Основными методами селекции животных являются гибридизация и отбор. Различают те же методы скрещивания — близкородственное скрещивание, инбридинг, и неродственное — аутбридинг. Инбридинг, как и у растений, приводит к депрессии. Отбор у животных проводится по экстерьеру (определенным параметрам внешнего строения), т. к. именно он является критерием породы. 1. Внутрипородное разведение: направлено на сохранение и улучшение породы. Практически выражается в отборе лучших производителей, выбраковке особей, не отвечающих требованиям породы. В племенных хозяйствах ведутся племенные книги, отражающие родословную, экстерьер и продуктивность животных за много поколений. 2. Межпородное скрещивание используют для создания новой породы. При этом часто проводят близкородственное скрещивание, родителей скрещивают с потомством, братьев с сестрами, это помогает получить большее число особей, обладающих нужными свойствами. Инбридинг сопровождается жестким постоянным отбором, обычно получают несколько линий, затем производят скрещивание разных линий. Хорошим примером может служить выведенная академиком М. Ф. Ивановым порода свиней — украинская белая степная. При создании этой породы использовались свиноматки местных украинских свиней с небольшой массой и невысоким качеством мяса и сала, но хорошо приспособленных к местным условиям. Самцами-производителями были хряки белой английской породы. Гибридное потомство вновь было скрещено с английскими хряками, в нескольких поколениях применялся инбридинг, были получены чистые линии, а при скрещивании их — родоначальники новой породы, которые по качеству мяса и массе не отличались от английской породы, по выносливости — от украинских свиней. 3. Использование эффекта гетерозиса. Часто при межпородном скрещивании в первом поколении проявляется 4. Испытание по потомству проводят для подбора самцов, у которых не проявляются некоторые качества (молочность и жирномолочность быков, яйценоскость петухов). Для этого производителей-самцов скрещивают с несколькими самками, оценивают продуктивность и другие качества дочерей, сравнивая их с материнскими и со среднепородными. 5. Искусственное осеменение используют для получения потомства от лучших самцов производителей, тем более что половые клетки можно хранить при температуре жидкого азота любое время. 6. С помощью гормональной суперовуляции и трансплан тации у выдающихся коров можно забирать десятки эмбрионов в год, а затем имплантировать их в других коров, эмбрионы также хранятся при температуре жидкого азота. Это даст возможность увеличить в несколько разчисло потомков от выдающихся производителей. 7. Отдаленная гибридизация, межвидовое скрещивание, известна с древних времен. Чаще всего межвидовые гибриды стерильны, у них нарушается мейоз, что приводит к нарушению гаметогенеза. С глубокой древности человек использует гибрид кобылицы с ослом — мула, который отличается выносливостью и долгожительством. Но иногда гаметогенез у отдаленных гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары, могут пастись высоко в горах, а, как мериносы, дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного крупного рогатого скота с яками и зебу. При скрещивании белуги и стерляди получен — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасем. Полиплоидия крайне редко встречается у животных. Интересен факт межвидового скрещивания тутового шелкопряда.
Тема 20. Популяционная генетика Борьба за существование Задачи: 1. Знакомство с различными формами борьбы за существование, которые осуществляются в природе. 2. Правильное определение форм борьбы за существование, сравнение их друг с другом. 3. Сформировать правильное отношение к проблеме реализации идеи «борьбы за существование» в природе и человеческом обществе. Основное содержание и методические материалы: Ч. Дарвин впервые пришел к мысли о том, что движущая сила всего эволюционного процесса есть результат взаимодействия организмов между собой и с внешней средой, т. е. борьбы за существование. Он указал, что несоответствие между возможностью видов к беспредельному размножению и ограниченностью ресурсов — главная причина борьбы за существование. Наиболее острой и напряженной является внутривидовая борьба, поскольку конкурирующие особи одного и того же вида предъявляют сходные или полностью идентичные требования к условиям, в которых они существуют (например, питаются одинаковыми кормовыми объектами, количество которых может быть ограниченным). Требования представителей разных биологических видов и разных экологических групп организмов к условиям внешней среды в той или иной степени расходятся, что нередко позволяет им ослабить остроту конкуренции друг с другом и мирно сосуществовать в общем ареале.
Образование новых видов. макроэволюция. Современная система органического мира Задачи:
Основное учебное содержание и методические материалы: При характеристике эволюционного процесса необходимо четко определить, что понимают современные биологи под макроэволюцией и микроэволюцией, так как эти термины постоянно встречаются в специальной литературе. В большинстве случаев под макроэволюцией понимают процесс образования новых семейств, отрядов, классов, типов и других надвидовых систематических единиц (таксономических групп живых существ). Под микроэволюцией понимают сложнейший эволюционный процесс возникновения нового вида. При этом новый вид превращается в обособленную совокупность организмов. Микроэволюцию нередко называют также видообразованием. Механизмы микроэволюции и макроэволюции принципиально сходны. Идеи Ч. Дарвина заложили основу для современного понимания процессов видообразования. Вспоминая историю своих исследований, он подчеркивал, что долгое время после открытия принципа естественного отбора он не мог решить вопрос о возникновении разнообразия видов. Термин «видообразование» означает расщепление одного вида на несколько, т. е. увеличение числа видов. Виды стремятся к максимально возможному заполнению мест обитания и освоению различных способов существования. При этом происходит дивергенция (расхождение признаков) у групп особей, осваивающих различную среду обитания. Однако бывают ситуации, когда представители эволюционно и систематически удаленных друг от друга. Итогом экологического видообразования является, например, формирование разных популяций речного окуня, обитающих в одном водоеме, но на разных глубинах. Как уже отмечалось выше, видообразование лежит в основе макроэволюционного процесса. Отображением же эволюции в целом является современная классификация живых организмов. Система органического мира, предложенная К. Линнеем, впоследствии была существенно изменена. В самом общем виде современную классификацию органического мира можно представить следующим образом.
Важно также понимать, что лишайники — это особая, «сборная» группа организмов. В состав лишайника обязательно входит представитель царства грибов. Вторым компонентом лишайника является либо, одноклеточная водоросль, либо представитель группы сине – зеленых. Между организмами, входящими в состав тела лишайника, складываются особые взаимоотношения, часто выражающиеся в форме взаимопомощи. Гифы гриба, оплетающие клетки водоросли, доставляют к ним минеральные соли, растворенные в воде. Одноклеточная водоросль (или представитель сине-зеленых) осуществляет процесс фотосинтеза и созданными таким образом органическими веществами «делится» с грибом. Наши знания о строении и жизнедеятельности лишайников не позволяют отнести их к какой-то одной систематической группе (растениям, грибам или дробянкам). Каждое из перечисленных царств, включает в себя последовательно несколько более мелких систематических единиц. Рассмотрим эту ситуацию на примере царства животных и царства растений.
Класс (Млекопитающие) Класс (Однодольные)
Семейство (Кошачьи) Семейство (Злаковые)
Род (Кошка) Род (Пырей)
Вид (Кошка домашняя) Вид (Пырей ползучий)
При классификации организмов современные ученые-систематики обращают внимание на целый комплекс признаков, характеризующих данное растение или животное. Рассмотрим некоторые признаки, на основании которых цветковые растения разделяются на два класса — Однодольные и Двудольные.
Эволюционное учение
Задачи: 1. Систематизировать и обобщить знания об основных положениях эволюционного учения, проверить качество их усвоения. 2. Создать условия для развития мыслительных способностей в процессе решения проблемных вопросов и выполнения заданий об эволюции органического мира. 3. Сформировать убежденность в том, что эволюционное учение является одним из наиболее фундаментальных теоретических обобщений современной биологии.
Основное учебное содержание и методические материалы:
На заключительно-обобщающем занятии делается вывод о том, что классические положения эволюционного учения Ч. Дарвина на определенном этапе развития науки вступили в противоречие с некоторыми открытыми впоследствии биологическими закономерностями (в частности, из области генетики). Это привело к уточнению первоначальных эволюционных представлений, обогащению их новыми фактами, дальнейшему развитию и обоснованию с позиций иных биологических наук. В разработку современной теории эволюции внесли свой вклад такие отечественные ученые, как Н. И. Вавилов, В. О. Ковалевский, А. Н. Северцов, Н. В. Тимофеев-Ресовский, С. С. Четвериков, И. И. Шмальгаузен и др. Как следствие этого процесса в середине XX в. сформировалась синтетическая теория эволюции. Задача 1 В одном из зоопарков Индии у пары тигров с нормальной окраской родился тигр альбинос (такие формы встречаются довольно редко). Почему у фенотипически одинаковых животных родился детеныш без пигментации? Какие действия должны провести селекционеры, чтобы как можно быстрее получить максимальное количество тигрят с данным признаком? А - нормальная пигментация Р: ♀ Аа х ♂ Аа
 а – отсутствие пигментации а – отсутствие пигментации

нет расщеплений нет расщеплений F1: АА, Аа, Аа, аа
пигментир. альбинос
Самостоятельная работа «Создай лицо ребенка» У человека целый ряд признаков лицевого отдела черепа относиться к разряду менделирующих. Условие задачи: Братья – монозиготные близнецы женились на моногозиготных сестрах близнецах. Будут ли дети этих семейных пар также похожи между собой, как и их родители? Родители-близнецы гетерозиготны по следующим признакам:
1. Форма лица: круглая (А-), квадратная (аа) 2. Форма подбородка: круглая (В-), квадратная (вв) 3. Ямочка на подбородке: присутствует (С-), отсутствует (сс) 4. Тип волос: курчавые (РР), вьющиеся (Рр), прямые (рр) 5. Размер глаз: большие (ЕЕ), средние (Ее), маленькие (ее) 6. Расстояние между глазами: близко посажены (ТТ), среднее расстояние (Тt), широко расставлены (tt) 7. Ресницы: длинные (L-), короткие (ll) 8. Толщина бровей: густые (К-), тонкие (кк) 9. Расположение бровей: не соединяются (N-), соединяются (nn) 10. Размер носа большой (ГГ), средний (Гг), маленький (гг) 11. Форма носа: круглый (О-), заостренный (оо) 12. Размер рта: большой (ММ), средний (Мм), маленький (мм) 13. Толщина губ: полные (D-), тонкие (dd) 14. Цвет глаз: темный (Н-), светлый (нн) 15. Уши – с нормальной мочкой (Ф), с приросшей (фф)
Ход работы: Возьмите монетку и подбрасывайте ее на столе: «орел» - доминантный признак; «решка» - рецессивный признак. Так как родители гетерозиготны, вероятность передачи ребенку доминантного или рецессивного аллеля – 1:1. Подбрасывая монетку, определите сначала пол ребенка («решка» – девочка, «орел» – мальчик), а затем аллели, которые родители передадут своему малышу. Заполните табличку. В конце работы постарайтесь нарисовать портрет ребенка в возрасте 5-6 лет. Сравните портреты, полученные другими студентами, и ответьте на заданный вопрос.
Форма лица:
круглая (АА; Аа) квадратная (аа) Очертания подбородка:
Форма глаз:
удлиненная (АА, аа) круглая (аа) Расположение углов глаз:
горизонтальное (НН, Hh ) угол поднят кверху ( hh ) Ресницы:
длинные ( LL , LI ) короткие (11) Размеры рта:
большой (ММ) средний (Мм) маленький (мм) Выпуклость губ:
очень умеренно не выпуклые Форма носа:
круглый ( Rr ) заостренный (гг) Мочка уха:
Веснушки на щеках:
Ec ть( FF ; Ff ) нет ( ff ) Веснушки на лбу:
есть ( ZZ , Zz ) нет {гг)
Цвет кожи Наследуется полимерными генами, которые обладают аддитивным (суммарным) эффектом. Чем больше доминантных генов присутствует в генотипе, тем более активно проявляется пигментация кожи: 6 доминантных генов — очень черная кожа 5 доминантных генов — очень коричневая кожа 3 доминантных гена — коричневая кожа 2 доминантных гена — светло-коричневая кожа 1 доминантный ген — смуглая 0 доминантных генов — белая Цвет волос Подобно цвету кожи, цвет волос наследуется четырьмя полимерными генами. 8 доминантных генов — черные волосы 7 доминантных генов— очень коричневые волосы 6 доминантных генов — темно-каштановые 5 доминантных генов — каштановые, 4 доминантных гена — светло-русые, 3 доминантных гена — рыжеватая блондинка 2 доминантных гена — блондинка 1 доминантный ген — очень светлая блондинка 0 доминантных генов — белые Рыжие волосы определяются единственным геном, представленным двумя аллелями Red (R) и red (г): RR — ярко-рыжие; Rr — светло-рыжие; гг — отсутствие рыжих волос. Рыжие волосы проявляются только в том случае, если у человека меньше 6 доминантных аллелей по выше представленным полимерным генам. З а д а ч а 1 У мухи дрозофилы серый цвет тела доминирует над чёрным. При скрещивании серых и чёрных мух в потомстве половина особей имела серую окраску, половина – чёрную: а) определите генотипы родительских форм; б) подчиняется ли наследование окраски тела дрозофилы законам Менделя? З а д а ч а 2 При скрещивании норок коричневого и голубовато – серого цвета получен приплод, все особи которого имели коричневую окраску. При скрещивании зверьков первого поколения получен приплод, в котором 167 норок имели коричневый цвет, а 58 – голубовато – серый: а) ген какой окраски является доминантным? б) относится ли этот признак (окраска) к менделирующим? II. Множественный аллелизм
При анализе наследования ряда признаков выявляются отклонения от законов Менделя в распределении генотипов и фенотипов в потомстве. Одной из причин таких отклонений является множественный аллелизм. Он проявляется в увеличении числа аллельных генов, контролирующих тот или иной признак. З а д а ч а 1 У мышей длина хвоста определяется серией аллельных генов: А, а, а1, а2. Гетерозиготы Аа имеют укороченный хвост, Аа1 – бесхвостые, аа1, аа2 и а1а2 имеют нормальную длину хвоста. Гомозиготные организмы (АА, аа, а1а1 и т.д.) погибают на стадии эмбрионов: а) определите вероятные генотипы и фенотипы потомства от скрещивания бесхвостых мышей с мышами, имеющими укороченный хвост; б) будет ли распределение генотипов соответствовать законам Менделя? З а д а ч а 2 У кроликов разные варианты окраски меха (распределение пигментированных тёмных пятен) контролируются четырьмя аллельными генами: С, ch, cch,c. Ген С доминирует над остальными и обусловливает сплошную тёмную окраску тела, аллель ch определяет тёмную окраску лап, хвоста, ушей и носа (гималайская окраска), он рецессивен по отношению к гену С и доминантен по отношению к гену с. Ген cch определяет серую шиншилловую окраску; аллель с обусловливает белую окраску тела, т.е. полное отсутствие пигмента (альбинизм); cchch – большие чёрно – белые пятна ("бабочка"). а) кролик со сплошной пигментацией, гетерозиготный по гену альбинизма, скрещен с гималайским кроликом, также гетерозиготным по гену альбинизма. Какой генотип и фенотип будет иметь потомство F1 от этого скрещивания? б) школьники в живом уголке скрестили кроликов со сплошной тёмной пигментацией с гималайским и получили 81 крольчонка. Из них 41 имел сплошную тёмную окраску, 19 были гималайскими и 21 – альбиносами. Определите генотипы родительских форм и потомков.
З а д а ч а 1 У мух дрозофил узкие глаза доминируют над нормальными, а загнутые кверху крылья над прямыми. Гомозиготная по признакам узких глаз и загнутых кверху крыльев муха скрещена с мухой с нормальными глазами и прямыми крыльями: а) какой генотип и фенотип будет у потомства F1 и F2; б) какая часть узкоглазых с загнутыми кверху крыльями особей потомства F2 будет гомозиготна по обоим признакам? З а д а ч а 2 У крупного рогатого скота ген комолости доминирует над геном рогатости, а ген чёрного цвета тела над красным. Обе пары генов не сцепленные: а) скрещивается гетерозиготный по обоим признакам чёрный комолый бык с такой же коровой. Какими окажутся телята? б) в хозяйстве от 1000 рогаых красных коров получено 984 телёнка, из них красных 472, комолых 483, рогатых 581. Определите генотипы родителей и количество чёрных телят (в процентах).
З а д а ч а 1 Какова вероятность рождения в браке тетрагетерозигот детей с фенотипами: А-В-ccD-; aabbccD-? З а д а ч а 2 Установите генотип людей, имеющих следующие признаки: близорукий мужчина, "правша", с веснушками на лице и низким голосом (бас); все признаки доминантные. Если вы решили, что люди с таким фенотипом могут иметь различные генотипы, то укажите эти генотипы. З а д а ч а 3 Полидактилия, близорукость и отсутствие малых коренных зубов передаются как доминантные аутосомные признаки. Гены всех трёх признаков находятся в разных парах хромосом: а) какова вероятность рождения детей без аномалий, если оба родителя страдают всеми тремя недостатками, но гетерозиготны по всем трём парам генов? б) определите вероятность рождения детей без аномалий в семье, о которой известно, что бабушка по линии жены была шестипалой, а дедушка близорукий. В отношении других признаков они нормальны. Дочь же унаследовала от своих родителей обе аномалии. Бабушка по линии мужа не имела малых коренных зубов, имела нормальное зрение и пятипалую кисть. Дедушка был нормален в отношении всех трёх признаков. Сын унаследовал аномалию матери.
I . Эпистаз В 1932 году Кальвин Бриджес доказал, что гены лежащие в разных хромосомных парах и отвечающие за один и тот же признак не всегда взаимно дополняют друг друга. Существует явление совершенно обратное, в этом случае 1 ген подавляет фенотипическое проявление гена другого локуса. Это явление названо эпистазом. Ген – подавитель Вильям Бетсон назвал ингибитором. Он может называться супрессором или эпистатическим. Подавляемый ген называется гипостатическим.
двойной рецессивный Задание II. Рассмотрите пример наследования окраски тыквы (признак – сложный, наследуется дигенно) У – синтез желтого пигмента (тыква желтая), у – синтез зеленого пигмента (тыква зеленая), W – ген–супрессор, подавляющий синтез пигмента, w – синтез не подавляется, тыква пигментирована, то есть окрашена.
WWYY x wwyy WwYy x WwYy Белая зеленая бел. бел. WY ___________wy F 2: 12 : 3 : 1 F1: WwYy Единообразие, тыквы по окраске белоплодные. Доминантный аллель W ингибировал синтез и желтого и зеленого пигмента. W > Y, W > y. Мы разобрали пример простого доминантного эпистаза.
Задача 1. У одной из линий лабораторных мышей: А – детерминирует серый цвет шерсти, а - черный цвет, В – синтез пигмента не нарушается (проявитель окраски), в - синтез пигмента нарушен (подавитель окраски).
ААВВ х аавв серый белая АаВв х АаВв серые серые
А-В - ааВ - А-вв аавв Серые черн. Белые белые 9 : 3 : 3 : 1 Рецессивные аллели одного гена в гомозиготном состоянии ингибировали фенотипическое проявление доминантных и рецессивных аллелей другого гена вв > A , вв > аа. Это пример простого рецессивного эпистаза.
При эпистазе может наблюдаться расщепление 12 : 3 : 1, а может и 13 : 3.
Задача 2. В генотипе кур есть ген, определяющий окраску пера: С – перо окрашено в результате синтеза пигмента, с – перо не окрашено (пигмента нет). Кроме того, у кур есть другой ген, который проявляет эпистаз по отношению к гену окраски: I – ген-супрессор, i - не подавляет синтез пигмента. Куры породы Леггорн имеют белое оперение и генотип IICC, куры породы Виандот имеют белое оперение и генотип iicc. Потомки F1 от скрещивания этих линий имеют генотипы IiCc и такое же белое оперение. Однако потомство F2 состоит из белых и окрашенных особей в соотношении 13 : 3. I- - - iiCC iiCC iiCC Белые Окрашенные Предстоит разобрать пример двойного рецессивного эпистаза, дающего расщепление 9:7. Задача 3. Фиолетовую окраску цветка детерминируют два неаллельных гена, при этом: А – определяет синтез красного пигмента, В – определяет синтез синего пигмента. Рецессивный аллель а эпистатичен по отношению к доминантному аллелю В, рецессивный аллель в эпистатичен по отношению к доминантному аллелю А (подавляет синтез пигмента).
ААВВ х аавв АаВв х АаВв фиолет. пигмента нет, форма белая АВ Ав аВ ав
Ав фиол. бел. фиол. бел. F1: АаВв аВ фиол. фиол. бел. бел. единообразие, ав фиол. бел. бел. бел. фиолетов.
При эпистазе расщепление идет в соотношениях: 12 : 3 : 1, 9 : 3 : 4, 13 : 3, редко 9:7. Этот тип взаимодействия наиболее характерен для генов, которые участвуют в работе иммунной системы человека, а так же генов, контролирующих, регулирующих работу других генов в онтогенезе. В настоящее время в научной литературе два термина «эпистаз» и «супрессия» используются как синонимы, но чаще – термин «супрессия». III . Полимерия Явление, при котором один фенотипический признак детерминируется действием сразу нескольких однотипных генов.Однотипные гены часто называются эквивалентными или полимерными и обозначаются одной и той же буквой, но с разными индексами.
Кумулятивная полимерия Задание I . Проанализируйте механизм наследования пигментации кожи человека. Признак наследуется полимерными генами, которые обладают суммарным (аддитивным) эффектом. Общее число доминантных аллей полимерных генов влияет на степень выраженности фенотипического признака – чем больше доминантных аллей в генотипе, тем больше синтезируется меланина и тем интенсивнее будет пигментация кожи. Если взять 3 полимерных гена, то генотипы могут иметь совершенно разные фенотипы: А1А1А2А2А3А3, 6 доминантных аллей – очень черная кожа; А1а1А2А2А3А3, 5 доминантных аллей – очень коричневая; а1а1А2А2А3А3, 4 доминантных аллей – темно–коричневая; 3 - коричневая; 2 - светло- коричневая; 1 - смуглая; 0 - светлая европеоидная. Р: А1А1А2А2А3А3 х а1а1а2а2а3а3 Черный эфиоп Русская девушка
F1: А1а1А2а2А3 а3 Коричневый мулат (Явление альбинизма не рассматривается).
Задача 1: В браке гетерозиготных мулатов рождается ребенок с белой кожей, а затем негритенок. Возможно ли такое? Чтобы облегчить запись, предположим, что признак определяется двумя полимерными генами. Р : А1а1А2а2 х А1а1А2а2 Мулатка мулат гаметы матери и отца совпадут: А1А2 А1а2 а1А2 а1а2 Расщепление: 4 дом. А – негритенок, 3 дом. А – темно-коричневый, 2 дом. А - коричневый мулат, 1 дом. А – желтый, рецессивные аллели – белый цвет кожи. 1 : 4 : 6 : 4 : 1 негр корич. мулат жел. белый Еще раз подчеркнем, что при формировании данного признака не важно, какому гену (1,2) принадлежат доминантные аллели, важно их общее количество в генотипе. При кумулятивной полимерии эффекты доминантных аллелей складываются в своем действии, и степень выраженности признака прямо пропорциональна количеству доминантных генов в генотипе.
Задача 2: Подобно цвету кожи наследуется цвет волос человека. По всей видимости, цвет волос определяют сразу 4 полимерных гена. Если в генотипе: 4 дом.ал. – русые; 8 дом.ал.- волосы черные; 3 дом.ал. – светло-русый блондин; 7 дом.ал.- темно-коричн.; 2 дом.ал. – блондин; 6 дом.ал. – темно-каштан.; 1 дом.ал. – очень светлый блондин; 5 дом.ал. – каштановые; рецессивные ал. – белые волосы. Изменение цвета волос от темного до светлого связано с уменьшением пигмента – меланина. Многих интересует, как наследуется рыжий цвет волос? Существует другой пигмент – феумеланин, который несколько отличается по составу от меланина. Образование феумеланина приводит к появлению рыжих волос. Рыжие волосы определяются независимым геном, представленным двумя аллелями: R – доминантной, r – рецессивной. Их сочетание в генотипе: RR – ярко-рыжие, Rr - светло-рыжие, rr - рыжий цвет отсутствует.Надо помнить о том, что рыжий цвет заметен фенотипически только тогда, когда в генотипе меньше 6 доминантных аллелей по выше названным полимерным генам. Полимерный тип взаимодействия характерен не только для качественных, но и большинства количественных признаков. Есть мнение, что рост человека наследуется полимерными генами. Некумулятивная полимерия Чтобы признак проявил себя по максимуму, достаточно иметь в генотипе хотя бы 1 доминантный аллель из серии полимерных генов. Можно сказать иначе: при некумулятивной полимерии не важно количество доминантных аллелей в генотипе, а важно присутствие хотя бы одного из них, чтобы признак имел свое полное фенотипическое проявление. Пример: чистые линии кур с оперенными ногами – гомозиготны по двум однотипным доминантным генам (А1А1А2А2); голоногие куры – рецессивны. Гибриды F1 (А1а1А2а2) не уступают по качеству оперения ног доминантной форме. У потомства F2 соотношение расщепления составит 15 : 1. Явление полимерии имеет важное значение в сельском хозяйстве. Данный механизм лежит в наследовании таких признаков как содержание сахара, витаминов в плодах и корнеплодах, длины колоса, початка кукурузы, яйценоскости, молочности, массы тела. Задача1. У крупного рогатого скота в генотипе есть ген, доминантный аллель которого (А) определяет сплошную окраску шерсти, а рецессивный (а) - пегую (т.е. пятнистую). Доминантный аллель другого гена - В – детерминирует черный пигмент, рецессивный аллель в – рыжий пигмент. Р: ААВВ х аавв сплошной пятнистая черный рыжая
F 1: АаВв – единообразие (сплошное черное потомство).
В генотипе животных присутствует ген-модификатор: С – усиление непигментных участков, с – уменьшение непигментных участков.
Бык с генотипом ААВВСС – сплошной черный, у него нет непигментированных участков; корова с генотипом ааввСС почти вся белая с маленькими рыжими пятнами; корова с генотипом ааввСс – со средними бело-рыжими пятнами; с генотипом ааввсс – почти вся рыжая с маленькими белыми пятнышками. Действие модификатора обнаруживается в присутствии гомозиготы по рецессиву (пегости), в силу чего меняются размеры непигментных участков.Такой тип наследования признаков встречается очень часто. Число степеней Свободы |
Уровень достоверности | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5% | 1% | 0,1% | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 | 3,84 | 6,64 | 10,83 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 5,99 | 9,21 | 13,82 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | 67,82 | 11,34 | 16,27 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 и т.д. до 10 | 9,49 | 13,28 | 18,47 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В задаче 1 степень свободы, χ2 = 0,59, а это меньше 5%-го уровня достоверности (3,84), значит расхождение допустимо, эксперимент согласуется с законом Менделя.
Задача 2.
При скрещивании пестрого петуха с пестрой курицей получено 38 цыплят с разной окраской оперения:
| Наблюдаемые значения (Н) | Черные 11 | Пестрые 18 | Белые 9 | Всего: 38 |
| Ожидаемые значения (О) | 38*1/4=9,5 | 38*2/4=19 | 38*1/4=9,5 | 38 |
| Разница Н-О | +1,5 | -1 | -0,5 | |
| (Н-О)2 | 2,25 | 1 | 0,25 | |
 (Н-О)2
О (Н-О)2
О
| 2,25/9,5=0,24 | 1/19=0,05 | 0,25/9,5=0,03 |
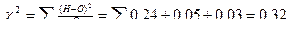
 Необходимо найти число степеней свободы: Ф-1 .У нас 3 фенотипа цыплят (черные, пестрые, белые), следовательно, 3-1=2.
Необходимо найти число степеней свободы: Ф-1 .У нас 3 фенотипа цыплят (черные, пестрые, белые), следовательно, 3-1=2.
Находим по таблице значения χ2 при двух степенях свободы. Для 5%-го уровня достоверности значение составляет 5,99, а это меньше рассчетного - 0,32.
Ответ: расхождение между теоретически ожидаемыми и экспериментально полученными значениями допустимо, отклонения можно считать случайными, а сам эксперимент согласуется с законами Менделя.
Задача 3.
Скрестили дигетерозиготных стрекоз с нормальными крыльями и темной окраской глаз. Во втором поколении наблюдали следующее распределение:
| Наблюдаемые значения (Н) | Норм.крылья, темные глаза: 268 | Корот.крылья, темные глаза: 50 | Норм. крылья, яркие глаза: 54 | Корот.крылья, яркие глаза: 28 | Всего: 400 |
| Ожидаемые значения (О) | 400 * 9/16 = 225 | 400 * 3/16 = 75 | 400 * 3/16 = 75 | 400 * 1/16 = 25 | 400 |
| Разница Н-О | +43 | -25 | -21 | +3 | |
| (Н-О)2 | 1849 | 625 | 441 | 9 | |
 (Н-О)2
О (Н-О)2
О
| 1849/225= 8,22 | 625/75= 8,33 | 441/75= 5,88 | 9/25= 0,36 |
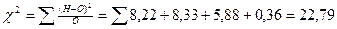
Находим число степеней свободы: 4 фенотипа потомства – 1 = 3.
Смотрим по таблице значений χ2 при 3 степенях свободы: 22,79>7,82>11,34>16,27
Ответ: В данном опыте нельзя говорить о соблюдении закона независимого расщепления.
Питание плодовой мушки
В природе питается подгнившими плодами. В лаборатории питательная смесь должна быть стандартной: манная крупа, сахар (патока), изюм (сухой виноград), агар–агар (склеивающее вещество морских водорослей), вода.
Цикл развития от яйца до взрослой особи:
при температуре 21-22°С – 12 – 14 дней;
при температуре 24-25°С – 10 – 11 дней;
повышение температуры до 31°С приводит к частичной или полной
стерильности; < t° – замедляет рост.
Мух содержат в пробирках с питательной средой в термостатах с освещением при стандартной t и влажности. Посуда должна быть стерильной (2 часа при t +160°С).
I . Мутация глаз
К числу органов дрозофилы, чаще всего подвергающихся мутационным изменениям, относятся глаза. Глаза нормальной мухи устроены по типу сложных фасеточных глаз насекомых. Сложный глаз состоит из многих простых глазков – фасеток. У здоровых мух дикого типа фасетки напоминают по форме пчелиные соты. Суммарное число фасеток у самок = 780, у самцов = 740.
а) У мутантных мух линии Star – звездчатые глаза.Фасетки неправильной формы, волоски растут пучками.
б) У мутантных мух линии glass – глаза стекловидные, их поверхность ровная и гладкая.
в) Многие мутации связаны с изменением пигментации глаза. Например, у мух линии Bar глаза – полосковидные. Мухи линии eyeless – безглазые, у них не только резко уменьшена голова, но и почти нет фасеток, их суммарное число уменьшено до 70 шт.
г) Есть линии с лопастной формой глаза.
Следствием множественного аллелизма является разнообразная окраска глаз:
линия P (pink) – розовые глаза;
линия apr (apricon ) – абрикосовые глаза;
линия bw (brown ) – коричневые глаза;
линия w (white) – белые глаза.
II . Мутации крыла
а) Мутации, связанные с изменением общей конфигурации крыла:
curled– крылья, закрученные к верху;
curved– крылья, закрученные к низу;
notch – вырезка на дистальном конце крыла;
ski – форма лыжи;
arc – арка;
aeroplane – крылья сложены в полете подобно самолету.
б) Группа мутаций, затрагивающих жилкование крыла (на крыле появляется одна или несколько добавочных жилок, что ведет к изменению и общего количества ячеек, расположенных между продольными жилками).
Задание: рассмотреть на представленной схеме продольные жилки и ячейки между ними у нормальной мухи. Сравнить с мутантными линиями коллекционного материала. Обратить внимание на то, что у мух линии plexus жилки разветвленные. Зарисовать в тетрадь.
в) Мутации связанные с изменением размера крыла:
vestigial – зачаточные крылья;
rudimentary – рудиментарные;
apterous – бескрылые формы.
III . Мутации щетинок
Тело мушки покрыто многочисленными щетинками. Щетинки (макрохеты) являются одним из систематических признаков мухи. Мутации связаны либо с исчезновением щетинок на голове, груди, либо (что наблюдается реже) с их удвоение, изменением длинны:
spineless – очень короткие щетинки; есть линия с очень длинными щетинками;
forked – вильчатые щетинки.
Процедура введения вещества
Введение ХВ дрозофиле обычно производится либо путем внутрибрюшной инъекции, либо с кормом на стадии имаго или личинки. Другие способы обработки мух – ингаляционный или опрыскивание с помощью аэрозоля. Для тестирования рекомендуется использовать взрослых самцов, поскольку самки более легко стерилизуются под воздействием ХВ и, как доказано, более устойчивы к индукции наследуемых генетических изменений. Опыты с мухами доказывают, что химические мутагены обладают высокой специфичностью в отношении той или иной стадии развития, т.е. проявляют более или менее выраженное мутагенное действие на различных стадиях развития половых клеток. Весьма важно проанализировать потомство самцов, которые были обработаны ХВ на стадии сперматозоидов, поздних и ранних сперматид, а также сперматоцитов.
При скрещивании на 1 самца необходимо брать избыток в 3-5 самок, что обеспечивает отбор всех половых клеток, находящихся в стадии зрелости (3-4 скрещивания за период в 7-9 дней).
Решение: ав
Мужчина несет два рецессивных признака. Значит, его генотип ===.
ав
Женщина страдает обоими заболеваниями, следовательно, содержит доминантные гены А и В и их рецессивные аллели а и в. Одну болезнь она унаследовала от отца, а другую от матери, значит, доминантные гены расположены не в одной, а в разных гомологичных хромосомах, и генотип
Ав
женщины: == .
аВ
Запись брака
Р: Ав х ав
♀ == ♂ ==
аВ ав
катаракта, полидактилия здор., здор.
Гаметы: Ав, аВ ав
F 1: Ав ав
== 50% == 50%
ав ав
катаракта полидактилия
Ответ: 50% детей будут многопалыми, а 50% будут болеть полидактилией.
Негомологичных хромосом
З а д а ч а 1 .
В семье, где отец имел гипертрихоз (сцеплен с Y-хромосомой), а мать - полидактилию (аутосомно-доминантный признак), родилась нормальная в отношении двух признаков дочь. Какова вероятность того, что следующий ребенок будет без аномалий, какой пол возможен у ребенка?
З а д а ч а 2.
В браке здоровых родителей родился сын-альбинос (аутосомно-рецессивный признак), страдающий гемофилией. Определите вероятные генотипы родителей, а также оцените вероятность рождения второго сына здоровым?
З а д а ч а 1.
В молекуле и-РНК количество уридиловых нуклеотидов на 10% больше, чем цитидиловых. Определите количество всех нуклеотидов в молекуле ДНК, с которой получена эта и-РНК.
Ответ: Известно, что гуаниловый нуклеотид комплементарен цитидиловому, адениловый комплементарен уридиловому и число этих нуклеиновых оснований в комплементарных парах в молекуле ДНК в целом, в соответствии с правилами Чаргаффа, равно друг другу (А=Т; Г=Ц). Обозначим общее число цитидиловых нуклеотидов через х%, число уридиловых через х+10%, в этом случае число гуаниловых составит х, количество адениловых х+10%. Составляем уравнение: 4х+20%=100%; х=20%.
З а д а ч а 2.
Дан полипептид, состоящий из 10 аминокислот: мет-вал-сер-три-глу-тре-лиз-асп-тре-тир. Постройте цепь молекулы и-РНК и комплементарную ей цепь молекулы ДНК, воспользовавшись таблицей генетического кода. Рассмотрите различные варианты.
Таблица 1.
Генетический код
| Аминокислота | Смысловые кодоны и-РНК | Примечания |
| Фенилаланин | УУУ, УУЦ |
В митохондриях позвоночных кодоны АГА и АГГ выполняют функцию нонсенсов. (С.Г.Инге-Вечтомов, 1996).
З а д а ч а 3.
Фрагмент молекулы белка миоглобина содержит аминокислоты, расположенные в следующем порядке: вал-ала-глу-тир-сер-глн. Напишите структуру участка молекулы ДНК, кодирующего эту последовательность аминокислот.
Ответ: Исходными являются данные о последовательности аминокислот в молекуле миоглобина.
1. По таблице 1 находим триплеты, кодирующие каждую из указанных аминокислот (в таблице указаны кодоны и-РНК, а не ДНК поскольку в синтезе белка участвуют непосредственно молекулы и-РНК, а не ДНК). Код для аминокислот: валина - ГУУ, аланина ГЦУ и др. Если аминокислота закодирована несколькими кодонами, то можно выбрать любой из них. Затем выписываем кодоны всех аминокислот в последовательности, соответствующие порядку аминокислот. Полученная цепочка отражает строение молекулы и-РНК: ГУУ-ГЦУ-ГАА-УАУ-УЦУ-ГАА.
2. Определяем строение той цепочки ДНК, которая кодировала строение и-РНК. Для этого под каждым кодоном молекулы и-РНК записываем комплементарный ему кодон молекулы ДНК 3. ДНК состоит из двух цепочек, поэтому под кодонами первой цепочки ДНК записываем кодоны второй цепочки, образованные по принципу комплементарности.
и-РНК:ГУУ - ГЦУ - ГАА - УАУ - УЦУ- ЦАА
ДНК: ЦАА – ЦГА –ЦТТ – АТА – АГА – ГТТ
ГТТ – ГЦТ – ГАА - ТАТ - ТЦТ – ЦАА
З а д а ч а 4.
Полипептидная цепь А-инсулина включает 20 аминокислот: глицин-изолейцин-валин-глутамин-глицин-цистеин-цистеин-серин-валин-цистеин-серин-лейцин-тирозин-глицин-лейцин-глутамин-аспарагин-тирозин-цистеин-аспарагин. Определите структуру участка молекулы ДНК, кодирующего эту полипептидную цепь.
Таблица 2 . Код ДНК (основания – ДНК)
Первое основание
Второе основание
Третье основание
По таблице 2 находим триплеты, кодирующие аминокислоты в полипептидной цели инсулина. Первая цепочка:
ЦЦА - ТАА - ЦАА - ГТТ - ЦЦА - АЦА - АЦГ - АГА - ЦАА - АЦА - ТЦА - ГАА - АТА - ЦЦА - ГАА - ГТТ - ТТА - АТА – АЦА- ТТА.
Состав ДНК можно определить по таблице 1, если сначала составить цепочку и-РНК, а затем ДНК по принципу комплементарности. Вторую цепочку ДНК достройте самостоятельно.
Аудиторная работа
Цель. Уметь составлять родословную для последующего ее анализа.
Оборудование. Таблицы: условные обозначения для составления родословных; родословные, отражающие разные типы наследования.
Оценка генеалогического анамнеза (ГА)
Генеалогический (наследственный) анамнез собирается методом составления родословной семьи ребенка. Лицо, для которого строится родословная (в данном случае - студент), называется пробандом. Родословная должна содержать сведения не менее трех поколений, включая самого пробанда.
Основные цели исследования:
1. Выявление наличия в родословной пробанда наследственных заболеваний (болезнь Дауна, фенилкетонурия, муковисцедоз, целиакия, лактозная недостаточность, глухонемота и др.).
2. Определение отягощенности генеалогического анамнеза - индекса отягощенности (J). Для этого необходимо сосчитать общее число длительно текущих, хронических заболеваний и врожденных пороков развития у всех родственников и членов семьи (исключая пробанда) и соотнести его с числом самих родственников,сведения о которых брались во внимание. А
J = — , где
Б
J - индекс отягощенности наследственного анамнеза;
А - общее число длительно текущих, хронических заболеваний и врожденных заболеваний у всех родственников, о которых есть сведения (пробанд не учитывается);
Б - общее число родственников, кроме пробанда, в плане которых есть сведения о состоянии здоровья.
Наследственный анамнез считается:
· благополучным - при числовом выражении индекса J от 0,1 до 0,3;
· условно благополучным - при J от 0,4 до 0,7;
· неблагополучным - при J более 0,7.
3. Установление направленности отягощенности:
- если в поколениях родословной выявляются хронические болезни одних и тех же органов и систем - это свидетельствует об однонаправленности отягощенности;
- если в поколениях родословной выявляются хронические заболевания разных органов и систем - отягощенность мультифакториальная.
Примеры оценки генеалогического анамнеза
Пример I .
 |  |
дед баб. дед баб. С
1 2 3 4

 ----------------------------------------------------------
----------------------------------------------------------
 |  |
отец мать тетя В
1 2 3
----------------------------------------------------------


 1 2 3 А
1 2 3 А
пробанд аборт брат пробанда
С1 — дед, 65 лет, здоров;
С2 — бабушка, 60 лет, сахарный диабет;
С3 — дед, 58 лет, хроническая пневмония;
С4 — бабушка, 57 лет, здорова;
В1 — отец, 37 лет, здоров;
В2 — мать, 36 лет, здорова;
В3 — тетя; 20 лет, врожденный порок сердца;
А1 — пробанд, 16 лет;
А2 — медицинский аборт;
А3 — брат пробанда, 11 лет, здоров.
3
Оценка ГА: J = — = 0,37 (анамнез условно благополучный).
8
 Пример 2.
Пример 2.
 | |  |
дед баб. дед баб. С
1 2 3 4
--------------------------------------------------------
 |  |
тетя отец мать В
1 2 3
 --------------------------------------------------------
--------------------------------------------------------

пробанд сестра А
1 2
С1 — дед, 62 года, умер, рак поджелудочной железы;
С2 — бабушка, 60 лет, здорова;
С3 — дед, 69 лет, хронический гепатит, гипертоническая болезнь;
С4 — бабушка, 65 лет, ишемическая болезнь сердца;
В1 — тетя, 40 лет, желчно-каменная болезнь, гипертоническая болезнь;
В2 — отец, 39 лет, язва 12-перстной кишки;
В3 — мать, 38 лет, здорова;
А1 — пробанд, 17 лет;
А2 — сестра, 15 лет, пищевая аллергия, хронический тонзиллит.
9
Оценка ГА: J = — = 1,1 (анамнез неблагополучный).
8
Направленность отягощенности: возможность желудочно-кишечных
заболеваний.
Пальмоскопия
Кроме папиллярных узоров на ладонной поверхности и стопе хорошо видны различные линии, они также изучаются и называются «сгибательными складками».
Задание:
- Раскрыв ладонь, поработайте большим пальцем. Сгибательная складка большого пальца (1) охватит полукругом подушечку у его основания.
- Попробуйте согнуть три пальца (кроме большого и указательного), на ладони образуется трехпальцевая складка (2).
- При движении всех пальцев кисти к центру, образуется единая пятипальцевая складка (3).
Рассмотрите на рисунках как выглядит топографическая схема ладони человека, не имеющего наследственной патологии. При генных и хромосомных аномалиях, еще во внутриутробном периоде, на ладони образуется четырехпальцевая поперечная линия (ЧПЛ), при этом трех- и пятипальцевые борозды отсутствуют.
Наследственные заболевания, при которых выявляется ЧПЛ:
- Брахидактилия (короткопалость) - аутосомно-доминантное заболевание, связано с недоразвитием дистальных (ногтевых) фаланг пальцев.
- Анонихия - аутосомно-доминантный характер наследования, недоразвитие ногтей, аномалия развития стоп и кистей, отсутствие одного или несколько пальцев.
- Нейрофиброматоз - аутосомно-доминантное злокачественное заболевание, характеризуется множественным образованием опухолей нервных стволов. Опухоли локализуются в любых органах и тканях, в ЦНС, но чаще на коже в виде пигментированных бородавок с избыточным ростом волос. Наблюдается отставание физического и умственного развития. Болезнь обнаруживает себя или сразу после рождения или в подростковом возрасте.
- Синдром Патау - дополнительная хромосома в 13-15 паре аутосомных хромосом. Множественные пороки головного мозга (недоразвитие лобных долей, мозжечка, третьего желудочка мозга), патология сердечно - сосудистой системы, почек, патология зрения. Все это приводит к ранней смерти (на 3-4 мес. жизни). Синдром Патау чаще всего обнаруживается у девочек (1 случай на 4 тыс. родившихся)
- Синдром Эдвардса - трисомия 18-й хромосомной пары. Множественные дефекты жизненно важных органов: головного мозга, мозжечка, сердца, легких, почек. До 1 мес. доживают 70% больных, до 1 года - только 7%, до 10 лет - 1% детей. Синдром чаще встречается у девочек.
- Синдром Дауна (1 случай на 600 новорожденных) - трисомия по 21 хромосомной паре. Множественные пороки внутренних органов, сердечно - сосудистой, пищеварительной, половой систем. Сопротивляемость к инфекциям снижена. Умственная отсталость (идиотия), деформация черепа. Продолжительность жизни небольшая. У лиц мужского пола - бесплодие, у женщин - 1/2 часть детей - с патологией.
Таким образом, выявлена корреляция между уклонением папиллярного рисунка с серьезными наследственными заболеваниями. Доля риска существует, если один из родителей, даже здоровый имеет на ладони ЧПЛ. Специалисты изучают даже небольшие патологические уклонения рисунка (стигмы), которые свидетельствуют о предрасположенности к той или иной патологии, заранее разъясняя лицам "группы риска", чего следует избегать, чтобы болезнь не развилась. У каждого человека своя уникальная гравюра, к чему бы мы не прикасались, - всюду остаются отпечатки наших пальцев. В музее антропологии собраны десятки тысяч отпечатков людей нашей планеты.
Система групп крови АВ0
В 1900 г. К. Ландштейнером была открыта первая АВО-система групп крови. Годом позже было показано, что группы крови наследуются. В 1924 г. Ф. Берпцтейн установил, что система групп крови АВО определяется тремя аллелями (А, В, О) одного и того же гена, т. е. эта система групп крови наследуется по типу множественных аллелей. В различных сочетаниях генов образуются четыре группы крови: I группа - ОО, II группа –АА, АО, III группа – ВВ, ВО, IV группа - АВ.
Любой диплоидньй организм имеет две гомологичные хромосомы и, соответственно, не более двух аллелей одного гена. Аллели А и В являются доминантными по отношению к аллелю О. Аллели А и В не подавляют друг друга, их действие проявляется независимо при образовании IV группы крови. Это явление получило название кодоминирования.
По данным ряда исследователей, среди европейского населения наиболее часто встречаются люди с I группой крови, реже с IV группой.
Распространение аллелей групп крови АВО в различных странах мира (%)
| Популяция | 0 | А | В | АВ |
| Русские (Москва) | 33,3 | 37,4 | 22,8 | 6,5 |
| Французы | 41,6 | 47,0 | 8,0 | 3.3 |
| Китайцы | 28.6 | 26.6 | 32.0 | 12.8 |
| Японцы | 30.1 | 38.4 | 21.9 | 9.7 |
| Австралийцы | 60.7 | 39.3 | 0 | 0 |
| Бразильцы (Бороро) | 100,0 | 0 | 0 | 0 |
Различия крови человека по системе АВО основаны на комбинациях двух изоантигенов (А и В) в эритроцитах и двух антител (a и b) в плазме крови. Эти различия необходимо учитывать при переливании крови.
Резус-фактор
В 1940 г. К. Ландштейнер и А. Винер обнаружили антиген, содержащийся в крови человека и макаки-резус, — резус-фактор (Rh). В простейшем случае резус-положительность (Rh) доминирует над резус-отрицательностью (rh). Известно, что 85% людей являются резус-положительными и 15% — резус-отрицательными.
Установлено, что резус-фактор определяется рядом тесно сцепленных генов, что обусловливает большое разнообразие вариантов резус-несовместимости. Кроме того, показано, что несовместимость по резус-фактору матери и плода является причиной иммунного конфликта, приводящего к эритробластозу плода и гемолитической болезни новорожденных. Иммунный конфликт возникает у резус-отрицательной матери (rhrh) и резус-положительного плода (Rhrh).
В период беременности у матери начинают вырабатываться Rh-антитела, которые губительно действуют на резус-положительный белок крови плода. Особенно опасны вторые и последующие роды, когда антитела в крови матери накапливаются в большом количестве. Это приводит к рождению детей, страдающих анемией, гемолитической болезнью (желтухой) и другими патологиями.
Основной способ лечения эритробластоза — удаление антител из крови больного ребенка и замещение крови (переливание крови сразу после рождения).
Тема 17. БИОХИМИЧЕСКИЙ СКРИНИНГ БОЛЕЗНЕЙ ОБМЕНА ВЕЩЕСТВ
Существует группа заболеваний под общим названием – ферментопатии (или энзимопатии). Заболевания данной группы развиваются вследствие полного отсутствия того или иного фермента или снижения его активности. Ферменты (энзимы) являются специфическими катализаторами белковой природы, способные во много раз ускорять протекающие в живых организмах реакции. Благодаря ферментам все химические реакции упорядочены, строго согласованы, образуют единую систему, которую мы называем обменом веществ и энергии.
Энзимопатии подразделяются на наследственные и приобретенные. Наследственные - связаны с генетически обусловленной недостаточностью одного или нескольких ферментов. Первоначально заболевание носит скрытую форму, то есть выраженных клинических симптомов нет. Установить диагноз «энзимопатия» можно на основании биохимических исследований крови, мочи, реже кала или слюны. В идеале такие исследования должны проводиться в роддоме, так как ферментативный дефект способствует накоплению в организме промежуточных продуктов обмена веществ, которые повышая свою концентрацию, нарушают функции определенных органов. И лишь тогда появляются первые клинические симптомы болезни. Иногда это может обнаруживаться в подростковом возрасте.
Ферменты, в зависимости от типа катализируемой реакции, подразделяются на 6 основных классов. По этому же принципу выделяют и группы наследственных энзимопатий (смотря какой обмен веществ нарушается).
Гистидинемия. Отсутствует фермент, расщепляющий аминокислоту гистидин. Это приводит к нарушениям функций ЦНС, результатом чего являются: слабоумие, изменения мышечного тонуса, в силу чего у больных отмечаются частые судороги и др.
Конкордантность некоторых признаков человека у однояйцовых (ОБ) и двуяйцовых (ДБ) близнецов
| Признаки | Конкордантность, % | |
| ОБ | ДБ | |
| Нормальные | ||
| Группа крови | 100 | 46 |
| Цвет глаз | 99,5 | 28 |
| Цвет волос | 97 | 23 |
| Папиллярные узоры | 92 | 40 |
| Патологические | ||
| Косолапость | 32 | 3 |
| «Заячья губа» | 33 | 5 |
| Врожденный вывих бедра | 41 | 3 |
| Паралитический полиомиелит | 36 | 6 |
| Бронхиальная астма | 19 | 4,8 |
| Корь | 98 | 94 |
| Эпидемический паротит | 82 | 74 |
| Туберкулез | 37 | 15 |
| Дифтерия | 50 | 38 |
| Эпилепсия | 67 | 3 |
| Шизофрения | 70 | 13 |
| Гипертония | 70 | 13 |
| Ревматизм | 20,3 | 6,1 |
Если значение коэффициента конкордантности примерно близко у монозиготных и дизиготных близнецов, то считают, что развитие признака определяется, главным образом, негенетическими факторами, т. е. условиями среды.
Если в развитии изучаемого признака участвуют как генетические, так и негенетические факторы, то у монозиготных близнецов будут иметь место определенные внутрипарные различия. При этом будут уменьшаться различия между моно- и дизиготными близнецами по степени конкордантности. В этом случае считают, что к развитию признака имеется наследственная предрасположенность.
Для количественной оценки роли наследственности и среды в развитии того или иного признака используют различные формулы. Чаще всего пользуются коэффициентом наследуемости, который вычисляется по формулам:
Н = КМБ - КДБ : 100 - КДБ (в процентах);
Н = КМБ - КДБ : 1 - КДБ (в долях единицы), где
Н — коэффициент наследуемости,
К — коэффициент парной конкордантности
в группе монозйготных или дизиготных близнецов.
В зависимости от значения Н судят о влиянии генетических и средовых факторов на развитие признака. Например, если значение Н близко к нулю, считают, что развитие признака обусловлено только факторами внешней среды. При значении Н от 1 до 0,7 — наследственные факторы имеют доминирующее значение в развитии признака или болезни; среднее значение Н от 0,4 до 0,7 свидетельствует о том, что признак развивается под действием факторов внешней среды при наличии генетической предрасположенности.
Рассмотрим несколько примеров. Как уже было отмечено, группы крови у человека полностью обусловлены генотипом и не изменяются под влиянием среды. Коэффициент наследуемости равен 100%. По некоторым морфологическим признакам (форме носа, бровей, губ и ушей, цветуглаз, волос и кожи) монозиготные близнецы конкордантны в 97—100%, а дизиготные (в зависимости от признака) — в 70—20% случаев. Конкордантность МБ по заболеваемости шизофренией равна 70%, а у ДБ — 13%. Тогда Н - (70 - 13): (100 -13) = 0,65 или 65%. В данном случае преобладают генетические факторы, но существенную роль играют и условия среды.
С помощью близнецового метода выявлено значение генотипа и среды в патогенезе многих инфекционных болезней. Так, при заболевании корью и коклюшем ведущее значение имеют инфекционные факторы, а при туберкулезной инфекции существенное влияние оказывает генотип. Исследования, проводимые на близнецах, помогут ответить на такие вопросы, как влияние наследственных и средовых факторов на продолжительность жизни человека, развитие одаренности, чувствительность к лекарственным препаратам и др.
В настоящее время близнецовый метод в генетике человека используется в сочетании с другими методами генетического анализа.
I. Группировка данных
Цель занятия. Освоение техники построения вариационного ряда в малых и больших выборках. Построение сгруппированного вариационного ряда и его графическое изображение.
Методические указания. Систематизация первичных данных называется группировкой. Чаще всего данные группируют в вариационные ряды (в малых выборках варианты располагаются от min значения к max):
1,0 1,1 1,2 1,3 1,4 1,5
Техника построения вариационного ряда в больших выборках несколько иная:
1. Определяют объем выборки: п = 50 (70, 80 и т.д.).
2. Находят min и max лимиты и разницу между ними: Vmax – Vmin .
3. Выбирают число классов, пользуясь ниже приведенной таблицей.
Критерий хи-квадрат
Данный показатель используют при сравнении эмпирических (опытных) рядов с теоретическими: по распределению частот в гибридологическом анализе, при сравнении двух эмпирических рядов, при оценке эффективности применения биологически активных соединений и др.
Пример. При скрещивании самок дрозофилы с серым телом с темнотелыми самцами в F 1 получили всех серотелых мух, а в F 2 произошло расщепление: из 1584 мух 1199 оказались серыми, а 385 были темнотелыми. Соответствует ли наблюдаемое расщепление теоретически ожидаемому 3:1?
Вычисление критерия х2 (хи-квадрат)
| Классы | Наблюдаемые данные (О) | Ожидаемые данные (Е) | О – Е | (О – Е)2 | (О – Е)2 Е |
| серотелые | 1199 | 1188 | +11 | 121 | 0,10 |
| Темнотелые | 385 | 396 | -11 | 121 | 0,31 |
Σ = 1584 Σ = 1584 Σ= 0,41
Вычисленное значение х2= 0,41 сравнивают со стандартным значением х2.
Стандартные значения х2
Числа степеней свободы
Уровни значимости, %
Для вхождения в таблицу необходимо определить число степеней свободы (ν). В гибридологических задачах на расщепление оно всегда равно числу фенотипических классов без единицы. В нашем примере ν = 2 – 1 = 1.
В строке со степенью свободы равной 1 мы видим три значения: 3,84; 6,63; 10,83, которым соответствуют вероятности 0,05; 0,01 и 0,001. В таблице указаны вероятности нулевой гипотезы. Если вычисленное значение х2 ≤ х2 st при Р = 0,05, гипотезу принимают. х2 = 0,41< 3,84, следовательно, нулевую гипотезу можно принять, а наблюдаемые отклонения от теоретически ожидаемого результата следует отнести в разряд случайных. Если х2 > х2st при Р < 0,01 или Р < 0,001, то отклонения не случайные.
Литература:
1. А.М.Крюков, Н.В.Горбунова – Задачник по генетике животных.// Учебно-метод.пособие для самостоятельной работы студентов. // ФГОУ ВПО ПГСА, Пенза, 2005, 184С.
2. Н.В.Анисимова – Популяционная генетика. // Методическое пособие. Пенза, ПГПУ, 2007.
Вариант тестирования на знание исторических дат, связанных с выдающимися событиями в области генетики:
· 1865 г. - Открытие Г.Менделем основных законов наследственности. Доклад среди членов Общества естествоиспытателей.
· 1865 г. – Создание Френсисом Гальтоном генеалогического метода составления родословных человека.
· 1871 г. – Открытие нуклеиновых кислот.
· 1880 г. – О.Гертвиг обнаружил слияние ядра мужской гаметы с ядром женской гаметы у иглокожих, затем у голосемянных и покрытосемянных растений.
· 1882 г. – Вальтер Флеминг, изучая клетки ящериц под микроскопом, обнаружил в них маленькие «ниточки», которые делились на части. Позднее оказалось, что это хромосомы.
· 1888 г. – Способные к делению ядерные элементы клетки В.Вальдейер назвал «хромосомами».
· 1889 г. – Теодор Бовери установил факт редукции числа хромосом в половых клетках вдвое.
· 1892 г. – Замена Флеммингом ранее используемого термина «кариокинез» новым - «митоз», с расшифровкой 3-х основных стадий: профазы, метафазы, анафазы.
· 1894 г. – М.Гейденгайн назвал заключительную стадию деления митозом «телофазой».
· 1900 г. - Официальная дата принятия нового научного направления, связанного с изучением механизмов передачи наследственной информации живых организмов. Переоткрытие законов Г.Менделя.
· 1906 г. – Вильям Бэтсон придумывает официальное название «генетика», экспериментально доказывает универсальность Менделевских законов на животных объектах (наследование формы гребня у кур).
· 1909 г. – Иогансен заменяет термин «наследственные задатки» более лаконичным – «ген».
· 1910 г. - Томас Хант Морган, изучая фруктовую тлю, плодовую мошку, доказал, что гены, определяющие видовые признаки, локализованы в хромосомах.
· 1913 г. – Т.Морган – линейное расположение генов в хромосомах.
· 1926 г.- Американский биолог Герман Меллер открыл влияние рентгеновских лучей на генетические изменения в организме (мутации).
· 1931 г. – Описание механизма рекомбинации.
· г. – Освальд Эвери, Колин Мак Леод и Маклин МакКарти доказали окончательно, что ДНК, а не белки отвечают за наследственность, являясь генетическим материалом.
· г. – Описание механизма кодирования белков генами.
· 1951 г. – Расшифровка последовательности аминокислот в белке.
· 1953 г – Открытие двойной спирали ДНК Уотсоном и Криком.
· 1957 г. – Описание структуры нуклеотидов и нуклеотидсодержащих коферментов.
· 1963 г. – Чарльз Яновский из Стенфорда с коллегами доказал, что последовательность ДНК в точности соответствует последовательности аминокислот в белке.
· 1972 г. В лаборатории Пола Берга (США) создана первая рекомбинантная ДНК. Официальное признание биотехнологии как нового научного направления.
· 1973 г. – Американские биохимики Стэнли Коэн и Герберт Бойер впервые встроили ген африканской жабы в ДНК бактерии, при этом ген стал успешно работать.
· 1978 г. – Клонирован ген человеческого инсулина.
· 1980 г. – Человеческий альфа-интерферон, созданный методом биотехнологии.
· 1982 г. – Человеческий инсулин, созданный методами биотехнологии, допущен в аптечную сеть.
· 1984 г. – Алекс Джеффри (Британский университет) разработал систему «генетических отпечатков» ДНК для идентификации личности.
· 1986 г. – Первая вакцина для человека (от гепатита В), полученная методами биотехнологии.
· 1988 г. – Получена генетически измененная мышь (патент).
· 1990 г. – Американский генетик Ф.Андерсон провел первую генетическую операцию на 4-х летней девочке с нарушениями функции имунной системы.
· 1992 г. – Американские и британские ученые разработали технику генетического тестирования эмбриона на наличие генных нарушений.
· 1997 г. – Ян Вилмут (шотландский институт) клонировал овечку Долли.
· 1998 г. – Создание клонов мышей, поколения клонов.
· 1998 г. – В японском университете Кинки созданы 8 одинаковых телят из клеток, взятых от одной взрослой коровы.
ГЕНЕТИКА ПОПУЛЯЦИЙ
Генетика популяций призвана ответить на целый ряд вопросов:
· как реализуются законы Г.Менделя на уровне популяций;
· как влияют на генетическую структуру популяций такие факторы, как мутационный процесс, отбор, миграции, случайное изменение генных частот.
Знание популяционной генетики необходимо для понимания эпидемиологии наследственных болезней животных и человека, для планирования мероприятий по предупреждению неблагоприятного воздействия на генетический аппарат факторов окружающей среды. Еще одна сфера приложения популяционно-генетических исследований - теория эволюции. Теоретические основы популяционной генетики подробно разработаны в работах Р.А.Фишера, Дж.Б.С.Холдейна, С.Райта, Ф.Айала, Дж.Кайгера, Ф.Фогеля, А.Мотульски.
По мнению специалистов исследования в области популяционной генетики условно можно разделить на две основные группы:
Оба эти подхода тесно взаимосвязаны между собой.
Основная
1. Инге-Вечтомов С.Г. Генетика с основами селекции./ М.:Высшая школа, 1989.
2. Лобашев М.Е. Генетика с основами селекции. М.,1979.
3. Хелевин Н.В., Лобанов А.М., Колесова О.Ф. Задачник по общей и медицинской генетике./ М.: Высшая школа, 1987.
4. Айала Ф., Кайгер Дж. Современная генетика /Т.1-3. М.:Мир, 1987.
5. Льюин Б. Гены. / М.:Мир, 1987.
6. Сингер М., Берг П. Гены и геномы. /Т.1-2, М.:Мир, 1998.
7. Фогель Ф., Мотульски А. Генетика человека. / М.:Мир, 1989.
8. Албертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки./ М.: Мир, 1-4., 1994.
9. Георгиев Г.П. Гены высших организмов и их репрессия./ М.:Наука, 1989.
10. Вудо Р. Биохимическая генетика./ М.Мир, 1982.
11. Айала Ф. Введение в популяционную генетику./ М.Мир, 1984.
Дополнительная
1. Гилберт С. Биология развития./ М.: Мир, 1,2 том, 1994.
2. Грин Н., Стаут У., Тейлор Д. Биология./ М.: Мир, 1993.
3. Докинз Р. Эгоистический ген./ М.: Мир, 1993.
4. Карузина И.П. Учебное пособие по основам генетики./ М.:Медицина, 1980.
5. Левонтин Р. Человеческая индивидуальность: наследственность и среда./ М.: Прогресс, 1993.
6. Маркерт К., Уршпруг Г. Генетика развития./ М.: Мир, 1973.
7. Приходченко Н.Н., Шкурат Т.П. Основы генетики человека./ Ростов-на Дону: Феникс, 1997.
8. Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция./ М.: Мир, 1986.
9. Сингер М., Берг П. Гены и геномы./ М.: Мир. 1,2 том, 1998.
10. Тихомирова М.М. Генетический анализ./ М.: Изд-во Ленингр.ун-та, 1990.
11. Фогель Ф., Мотульски А. Генетика человека./ М.: Мир, в 3 т.,1989.
12. Эфроимсон В.П. Введение в медицинскую генетику./ М.: Мед.литература, 1964.
13. Муртазин Г.М. Задачи и упражнения по общей биологии./ М.: Просвещение, 1981.
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
Пензенский государственный педагогический университет
имени В.Г. Белинского
Кафедра анатомии, физиологии и гигиены человека
УЧЕБНО-МЕТОДИЧЕСКИЙ КОМПЛЕКС ПО ДИСЦИПЛИНЕ
Генетика
Химия
Биология
Пенза - 2007 г.
Тексты лекций
Последнее изменение этой страницы: 2019-03-31; Просмотров: 300; Нарушение авторского права страницы
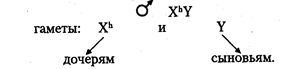
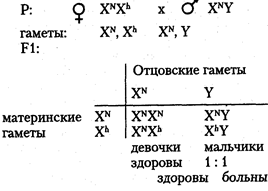

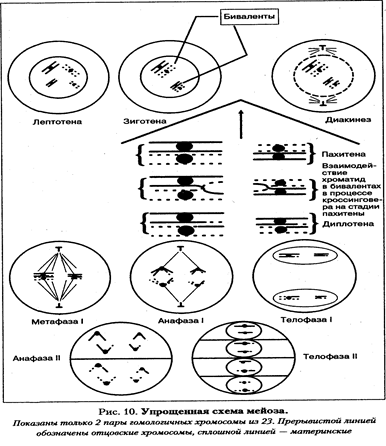
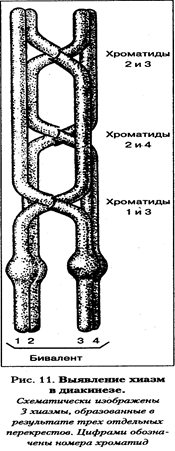
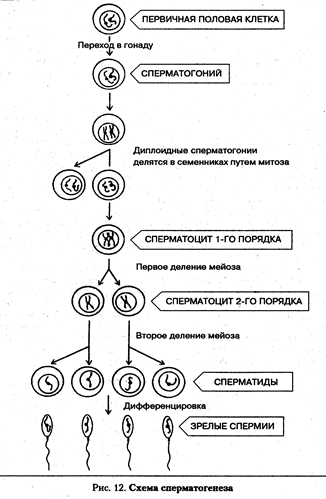
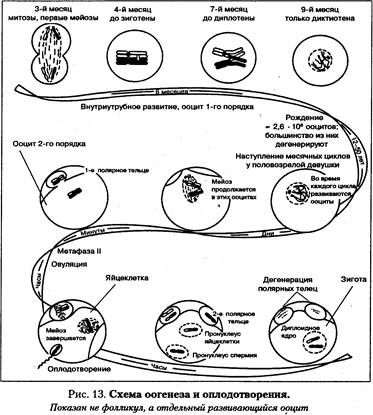
 эффект гетерозиса, гетерозисные животные отличаются скороспелостью и повышенной мясной продуктивностью. Например, при скрещивании двух мясных пород кур получают гетерозисных бройлерных кур, при скрещивании беркширской и дюрокджерсейской пород свиней получают скороспелых свиней с большой массой и хорошим качеством мяса и сала.
эффект гетерозиса, гетерозисные животные отличаются скороспелостью и повышенной мясной продуктивностью. Например, при скрещивании двух мясных пород кур получают гетерозисных бройлерных кур, при скрещивании беркширской и дюрокджерсейской пород свиней получают скороспелых свиней с большой массой и хорошим качеством мяса и сала.














 простой доминантный,
простой доминантный, 
 Различают несколько форм эпистаза: простой рецессивный,
Различают несколько форм эпистаза: простой рецессивный,  АВ ав АВ фиол. фиол. фиол. фиол. 9:7
АВ ав АВ фиол. фиол. фиол. фиол. 9:7  кумулятивную,
кумулятивную,  Выделяют 2 формы полимерии некумулятивную.
Выделяют 2 формы полимерии некумулятивную.