|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Катаболизм белков в мышечных волокнах
Белки, входящие в состав клеток организма, также постоянно обновляют свой состав, разрушаются (; катаболизм или протеолиз) и снова синтезируются. В мышечных волокнах распад белков осуществляется посредством специальных ферментов — катепсинов, которые находятся в лизосомах. Многие из этих ферментов проявляют высокую активность в кислой среде (рН = 5, 0). Поэтому при мышечной работе, приводящей к образованию в мышцах молочной кислоты, катепсины активизируются. Мембрана лизосом устроена таким образом, что она проницаема для белков, которые могут свободно проникать внутрь лизосом, но не проницаема для катепсинов. В лизосомах белки под действием катепсинов превращаются в олигопротеины (низкомолекулярные белки) и выходят из лизосом в саркоплазму мышечного волокна. В саркоплазме олигопротеины под действием ферментов расщепляются до аминокислот. Срок полупериода жизни белков мышц, то есть время, за которое они наполовину обновляют свой состав, составляет около 30 суток. Физические нагрузки усиливают синтез и распад белков в мышцах. Степень выраженности этих процессов зависит от интенсивности нагрузки и тренированности организма человека. В качестве специфического показателя распада сократительных белков актина и миозина используется 3-метилгистидин. Однократные физические нагрузки вызывают угнетение синтеза белка и усиление их катаболизма. Интенсивная силовая тренировка особенно усиливает катаболизм белков и некоторых структур мышц, на восстановление которых требуется продолжительное время (до двух- трех суток). Особенно увеличивается синтез тех белков, которые разрушаются в большей степени. Процессы распада белков и окисления аминокислот сопровождаются усиленным образованием при мышечной деятельности аммиака, который связывается в печени в цикле синтеза мочевины и выводится из организма. Поэтому физические нагрузки вызывают увеличение содержания мочевины в крови, а нормализация ее в период отдыха свидетельствует о восстановлении процессов распада и синтеза белков в тканях. Установлено, что адаптационные процессы, происходящие в скелетной мышце и приводящие к ее гипертрофии, связаны не только с повышенным синтезом белка, но и с пониженным его распадом. Этот феномен называется антикатаболическим эффектом. Он достигается за счет разрушения белка, индуцированного кортизолом. Считается, что гипертрофия мышечных волокон типа I зависит от уменьшения разрушения белков, а гипертрофия мышечных волокон типа II — от увеличения синтеза белка. Синтез белков в мышечных волокнах Синтез белка в мышечных волокнах протекает в несколько этапов. В настоящее время принято различать три этапа синтеза белка: транскрипцию, рекогницию и трансляцию.
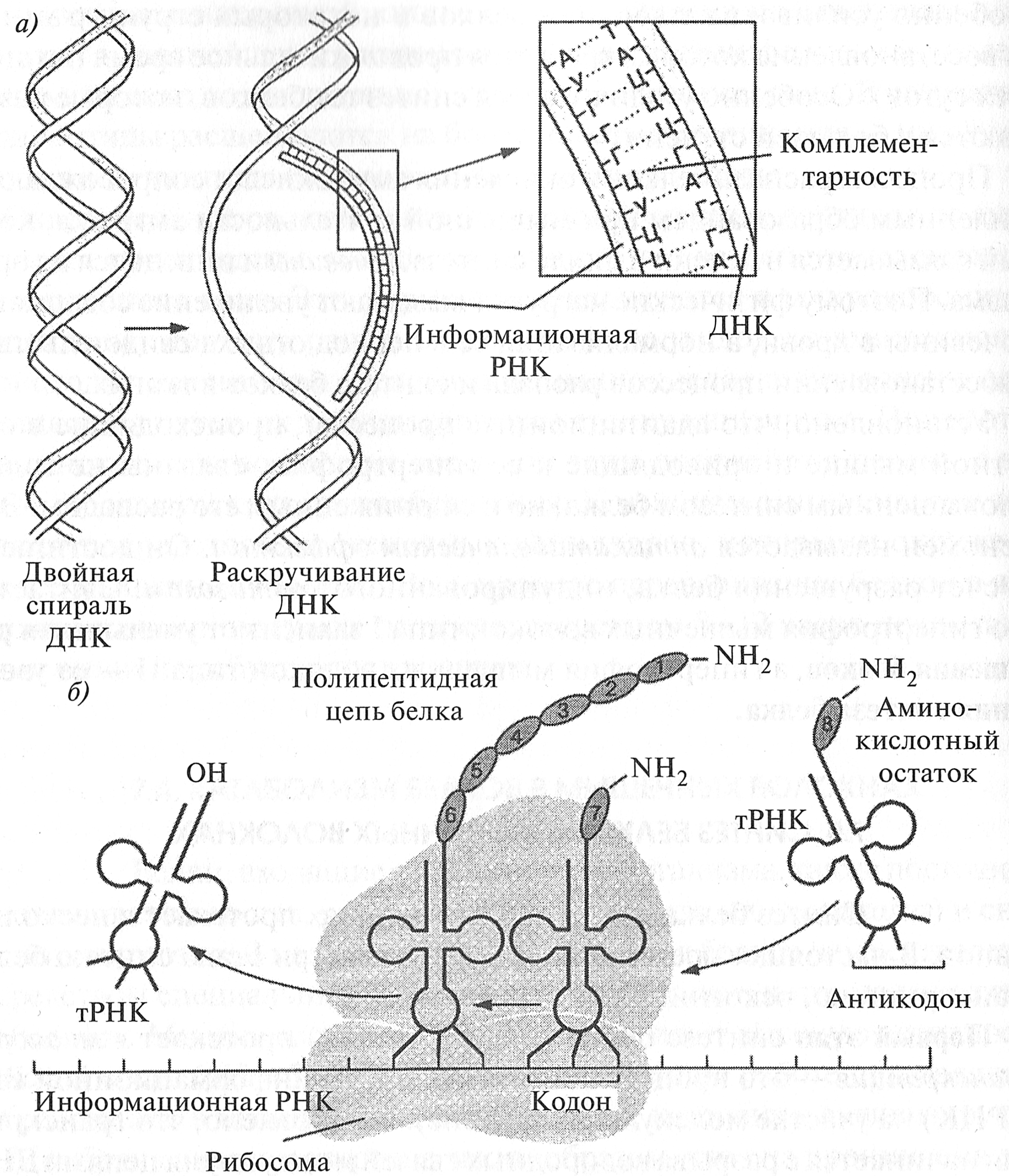
Рис. 7.2. Этапы синтеза белка: а - транскрипция; б - трансляция Первый этап синтеза белка (транскрипция) протекает в миоядрах. Транскрипция — это процесс синтеза молекулы информационной РНК (иРНК) на участке молекулы ДНК (гене). Установлено, что транскрипция начинается с разрыва водородных связей между двумя цепями ДНК. Затем происходит «раскручивание» участка спирали ДНК. Специальный фермент — РНК-полимераза, — двигаясь по цепи ДНК, подбирает по принципу комплементарности нуклеотиды и соединяет их в цепочку. В результате на одной из цепей ДНК синтезируется молекула иРНК (рис. 7.2). Таким образом происходит переписывание (транскрипция) информации о структуре синтезируемого белка с участка ДНК. Затем иРНК выходит из ядра в саркоплазму мышечного волокна и перемещается в область рибосом. ДНК восстанавливает свою структуру. Доказано, что силовая тренировка приводит к увеличению количества миоядер (F. Kadi et al., 1999). Это увеличение связано с усиленным делением клеток-сателлитов при повреждении мышечного волокна. Возрастание количества миоядер создает предпосылки для увеличения синтеза белка, так как увеличивается количество иРНК, синтезируемой в ядрах мышечных волокон. Второй этап синтеза белка — рекогниция — проходит в саркоплазме мышечного волокна. В процессе рекогниции транспортные РНК (тРНК) соединяются с аминокислотами и транспортируют их в область рибосом. Так как существуют 20 основных аминокислот, то насчитывается более 20 видов тРНК.
Второй этап синтеза белка — рекогницию — называют еще этапом активации аминокислот. Установлено, что для каждой аминокислоты имеются свои специфические ферменты, которые участвуют в ее активации. Эти ферменты проявляют высокую активность в присутствии ионов магния (Mg2+). Молекулы тРНК имеют по два специфических триплета. Один из них — кодон, с которым связывается аминокислота, другой — антико- дон, который соответствует кодону данной аминокислоты в иРНК (см. рис. 7.2). Благодаря этому аминокислоты при синтезе белка располагаются в последовательности, диктуемой последовательностью кодонов виРНК. Третий этап синтеза белка — трансляция — происходит на рибосомах. Из аминокислот, перемещаемых тРНК в область рибосом, формируется пептидная цепь (белковая молекула) в соответствии с информацией, закодированной в иРНК. Молекула иРНК передвигается между двумя субъединицами рибосомы — малой и большой. К малой субъединице присоединяется иРНК, к большой — фермент, синтезирующий белок. При передвижении иРНК между двумя субъединицами рибосом кодо- ны иРНК взаимодействуют с антикодонами тРНК по принципу комплементарное™. Завершение синтеза белка обеспечивается кодонами терминации (стоп-сигналом) в иРНК, с которыми не может связаться ни одна тРНК. Поэтому процесс завершения синтеза белка называется тер- минацией. Затем включается фактор освобождения и полипептидная цепь белка отделяется от рибосом. Таким образом образуется первичная структура белка. Затем вновь синтезированный белок приобретает определенную пространственную вторичную структуру. Сложная третичная структура белка формируется самопроизвольно в саркоплазме и определяется характером первичной структуры белка, а также окружающими условиями. Аналогичным образом осуществляется синтез молекул основных сократительных белков (миозина и актина). Следует отметить, что синтез белка требует огромных затрат энергии. Так, только для присоединения одной аминокислоты к полипептидной цепи синтезируемого белка используется по меньшей мере пять молекул АТФ, поэтому процесс синтеза белка во многом зависит от скорости восстановления уровня АТФ в мышечных волокнах. Миофибриллогенез Под термином миофибриллогенез понимается формирование в мышечном волокне специальных органелл — миофибрилл. Итак, после третьего этапа синтеза сократительных белков в мышечном волокне синтезируются молекулы различных белков: миозина, акти- * Триплет — комбинация из трех последовательно расположенных нуклеотидов в молекуле нуклеиновой кислоты. на, тропомиозина, тропонина и многих других. Каким же образом из этих молекул формируется структурная единица саркомера, состоящая из одного толстого и шести тонких филаментов? Каким образом увеличивается число миофибрилл, а также площадь их поперечного сечения? Установлено, что толстые и тонкие филаменты формируются путем самосборки. В растворах слабой ионной силы молекулы миозина начинают агрегировать (собираться) в более сложные структуры — толстые филаменты. При этом миозиновые молекулы объединяются «хвост в хвост» и образуются биполярные нити. Этот процесс продолжается за счет добавления молекул миозина на каждом конце нити до тех пор, пока их длина не достигнет 1, 5 мкм. К этому времени они становятся похожими на толстые филаменты. Следует заметить, что процесс образования такой сложной структуры, как толстый филамент, очень чувствителен к показателю рН. Вне области физиологических значений рН (7, 2—7, 5) агрегация (самосборка) часто происходит нерегулярно, так что возникают разветвляющиеся структуры и очень длинные агрегаты (Дж. Бендолл, 1970). Из этого следует, что смещение рН в мышцах в кислую сторону, что имеет место при накоплении молочной кислоты в мышцах, отодвигает сборку миофибрилл до того момента, пока рН мышц не достигнет физиологических значений. Тонкие филаменты также формируются путем самосборки. После того как на рибосомах синтезируется полипептидная цепь белка актина, она автоматически сворачивается в G-форму, которая представляет собой третичную структуру белка. При этом молекулы глобулярного актина (G-актина) в присутствии АТФ, ионов магния (Mg2+) и калия (К+) полимеризуются в длинные нити фибриллярного актина (F-актина), составляющего основу тонкого филамента. После этого нити фибриллярного актина скручиваются в двойную спираль. Если в растворе находятся толстые и тонкие филаменты, то они вступают друг с другом в связь и возникает так называемый актомио- зиновый комплекс {сот). В результате взаимодействия между толстыми и тонкими филаментами возникают структуры с периодическим чередованием толстых и тонких филаментов, сходные с натуральными по- перечно-полосатыми миофибриллами. При этом значительно повышается вязкость раствора. В 40-х годах XX века Альберт Сент-Дьерди показал, что возрастание вязкости уменьшается, если в раствор добавить молекулы АТФ. Было доказано, что добавление АТФ в раствор вызывает диссоциацию акто- миозинового комплекса на актин и миозин. Если же актомиозиновый комплекс помещался в раствор, содержащий ионы калия, магния и молекулы АТФ, актомиозиновые нити сокращались. В тех же условиях нити, образованные только из миозина, не сокращались. Эти опыты позволили установить, что мышечное сокращение возникает в результате взаимодействия миозина, актина и АТФ (Л. Страйер, 1985). На последней стадии морфогенеза миофибриллярного аппарата приводится в соответствие поперечная исчерченность соседних миофибрилл (Е.К. Жуков, 1974), то есть формируется цитоскелет мышечного волокна. Ответственными за поперечную исчерченность являются цитоскелетные белки, к одним из которых относится десмин. Этот белок связывает Z-диски соседних миофибрилл. Показано, что в процессе регенерации миофибрилл после эксцентрических упражнений увеличивается синтез десмина и происходит перестройка цитоскелета мышечного волокна (J. Friden, U. Kjorell, L.E. Thornell, 1984; J. Frieden, R.L. Lieber, 1992). Таким образом, общая картина морфогенеза миофибриллярного аппарат следующая. На первой стадии происходит сборка исходных белковых составных частей в неисчерченные нитевидные структуры миофиламентов. На второй стадии осуществляется распределение образовавшихся компонентов в поперечно-полосатые структуры — миофибриллы. На третьей стадии происходит приведение в соответствие расположение саркомеров в соседних миофибриллах (Е. К. Жуков, 1974). При рассмотрении параметров, влияющих на поперечное сечение мышечного волокна, было показано, что его увеличение может осуществляться за счет миофибриллярной гипертрофии — возрастания количества миофибрилл и их объема. Теория повреждения миофибрилл позволяет объяснить этот феномен. В детском возрасте миофибриллы «растут» с увеличением длины конечностей (J. D. MacDougall, 2003). Так как при росте конечностей мышца находится в постоянном растяжении, миофибриллы постепенно увеличивают свою длину. Это происходит за счет того, что на концах миофибрилл осуществляется их «достройка» посредством сборки дополнительных саркомеров. Таким способом увеличивается длина миофибрилл. Возможен также полный поперечный разрыв миофибриллы. При этом каждая «культя» увеличивает свою длину посредством достройки (сборки дополнительных саркомеров). В результате вместо одной миофибриллы получаются две.
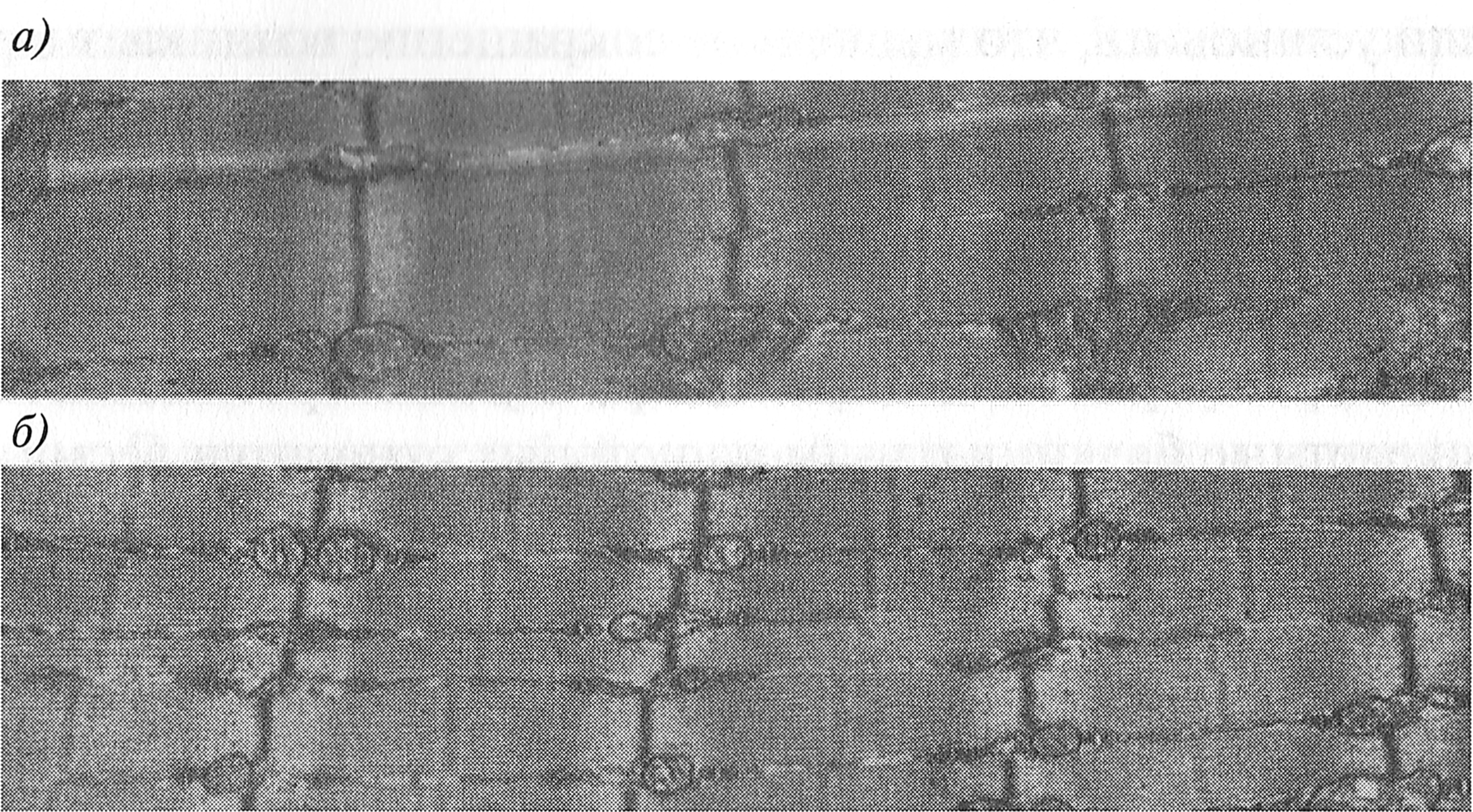
Рис. 7.3. Продольное расщепление миофибриллы двуглавой мышцы плеча мыши: а — начало расщепления миофибриллы в области двух Z-дисков; б — расщепленные миофибриллы (G. Goldspink, 1971) Доказано также продольное расщепление миофибрилл. Это происходит тогда, когда объем миофибриллы (или ее поперечное сечение) становится очень большим и достигает критических значений. Чаще всего расщепление миофибриллы начинается в области Z-дисков (рис. 7.3). При этом в центре диска появляется небольшое отверстие, которое затем расширяется. В результате образуются две «дочерние» миофибриллы (G. Goldspink, 1970, 1971). Причину продольного расщепления миофибриллы G. Goldspink (1971) видит в том, что тонкие филаменты прикрепляются к Z-диску под небольшим углом. При развитии напряжения мышцы этот угол увеличивается, сила тяги тонких филаментов возрастает и повреждает Z-диск. Если напряжение мышцы происходит очень быстро, что имеет место в быстрых мышечных волокнах, Z-диски повреждаются еще быстрее и чаще происходит расщепление миофибрилл. Поэтому, как считает G. Goldspink (1971), миофибриллы в быстрых мышечных волокнах, как правило, небольшие, в то время как в медленных — более крупные. Новые миофибриллы могут возникать в результате деления и объединения клеток сателлитов. Таким способом может увеличиваться число миофибрилл в мышечном волокне. Поперечное сечение миофибрилл возрастает за счет добавления синтезированных толстых и тонких филаментов на их поверхности (Е. К. Жуков, 1974; J. D. MacDougall, 2003). Однако плотность упаковки миофиламентов не возрастает, расстояние между толстым и тонкими филаментами не меняется. Так как увеличение количества миофибрилл и их объема влияет на объем мышечного волокна, возникает гипертрофия мышечных волокон по миофибриллярному типу. |
Последнее изменение этой страницы: 2019-04-09; Просмотров: 459; Нарушение авторского права страницы