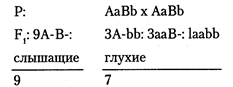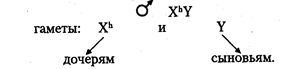|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Т ема 4. ПОЛИГЕННОЕ НАСЛЕДОВАНИЕ СЛОЖНЫХ ПРИЗНАКОВ. ТИПЫ ВЗАИМОДЕЙСТВИЕ ГЕНОВ
Задачи :
Зачастую признаки формируются при участии нескольких генов, взаимодействие между которыми отражается на проявлении фенотипа. Из классической генетики известно, что наиболее изученными являются три типа взаимодействия неаллельных генов: эпистаз, комплементарность и полимерия. Эти типы взаимодействия генов определяют многие наследуемые признаки и у человека. Эпистаз — это такой тип взаимодействия неаллельных генов, при котором одна пара аллельных генов подавляет действие другой пары. Различают эпистаз доминантный и рецессивный. Доминантный эпистаз проявляется в том, что доминантный аллель в гомози-готном (АА) или гетерозиготном (Аа) состоянии подавляет проявление другой пары аллелей. При рецессивном эпистазе ингибирующий ген в рецессивном гомозиготном состоянии (аа) не дает возможность проявиться эпистатируемому гену. Подавляющий ген называют геном-супрессором или ингибитором, а подавляемый — гипостатическим. Этот тип взаимодействия наиболее характерен для генов, участвующих в регуляции онтогенеза и иммунных систем человека. Примером рецессивного эпистаза у человека может служить «бомбейский» феномен. В Индии была описана семья, в которой родители имели вторую (АО) и первую (00) группы крови, а их дети — четвертую (АВ) и первую (00). Чтобы ребенок в этой семье имел группу крови АВ, мать должна иметь группу крови В, но никак ни О. Позже было выяснено, что в системе групп крови АВО имеются рецессивные гены-модификаторы, которые в гомозиготном состоянии подавляют экспрессию антигенов на поверхности эритроцитов. Например, человек с третьей группой крови на поверхности эритроцитов должен иметь антиген группы В, но эпис-татирующий ген-супрессор в рецессивном гомозиготном состоянии h/h подавляет действие гена В, соответствующие антигены не образуются, и фенотипически проявляется группа крови 0. Гены-суп-рессоры наследуются независимо от генов, определяющих группы крови АВО. Комплементарность — это тип взаимодействия генов, при котором за признак отвечает несколько неаллельных генов. Разное сочетание доминантных и рецессивных аллелей в парах приводит к разному фенотипическому проявлению признака. Но во всех случаях, когда гены расположены в разных парах хромосом, в основе расщеплений лежат цифровые законы, установленные Г. Менделем. Чтобы человек имел нормальный слух, необходима согласованная деятельность нескольких пар генов, каждый из которых может быть представлен доминантными или рецессивными аллелями. Нормальный слух развивается только в том случае, если каждый из этих пар генов имеет хотя бы один доминантный аллель в диплоидном наборе хромосом. Если хотя бы одна пара аллелей представлена рецессивной гомозиготой, то человек будет глухим. Рассмотрим этот тип взаимодействия генов на простом примере. Предположим, что нормальный слух формирует две пары генов. В этом случае люди с нормальным слухом могут иметь генотип: ААВВ, ААВb, АаВВ, АаВb. Люди с наследственной глухотой должны иметь генотип: aabb, Aabb, Aabb, aaBb, aaBB. Используя законы Менделя для дигибридного скрещивания, легко рассчитать, что глухие родители (aaBB x AAbb) могут иметь детей с нормальным слухом (AaBb), а родители с нормальным слухом при соответствующем генотипе (AaBb x AaBb) с высокой долей вероятности (более 40%) могут иметь глухих детей:
Полимерия — обусловленность определенного признака несколькими парами неаллельных генов, обладающих одинаковым действием. Такие гены называются полимерными. При кумулятивной полимерии число доминантных аллелей влияет на степень выраженности признака. Чем больше доминантных аллелей, тем более интенсивно выражен признак. По типу кумулятивной полимерии обычно наследуются признаки, которые можно выразить количественно (цвет кожи и волос, рост). Цвет кожи и волос человека обеспечивается пигментом меланином. Все цвета волос, за исключением рыжих, составляют непрерывный ряд от темного до светлого и наследуются полигенно по типу кумулятивной полимерии. Считается, что различия в цвете волос обусловлены чисто количественными изменениями в содержании пигмента. Цвет рыжих волос определяется наличием пигмента феумеланина. По типу кумулятивной полимерии наследуется пигментация кожи человека. На основе генетических исследований семей, члены которых имеют разную интенсивность пигментации кожи, предположили, что цвет кожи человека определяется тремя или четырьмя парами генов. Полимерные гены, как правило, обозначаются одинаковыми буквами, чтобы подчеркнуть однонаправленность их действия. При формировании признака не важно, какой паре генов принадлежат доминантные аллели, важно их количество. В таком случае генотип негра условно можно записать как АААААА, а генотип белого — как аааааа. Светлокожие негры будут иметь генотип АААААа или АаАААА или другие сочетания А и а. Мулаты должны соответствовать генотипу АаАаАа, чем больше а, тем светлее кожа мулата. Если упростить задачу и генотип, определяющий цвет кожи человека, обозначить не тремя парами генов, а двумя АаАа, то легко убедиться, что в браке людей с противоположным цветом кожи все дети будут мулатами: Р: АААА х аааа F: АаАа — мулаты В браке двух мулатов (АаАа х АаАа) существует вероятность рождения детей с разным цветом кожи: — негры (АААА), 1/16; — светлокожие негры (АААа), 4/16; — мулаты (АаАа), 6/16; — светлокожие мулаты (Аааа), 4/16; — белые (аааа), 1/16. Таким образом, с определенной долей вероятности родители могут иметь детей с более светлой или более темной, чем у них, кожей, более светлыми или более темными волосами. Плейотропия. Признание принципа взаимодействия генов наводит на мысль о том, что все гены так или иначе взаимосвязаны в своем действии. Один ген оказывает влияние на работу других, следовательно, он может оказывать влияние на проявление не только одного, но и нескольких признаков. Такое множественное действие гена называют плейотропией. Наиболее ярким примером плейотропного действия гена у человека является синдром Марфана. Симптомами его являются удлиненные пальцы, высокий рост, гиперподвижность суставов, одна из форм близорукости и аневризм аорты. Многие наследственные патологии обладают плейотропным действием. Гены обеспечивают определенные этапы метаболизма. Продукты метаболических реакций в свою очередь регулируют, а возможно, и контролируют другие метаболические реакции. Нарушения метаболизма на одном этапе отразятся на последующих этапах. В результате нарушение экспрессии одного гена окажет влияние на несколько элементарных признаков.
Т ема 5. СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ ГЕНОВ Задачи: 1. Сформировать представление о сцепленном наследовании, группах сцепления, генетическом картировании. 2. Научить объяснять причины сцепленного наследования генов, а также нарушения сцепления между ними (в профазе первого мейотического деления). 3. Показать, что генетическое картирование позволяет установить истинное местоположение (локализацию) отдельных генов в хромосоме с целью дальнейшего воздействия на материальную основу наследственности. Группы сцепления. Хромосомная теория наследственности бьша сформулирована и экспериментально доказана Т. Морганом и его сотрудниками. Согласно этой теории, гены находятся в хромосомах и расположены в них линейно. Гены, локализованные в одной хромосоме, называются сцепленными, наследуются вместе и образуют группу сцепления. Количество групп сцепления соответствует числу пар гомологичных хромосом. У человека 46 хромосом: 22 пары аутосом и одна пара половых хромосом (XX или XY), следовательно, у женщины 23 группы сцепления, а у мужчины — 24, так как половые хромосомы мужчины (XY) не полностью гомологичны друг другу. Каждая из половых хромосом мужчины имеет гены, характерные только для Х-хромосомы и только для Y-хромосомы соответственно имеются группы сцепления Х-хромосомы и Y-хромосомы. Теоретически различают два вида сцепления: полное и неполное. При полном сцеплении генов группа сцепления не нарушается, и все гены, локализованные в одной хромосоме, наследуются вместе. Это достаточно редкое явление. У человека оно не описано, а встречается у гетерогаметного пола тутового шелкопряда и дрозофилы. Неполное сцепление генов — наиболее часто встречающееся явление, которое характеризуется нарушением групп сцепления в результате процесса кроссинговера. Кроссинговер. Гены, локализованные в одной хромосоме и образующие группу сцепления, сцеплены не абсолютно. В зиготене профазы первого мейотического деления гомологичные хромосомы сливаются вместе с образованием бивалентов, затем в пахитене происходит Кроссинговер — обмен участками между хроматидами гомологичных хромосом. Кроссинговер является обязательным процессом и происходит в каждой паре гомологичных хромосом. Чем дальше друг от друга расположены гены в хромосоме, тем чаще между ними происходит Кроссинговер. Благодаря ему, возрастает разнообразие гамет с разным сочетанием генов. Например, пара гомологичных хромосом содержит сцепленные гены АВ и ab. В профазе мейоза гомологичные хромосомы конъюгируют и образуют бивалент: АВ//аЬ. Если Кроссинговер меду генами А и В не произойдет, то в результате мейоза образуется два типа некроссоверных гамет: АВ и ab. Если Кроссинговер состоится, то получатся кроссоверные гаметы: Ab и аВ. т.е. группы сцепления изменятся. Чем более удалены друг от друга гены А и В, тем больше возрастает вероятность образования кроссоверных гамет. Если гены расположены на достаточном друг от друга расстоянии в большой хромосоме и между ними в мейозе происходят многочисленные перекресты, то они могут наследоваться независимо. Частота кроссинговера зависит не только от расстояния между генами. Известны и другие факторы, влияющие на рекомбинацию сцепленных генов. Так, для ряда локусов показано, что частота кроссинговера в мейозе возрастает с возрастом. Генетические карты. Открытие кроссинговера позволило Т. Моргану и сотрудникам его школы в первые два десятилетия нашего века разработать принцип построения генетических карт хромосом. Явление сцепления было использовано ими для локализации генов, расположенных в одной хромосоме, и создания генных карт плодовой мушки Drosophila melanogaster. На генетических картах гены располагаются линейно друг за другом на определенном расстоянии, которое определяется в процентах кроссинговера или в морганидах (1% кроссинговера равен одной морганиде). Для построения генетических карт у растений и животных проводят анализирующие скрещивания, в которых достаточно просто рассчитать процент особей, образовавшихся в результате кроссинговера, и построить генетическую карту по трем сцепленным генам. У человека анализ сцепления генов классическими методами невозможен, поскольку невозможны экспериментальные бракосочетания. Для изучения групп сцепления и составления карт хромосом человека используют другие методы. Сцепление генов в некоторых случаях может быть установлено обзором обширных родословных. Статистический анализ позволяет определить параметры сцепления. Компьютерные программы дают возможность определить наиболее вероятные частоты рекомбинаций сцепленных генов на основе анализа большого количества родословньк. Установленные частоты рекомбинаций в группах сцепления позволяют построить генные карты по группам сцепления. Пары генов или группы сцепления аутосомных генов невозможно соотнести с конкретными хромосомами на основе использования только формально-генетического анализа родословных. Для локализации генов используют морфологические маркеры хромосом. Современные методы генетического картирования хромосом развивались на основе новых методов молекулярной биологии, генной инженерии, гибридизации на препаратах хромосом с ДНК-зондами. Данные о сцеплении генов в хромосомах, полученные современными методами картирования, достаточно хорошо согласуются с первоначальными результатами, основанными на методах классической генетики. В последние годы достигнуты большие успехи в выявлении сцепления и в локализации локусов сцепления в определенных хромосомах. Построены цитологические карты генов хромосом человека, в которых для каждого сегмента дифференциально окрашенной хромосомы показано, какие гены там находятся и на каком расстоянии друг от друга. Число локализованных генов быстро растет. В настоящее время изучены все 24 группы сцепления. Согласно международной программе «Геном человека», к 2005 г. будет картировано большинство генов. Основные положения хромосомной теории наследственности: — гены локализованы в хромосомах, расположены линейно вдоль хромосом в определенной последовательности; — каждый ген занимает определенное место (локус) в хромосоме; — в гомологичных хромосомах аллельные гены занимают одно и то же место;
- гены наследственно дискретны; - гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; - число групп сцепления соответствует гаплоидному набору хромосом и постоянно для каждого вида; - сцепление генов может нарушаться в процессе кроссинговера за счет образования рекомбинантных хромосом; - частота кроссинговера зависит прежде всего от расстояния между генами; чем дальше гены друг от друга, тем чаще между ними происходит кроссинговер; - гены относительно стабильны, но могут изменяться в результате мутационного процесса. Т ема 6. ГЕНЕТИКА ПОЛА Задачи :
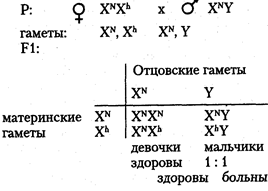
Хромосомный механизм, определяющий пол. В кариотипе человека 46 хромосом (23 пары), из них 22 пары хромосом одинаковы у мужчин и женщин (аутосомы), а по одной паре хромосом, называемой половой, мужчины и женщины отличаются. Половые хромосомы женщины — XX, половые хромосомы мужчины — XY (см. главу 6). Половые хромосомы представлены в каждой соматической клетке индивида. При образовании гамет во время мейоза гомологичные половые хромосомы расходятся в разные половые клетки. Следовательно, каждая яйцеклетка имеет 22 аутосомы и 1 половую хромосому Х (гаплоидный набор хромосом равен 23). Каждый сперматозоид также имеет гаплоидный набор хромосом (22 аутосомы и 1 половая). Половина сперматозоидов содержит Х-хромосому, другая половина — Y-хромосому. Поскольку у женщин половые хромосомы одинаковы и все яйцеклетки несут Х-хромосому, то женский пол у человека называют гомогаметным. У мужчин сперматозоиды различаются по наличию половых хромосом (X или Y), следовательно, мужской пол называют гетерогаметным. Пол человека определяется в момент оплодотворения. Женщина имеет один тип гамет (X) мужчина — два типа гамет (X и Y), причем они образуются в равной пропорции согласно законам мейоза. При оплодотворении хромосомные наборы гамет объединяются. В зиготе 22 пары аутосом и одна пара половых хромосом. Если яйцеклетка оплодотворена сперматозоидом с Х-хромосомой, то в зиготе пара половых хромосом XX. Из такой зиготы развивается девочка. Если яйцеклетка оплодотворена сперматозоидом с Y-xpo-мосомой, то набор половых хромосом в зиготе XY. Такая зигота даст начало мужскому организму. Таким образом, у человека пол будущего ребенка определяет мужчина, так как он гетерогаметен по половым хромосомам. Соотношение полов при рождении по данным статистики соответствует примерно 1:1. Наследование признаков, сцепленных с полом. Хромосомы Х и Y гомологичны, так как у них есть общие гомологичные участки, где локализованы аллельные гены. Однако эти хромосомы, несмотря на гомологию отдельных локусов, различаются по морфологии. Помимо общих участков Х- и Y-хромосомы имеют большой набор генов, по которым они различаются. В Х-хромосоме имеются гены, которые отсутствуют в Y-хромосоме, в свою очередь, в Y-хромосо-ме имеются гены, которых нет в Х-хромосоме. Таким образом, у мужчин в половых хромосомах имеются гены, которые не имеют второго аллеля в гомологичной хромосоме. В этом случае признак определяется не парой аллельных генов, как обычно менделирую-щий признак, а только одним аллелем. Такое состояние гена называется гемизиготным. Признаки, развитие которых обусловлено одним аллелем, расположенным в одной из альтернативных половых хромосом, называются сцепленными с полом. Они развиваются преимущественно у одного из двух полов. Эти признаки наследуются по-разному у лиц мужского и женского пола. Признаки, сцепленные с Х-хромосомой, могут быть и рецессивными, и доминантными. Рецессивные признаки — гемофилия, дальтонизм, атрофия зрительного нерва и миопатия Дюшена. Доминантные — рахит, не поддающийся лечению витамином D, и темная эмаль зубов. Рассмотрим сцепленное с Х-хромосомой наследование на примере рецессивного гена гемофилии. У мужчины половые хромосомы XY. Ген гемофилии локализован в Х-хромосоме и не имеет аллеля в Y-хромосоме, т. е. находится в гемизиготном состоянии. Следовательно, несмотря на то, что признак рецессивный, у мужчин он проявляется. У женщин половые хромосомы XX. Признак определяется парой аллельных генов, поэтому гемофилия проявляется только в гомозиготном состоянии: Основные формальные характеристики Х-сцепленного рецессивного наследования следующие. Обычно поражаются мужчины. Все их фенотипически здоровые дочери являются гетерозиготными носительницами, так как от отца в процессе оплодотворения получают X-хромосому:
Среди сыновей гетерозиготных матерей соотношение пораженных и непораженных 1:1.
Такое наследование получило название крисс-кросс (крест-накрест), т. е. сыновья наследуют фенотипический признак матери, а дочери — отца. Законы передачи признаков, сцепленных с Х-хромосомами, были впервые изучены Т. Морганом. Помимо Х-сцепленных признаков, у мужчин имеются Y-сцепленные признаки, называемые голандрическими. Гены, определяющие голандрические признаки, локализованы в тех районах Y-xpo-мосом, которые не имеют аналогичных участков в Х-хромосоме. Эти признаки также определяются только одним аллелем. Так как голандрические признаки определяются генами, находящимися только в Y-хромосоме, то выявляются они у мужчин и передаются от отца всем сыновьям. Голандрические признаки: волосатость ушей, перепонки между пальцами ног, ихтиоз (кожа имеет глубокую исчерченность и напоминает рыбью чешую). Гомологичные районы Х- и Y-хромосом содержат аллельные гены. Они с равной вероятностью встречаются у лиц мужского и женского пола. К числу таких признаков относятся общая цветовая слепота (отсутствие цветового зрения) и пигментная ксеродерма — заболевание, при котором под влиянием ультрафиолетовых лучей на открытых частях тела образуются опухоли. Оба признака являются рецессивными. Признаки, которые определяются аллель-ными генами, находящимися в Х- и Y-хромосомах, наследуются как классические менделирующие признаки. Наследование ограниченное и контролируемое полом. Признаки человека, наследование которых каким-то образом связано с полом, подразделяется на несколько категорий. Одна из категорий — признаки, ограниченные полом. Развитие этих признаков обусловлено генами, расположенными в аутосомах обоих полов, но проявляющимися только у одного пола. Например, гены, определяющие ширину таза женщины, локализованы в аутосомах, наследуются и от отца, и от матери, но проявляются только у женщин. То же касается возраста полового созревания женских индивидов. Среди мужских признаков, ограниченных полом, можно назвать количество и распределение волосяного покрова на теле. К иной категории относятся признаки, контролируемые полом или зависимые от пола. Развитие этих соматических признаков обусловлено генами, расположенными в аутосомах. Проявляются ониу мужчин и женщин, но по-разному. Например, у мужчин раннее облысение — признак доминантный, он проявляется у доминантных гомозигот (АА) и у гетерозигот (Аа). У женщин этот признак рецессивный и проявляется только у рецессивных гомозигот (аа). Для признаков, контролируемых полом, выраженность обусловлена в большой степени половыми гормонами. |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 481; Нарушение авторского права страницы