|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Тема 10. ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА
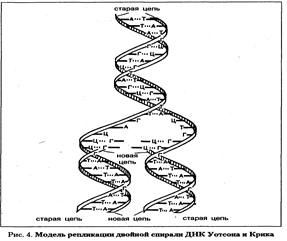
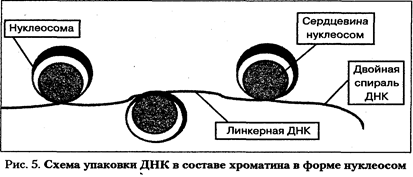
Хромосомы. Носителем наследственной информации в живых организмах являются молекулы ДНК. В 1953 г. Дж.Уотсон и Ф.Крик предложили модель структуры ДНК, которая с тех пор многократно проверялась и признана правильной. Нативная молекула ДНК представлена двумя длинными полимерными цепями, соединенными между собой и закрученными в форме двойной спирали. Основная структурная единица одной цепи — нуклеотид. Он состоит из соединенных ковалентньши связями дезоксирибозы, азотистого основания и фосфатной группы. ДНК содержит пури-новые азотистые основания (аденин и гуанин) и пиримидиновые основания (цитозин и тимин). Азотистое основание ковалентио соединено с первым атомом углерода дезоксирибозы и формирует структуру, называемую нуклеозидом. Фосфатные группы соединяют соседние нуклеозиды в полимерную цепочку. Сцепление между цепями обеспечивается особыми водородными связями между аденином (А) и тимином (Т) и между гуанином (Г) и цитозином (Ц). Водородные связи много слабее ковалентных, соединяющих отдельные атомы каждого нуклеотида, но достаточно сильны, чтобы обеспечить специфичность образования пар А—Т, Г—Ц. Такие специфичные пары азотистых оснований называются комплементарными. Комплементарность азотистых оснований — один из основных принципов организации молекулы ДНК. Две полимерные депи молекулы ДНК соединены между собой антипараллельно. Нуклеиновые кислоты — очень длинные полимерные цепочки. Интактные молекулы ДНК содержат в зависимости от вида организмов от нескольких тысяч до многих миллионов нуклеотидов. Для любой последовательности азотистых оснований возможна равная ей по длине комплементарная последовательность, составляющая вторую цепь двойной спирали. Конкретная последовательность пар А—Т и Г—Ц не влияет на структуру молекулы ДНК, образующей двойную спираль. Возможное число различных последовательностей пар оснований в молекуле ДНК практически бесконечно и способно кодировать колоссальное количество информации. Поскольку цепи ДНК комплементарны, каждая из них при рас-плетании двойной спирали способна служить матрицей для синтеза новой комплементарной цепи. Последовательность оснований во вновь синтезируемой цепи будет определяться спецификой водородных связей между азотистыми основаниями родительской и вновь синтезируемой цепи. Таким образом, генетическая информация, содержавшаяся в последовательности пар оснований родительской молекулы, будет полностью воспроизведена в двух дочерних молекулах. Более того, если в процессе удвоения ДНК произошла ошибка и какой-либо нуклеотид во вновь образуемой цепи выпал или оказался некомплементарным исходному, то это может изменить информационное содержание молекулы. Причем логично ожидать, что эта ошибка будет передана дочерним молекулам ДНК в следующих поколениях. Такая замена пары нуклеотидов будет обладать свойствами генетической мутации. Модель структуры ДНК Уотсона и Крика объясняет как способность генов к самоудвоению (репликации), так и их информационные свойства. Молекулы ДНК в эукариотических клетках упакованы с помощью белков и находятся в составе хроматина в ядре. Белки, входящие в состав хроматина, разнообразны. Обычно их разделяют на две группы: гистоны и негистоновые белки. Гистоны, характерные только для эукариотических клеток, осуществляют первые этапы упаковки ДНК, очень схожие у большинства изученных объектов. Их взаимодействие с ДНК происходит за счет ионных связей и не зависит от последовательности нуклеотидов в составе молекулы ДНК. Гистоны не отличаются большим разнообразием. Это глобулярные белки, представленные 5-ю - 7-ю типами молекул. Наиболее известны следующие классы гистонов: HI, H2A, Н2В, НЗ и Н4. Их основные свойства определяются относительно высоким содержанием аминокислот лизина и аргинина. Положительные заряды на аминогруппах указанных аминокислот обеспечивают электростатическую связь гистонов с отрицательными зарядами на фосфатных группах ДНК. Первый этап упаковки ДНК в хроматине осуществляется с помощью нуклеосом. Молекула ДНК накручивается на поверхность множества белковых частиц, каждый раз делая 1,75 оборота вокруг сердцевины. Сердцевина нуклеосомы всегда консервативна и содержит восемь молекул - по две молекулы гистонов Н4, НЗ, H2A, Н2В. По поверхности сердцевины располагается участок ДНК из 146 нуклеотидных пар. Небольшой участок ДНК между нуклеосо-мами остается несвязанным с сердцевиной, он называется линкером. Общий вид хроматина, представленного молекулой ДНК, упакованной с помощью нуклеосомных структур, можно сравнить с бусами на нитке. Первый нуклеосомный уровень компактизации укорачивает молекулу ДНК в 6-7 раз.
В следующий этап упаковки нуклеосомная структура хроматина вовлекается с помощью гистона HI, который связывается с линкерной частью ДНК и поверхностью нуклеосомы. Благодаря сложному взаимодействию всех компонентов, возникает упорядоченная структура спирального типа, которую часто называют соленоидом. Она повышает компактность ДНК еще в 40 раз. Такая спирализация ДНК может играть роль фактора, инактивирующего гены. Под электронным микроскопом соленоид выявляется в виде фибрилл хроматина. Более высокие уровни компактизации ДНК в хроматине связаны с негистоновыми белками. Некоторые из них специфично связываются с определенными участками ДНК, в результате чего фибриллы хроматина в местах связывания ДНК с негистоновыми белками образуют петли. Таким образом, более высокие уровни упаковки ДНК в составе хроматина обеспечиваются не спирализацией нитей хроматина, а образованием поперечной петлистой структуры вдоль хромосомы. На всех указанных этапах компактизации ДНК хроматин представлен в активной форме, в нем происходит транскрипция - синтез всех типов молекул РНК. Такой хроматин называют эухроматином. Дальнейшая упаковка хроматина ведет к переходу его в неактивное состояние с образованием гетерохроматина. Этот процесс связан со спирализацией групп петель и образованием из фибрилл хроматина розеткоподобных структур, которые называются хромомерами. Дальнейшая суперспирализация хроматина приведет к образованию хромосом. В настоящее время считается, что в состав каждой хромосомы входит одна двуспиральная молекула ДНК, которая может удваиваться при подготовке клеток к делению, после чего каждая из двух одинаковых молекул упаковывается независимо, образуя хроматиды. Хроматиды соединяются между собой в районе первичной перетяжки, или центромеры. Центромеры выполняют в хромосомах очень важные функции. Они соединяют две сестринские хроматиды, также велика их роль в организации веретена деления. В районе центромеры к хроматидам прикрепляются микротрубочки веретена деления. Центромера располагается внутри гетерохроматического района, где находится сателлитная ДНК, представленная кластерами высокоповторяющихся последовательностей. Одновременно с этим внутри центромеры выявляются уникальные последовательности, которые, вероятно, несут информацию о расхождении хромосом к противоположным полюсам клетки. Концы хромосом называются теломерами. Они также выполняют важные функции. Когда деление клетки закончено и формируются новые клеточные ядра, с помощью теломер хромосомы прикрепляются к внутренней ядерной мембране, в результате чего каждая хромосома в деспирализованном состоянии занимает в ядре строго определенное место. Помимо этого, теломерные районы предотвращают слипание хромосом своими концами и препятствуют образованию дицентриков — хромосом с двумя центромерами, наличие которых свидетельствует о патологических картинах митоза. Одновременно теломеры стабилизируют хромосомы, защищая их от деградации клеточными нуклеазами. В последнее время стало известно еще одно назначение теломерных концов. Благодаря им, происходит полное завершение редупликации хромосом при подготовке клетки к делению. Хромосомы, состоящие из двух хроматид, выявляются во время деления клеток. В Них генетический материал неактивен. В период между клеточными делениями хромосомы частично раскручиваются, деспирализуются, переходят в состояние хроматина, и гены, вошедшие в состав эухроматина, активно работают. Участки ДНК, упакованные в виде гетерохроматина, могут иметь двоякую природу. Различают гетерохроматин факультативный и конститутивный (структурный). Факультативный гетерохроматин представляет собой участки генома, временно инактивированные в тех или иных клетках. Примером такого хроматина служит половой гетерохроматин инактивированной Х-хромосомы в соматических клетках женщин. Структурный гетерохроматин во всех клетках постоянно находится в неактивном состоянии и, вероятно, выполняет структурные или регуляторные функции. Таким образом, упаковка ДНК в составе хроматина и хромосом имеет двоякое значение. С одной стороны, происходит упорядоченное укорачивание длинных молекул, с другой — изменяется активность генов в зависимости от плотности компактизации ДНК, т.е. чем более суперскручена молекула, тем менее активны гены, находящиеся в этой структуре. В частности, активность митотических хромосом практически равна нулю. |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 201; Нарушение авторского права страницы