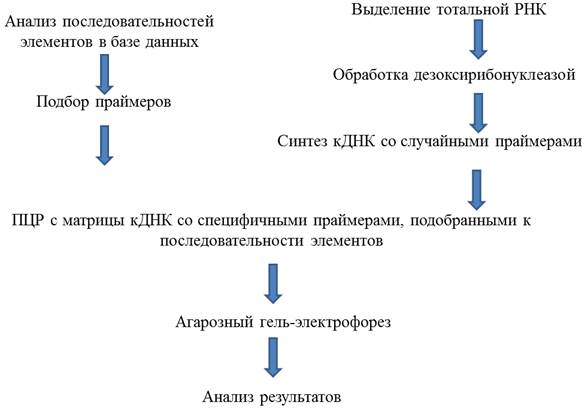
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Общая структура экологической генетики (Инге-Вечтомов и др.,Стр 1 из 9Следующая ⇒
Экологическая генетика. Экологическая генетика как наука образовалась во второй половине XX века. В 60-е годы Е.Б.Форд впервые сформулировал понятие «экологическая генетика» как генетику популяций животных в природных условиях (Ford, 1964). Экологическая генетика - междисциплинарная наука, вобравшая в себя закономерности и постулаты двух базовых дисциплин – экологии и генетики. Э. Геккель во второй половине XIX века определил экологию как науку об отношениях организмов с окружающей средой и с другими организмами (Гекель, 1866). Это определение охватывает не только область экологии, но и область смежных наук, в том числе и экологической генетики. Генетика, как определил ее У. Бэтсон в 1906 г.,- это наука о закономерностях наследственности и изменчивости. Таким образом, экологическая генетика - это область знания, использующая взаимовлияния генетических процессов и экологических отношений (Инге - Вечтомов, 2009). Область знаний, охватываемая экологической генетикой, представлена в табл.1.1. Табл. 1.1 Общая структура экологической генетики (Инге-Вечтомов и др., 1999).
Экологическая генетика опирается на мощную методологию генетического анализа и на методический арсенал экологии. Одним из вопросов экологической генетики является изучение генетического контроля устойчивости особей. Каждый организм в течение своей жизни подвергается воздействию неблагоприятных экологических факторов, таких как резкие перепады температуры, недостаток питания, гипероксия или гипоксия, поллютанты, ультрафиолетовое и ионизирующее излучение и др. Для перенесения неблагоприятных условий у живых организмов сформировались различные приспособления как на уровне индивида в течение онтогенеза (физиологическая адаптация), так и в ходе эволюции на уровне вида (генетическая адаптация). В последние годы, в связи с ухудшением состояния окружающей среды и возросшей на этом фоне заболеваемости человека, активно исследуются заболевания, развитию которых способствуют, с одной стороны, неблагоприятные факторы среды, а с другой стороны - неодинаковая реакция разных особей на данные факторы, обусловленная их генотипом. Было показано, что многие заболевания, например астма (Polonicov et al., 2009), колоректальный рак (Mala et al., 2009), папиллярный рак щитовидной железы (Siraj et al., 2008) и другие ассоциированы с некоторыми полиморфными вариантами генов ферментов трансформации ксенобиотиков. Биотрансформация ксенобиотиков – многостадийный процесс, и, соответственно, контролируется полигенно, в связи с чем, большое значение имеет исследование взаимодействия разных аллельных вариантов генов, участвующих в данном процессе. Важно отметить, что некоторые полиморфные варианты рассматриваемых генов могут иметь приспособительный характер. Например, частота аллеля 432L гена CYP1B1, выше у здоровых людей, по сравнению с людьми, имеющими бронхиальную астму. А отсутствие аллеля 462IV гена CYP1A1 приводит к увеличению вероятности возникновения астмы у курящих людей, тогда как у некурящих такого эффекта не наблюдается. Важно отметить, что один и тот же полиморфизм гена в разном генетическом окружении может по-разному влиять, оказывая как положительный, так и отрицательный эффект. Это объясняется взаимодействием генов. Например, взаимодействие аллеля Y113H гена EPHX1и аллеля V432L гена CYP1B1 приводит к увеличению вероятности развития заболевания (Polonicov et al., 2009). Другой яркий пример наследственной устойчивости к факторам окружающей среды – невосприимчивость отдельных индивидуумов к вирусу иммунодефицита человека (ВИЧ-1). Это обусловлено аллелем CCR5-Δ32 гена CC-CKR-5, в результате работы которого образуется нефункциональный белок – рецептор, вследствие чего, вирус иммунодефицита человека теряет способность связываться с рецептором и проникать в клетку (Liu et al., 1997; Djin-Ye Oh et al., 2008). Таким образом, генотип влияет на приспособленность организмов к окружающей среде. В борьбе за существование выживают более приспособленные особи. Менее приспособленные особи гибнут, либо не размножаются, либо производят на свет меньше полноценных потомков (Северцов, 2005). Характеристикой приспособленности может являться продолжительность жизни особи. Логично предположить, что чем выше продолжительность жизни, тем больше потомства может оставить особь. Продолжительность жизни является интегральным показателем стрессоустойчивости организма, так как она зависит от генотипа особи и от влияния экологических стрессоров (Петин, Сынзыныс, 1998).Поэтому, более узкой проблемой экологической генетики можно считать изучение влияния генотипа на продолжительность жизни модельных объектов. Продолжительность жизни. Продолжительность жизни - это мультифакторный признак, один из наиболее значимых количественных признаков организма. Старение с точки зрения биологии - это процесс постепенного угнетения основных функций организма, в том числе регенерационных и репродуктивных, вследствие чего организм становится менее приспособленным к условиям окружающей среды (теряется способность противостоять стрессам, болезням, травмам), что делает гибель организма неизбежной. Биология продолжительности жизни - это раздел биологии, занимающийся изучениемзакономерностей длительности жизни организмов, а также механизмов ее определяющих (Гаврилов и др.,1991). Среди наиболее актуальных ее проблем, выделяют: проблему наследуемости и изменчивости длительности жизни (Lints et al, 1989), проблему половых различий по срокам жизни (Hazzard, 1986), а также проблему изменения продолжительности жизни в процессе биологической эволюции (Wodhead, 1987). Таким образом, в задачи биологии продолжительности жизни входит выяснение причин индивидуальных различий по срокам жизни, а также причин межпопуляционных и межвидовых различий по этому признаку. II .МАТЕРИАЛЫ И МЕТОДЫ Материалы. В эксперименте использовались плодовые мухи Drosophila melanogaster (Meigen, 1830). Линии мух, используемые в эксперименте, и их характеристики представлены в таблице 2.1. Табл. 2.1 Линии мух и их характеристики.
Методы Синтез кДНК. Синтез кДНК делали по протоколу набора фирмы “Fermentas” “First Strand cDNA Synthesis Kit”. 1. РНК обрабатывали DNase I (“Fermentas”) в двойном объеме для удаления ДНК в растворе. Состав смеси показан в таблице 2.2 Табл. 2.2 Состав смеси для обработки DNase.
Табл. 2.3. Состав 10X буфера для DNaseI («Fermentas»)
2. Инкубировали смеси при температуре 37°С 1 час. 3. Добавляли к смеси EDTA до конечной концентрации в растворе 2,27 мМ для ингибирования DNase I, перемешивали, инкубировали при температуре 65°С 10 минут. 4. К раствору РНК добавляли Random Primer, перемешивали и инкубировали при 65°С 5 минут, затем охладили на льду (см. табл.2.4). 5. Добавляли dNTP, ингибитор RNase Ribolock, реакционный буфер, обратную транскриптазу (см. табл. 2.4). Табл. 2.4 Реактивы для обратной транскрипции.
Табл.2.5 Состав 5X Reaction buffer
6. Инкубировали при 25°С 5 минут, затем при 37°С 1 час, затем при 70°С 10 минут. Выделение ДНК из мух. 1. Мух в количестве 50 штук помещали в пробирку объемом 1,5 мл. 2. Гомогенизировали биологический материал с помощью тефлонового пестика до получения однородной массы в присутствии рабочего раствора (см. табл. 2.11). Табл. 2.11 Состав рабочего раствора.
Табл. 2.12 Состав 1X Стока А.
3. Инкубировали при температуре 70˚С 30 минут, добавили 5 М ацетат калия ( на 500 μl рабочего раствора 116 μl ацетата). 4. Инкубировали на льду 30 минут, центрифугировали 5 минут при 10g. 5. Добавили равный объем смеси фенола с хлороформом (1:1), центрифугировали 10 мин при 10 g. 6. Отобрали верхнюю фазу, добавили изопропанол (350 μl). 7. Инкубировали 7 минут при комнатной температуре, центрифугировали 15 минут при 10 g, супернатант сливали. 8. Добавляли 70% этанол (0,5 мл), центрифугировали 15 минут при 10 g, супернатант сливали. 9. Осадок просушивали, растворяли его в небольшом количестве воды Nuclease Free Water в зависимости от величины осадка (10-40 мкл). 10. Концентрацию ДНК измеряли с помощью спектрофотометра (фирма «Nanodrop»). Рестрикция. Рестрикцию проводили эндонуклеазами рестрикции фирмы “Fermentas” в специфических буферах, условия работы рестриктаз показаны в таблицах 2.13-2.19, состав рестрикционной смеси показан в таблице 2.20. Табл. 2.13 Условия работы и инактивации эндонуклеаз рестрикции.
Табл.2.20 Состав рестрикционной смеси.
Лигирование. Состав лигазной смеси показан в таблице 2.21. Табл. 2.21 Состав лигазной смеси.
Синтез зонда. Зонд синтезировали ПЦР - методом с геномной ДНК с специфическими праймерами. Состав ПЦР смеси указан в таблице, но в данной реакции ДНК брали около 1000 ng на реакцию. Параметры ПЦР: 1. 2.1. Денатурация ДНК: Т=95оС, 30 сек 2.2. Отжиг праймеров: Т (см. табл.), 1 мин 30 циклов 2.3. Синтез: Т=72оС, 1 мин 3. Синтез: Т=72оС, 3 мин Праймеры для ПЦР подбирались с помощью программы VectorNTI 9 (табл. 2.25). Табл. 2.25 Праймеры для синтеза зонда.
Очистка зонда. ПЦР продукт анализировали в агарозном гель - электрофорезе № 1 (см. табл.), вырезали подходящие по размеру полосы геля под UV- лампой. Собирали в пробирки объемом 1,5 мл. Выделяли ДНК набором «Fermentas». 1. Добавили 3 :1массе геля Binding Buffer. 2. Грели 5 минут при 55˚ С, каждую минуту инвентировали. 3. Добавить ресуспендированный Silica Powder Suspension. ≤2, 5 mg ДНК- 5 мкл ≥2,6 mg ДНК- 2 мкл на 1 mg ДНК 4. Инкубировать 5 минут при 55 ˚ С. Мешали каждую минуту, чтобы сохранить Silica Powder Suspension в состоянии суспензии. 5. Центрифугировали 5 секунд при 10 g, супернатант сливали. 6. Повторяли пункт 1-5. 7. Добавляли Wash Buffer. Табл.2.26 Состав Wash Buffer.
8. 1) Ресуспендировать осадок. 2) Центрифугировали 5 секунд при 10 g. 3) Удаляли супернатант. Повторяли подпункт 1-3 три раза 9. Сушили осадок 10минут 10. Растворяли его в небольшом количестве воды Nuclease Free Water в зависимости от величины осадка (10-40 мкл). 11. Концентрацию ДНК измеряли с помощью спектрофотометра (модель «Nanodrop» фирма Thermo Fisher Scientific). Мечение зонда. Метили зонд, гибридизовали мембрану с зондом, отмывали блот от излишков зонда с помощью набора «Amersham Gene Images AlkPhos Direct Labelling and Detection System». 1. ДНК доводили до концентрации 10 нг/ мкл, концентрация соли в ней должна быть не более 50 мМ, брали 10 мкл. 2. Денатурировали ДНК - кипятить в течение 5 мин, затем резко переносили в лед. 3. Добавляли Reaction buffer (10 мкл), аккуратно перемешать. Реакция проходит на льду. 4. Добавляли реагент маркировки Labeling reagent (10 мкл) , перемешивали аккуратно на льду. 5. Добавляли соединительного агента рабочего Cross-linker (2 мкл), центрифугировали при 10 g. Рабочий Cross-linker :20 мкл стокового Cross-linker и 8 мкл воды Nuclease Free Water. 6. Инкубировали 30 минут при 37 ˚ С. Зонд использовали либо сразу, либо в течение 2-х часов, при условии хранения его на льду. Отмывка блота. 1. Инкубировали Wash buffer 1 (см. табл.2.29) в чашке Петри на протяжении 10 минут при 55 ˚С. Табл. 2.29 Состав Wash buffer 1.
2. Переносили блот в Wash buffer 1 и промывали на протяжении 10 минут при 55 ˚С. Повторяли процедуру в новом согретом Wash buffer 1. 3. Переносили блот в Wash buffer 2 (см. табл.2.30) и инкубировали при комнатной температуре 5 минут. Повторяли процедуру в новом Wash buffer2. Табл. 2.30 Состав Wash buffer 2.
Табл. 2.31 Состав 20Х Secondary wash buffer.
4. Использовали в течение 30 минут. Постановка эксперимента. Отбирали 250 виргинных самок и самцов 3-х линий. Помещали отдельно самцов, отдельно самок в пробирки (высота - 20см., диаметр – 2см.) по 5 штук в каждую. Пересадка на свежий корм осуществлялась раз в неделю. Сбор данных. Подсчет умерших мух осуществляли ежедневно, кроме воскресенья. Общая схема эксперимента. Общая схема эксперимента представлена на рисунке 3.1.
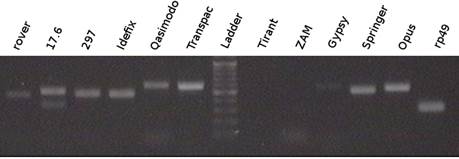
Рис. 3.1. Общая схема эксперимента. 3.2. Анализ экспрессии МГЭ группы Gypsy в линиях D . melanogaser . Как было отмечено выше, транспозиционная активность gypsy контролируется геном flamenco по механизму РНК - интерференции. Не исключено, что этот ген контролирует транспозицию и других элементов этой группы. В поставленном нами эксперименте участвовали три изогенные линии, отличающиеся по гену flamenco (см. табл. 2.1.): MS (145), SS (7K), SS (C3( 19)), причем, в линии MS (145) присутствует активный ретровирус gypsy. Первым этапом в цепи событий, происходящих при транспозиции ретроэлементов, является их транскрипция, в результате чего образуется РНК, часть которой используется как матрица для обратной транскриптазы, другая часть подвергается сплайсингу и транслируется. Если транскрипции элемента не происходит (например, элемент дефектный) или происходит деградация транскрипта сразу после его синтеза (РНК- интерференция), то последующие процессы - образование кДНК и ее интеграция в геном, невозможны. Таким образом, транскрипция является одним из лимитирующих факторов в процессе перемещения ретроэлементов. В связи с этим, мы охарактеризовали нашу модельную систему – изогенные линии D . melanogaster 7К, 145 и С3, с точки зрения наличия или отсутствия транскриптов одиннадцати элементов группы Gypsy: rover , 17,6, 297, Idefix , Quasimodo , Transpac , Tirant , Zam , gypsy , springer , opus. Схема эксперименты анализа экспрессии показана на рисунке 3.3.
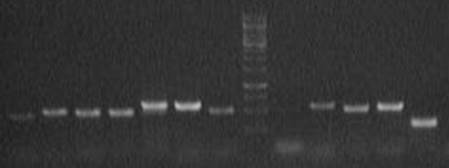
Рис.3.3. Схема эксперимента анализа экспрессии элементов. Результаты анализа экспрессии элементов представлены на 3.4.-3.9. В качестве референсного гена использовался ген rp49, который экспрессируется конститутивно.
Рис. 3.4. Результаты экспрессии МГЭ по самцам линии SS(7K)
Рис. 3.5. Результаты экспрессии МГЭ по самкам линии SS(7K)
Рис. 3.6. Результаты экспрессии МГЭ по самкам линии SS(С3(19))
Рис. 3.7. Результаты экспрессии МГЭ по самцам линии SS(С3(19))
Рис. 3.8. Результаты экспрессии МГЭ по самцам(7) линии MS(145)
Рис. 3.9. Результаты экспрессии МГЭ по самкам(8) линии MS(145) Элемент Zam во всех линиях и элемент Tirant в линии SS(7K) очень слабо экспрессировались, транскрипты остальных элементов детектировались достаточно уверенно. Наличие РНК элементов говорит об их гипотетической способност к транспозиции. 3.3. Анализ транспозиции МГЭ группы Gypsy в линиях D . melanogaser . Для дальнейшей характеристики изучаемых линий была проанализирована транспозиционная активность в линиях, мутантных по гену flamenco: MS (145) и SS (7K). В эксперименте изучались 11 МГЭ, представленных на рисунке 3.2. Эксперимент проходил в два этапа. Схема первого этапа эксперимента изображена на рисунке 3.10.
Рисунок 3.10. Схема эксперимента анализа трансозиции МГЭ (этап I). В результате для элементов: Zam , Idefix, rover, springer, gypsy, 17,6 (см. Приложение 1) в изогенных линиях MS(145) и SS(7K) были получены отличающиеся ПЦР – фрагменты. Для подтверждения специфичности продуктов был поставлен второй этап эксперимента: Саузерн- блоттинг (рис.3.11.).
Рис.3.11. Схема эксперимента анализа транспозиции МГЭ (этап II – Саузерн-блоттинг) Специфичность ПЦР- фрагментов была подтверждена для элементов : rover (см. рис. 3.12., 3.13.), springer (3.14.), gypsy (рис.3.15.).
Рис. 3.12. Результаты анализа транспозиции по элементу rover, полученные с использованием рестриктазы SmaI
Рис. 3.13. Результаты анализа транспозиции по элементу rover, полученные с использованием рестриктазы KpnI Как видно из рисунков 3.6. и 3.7. в изогенных линиях MS(145) и SS (7K) имеются различные ПЦР – фрагменты, то есть копии элемента rover, имеющие различное геномное окружение. На рисунке 3.6. в линии MS(145) ПЦР-фрагмент размером 3000 п.н., отсутствующий в линии SS (7K), а на рисунке 3.7. в линии MS два ПЦР- фрагмента, размерами 2000 п.н. и 1600 п.н. Это говорит о том, что элемент(ы) rover перемещались по геному, то есть о том, что они имеют транспозиционную активность в этих линиях.
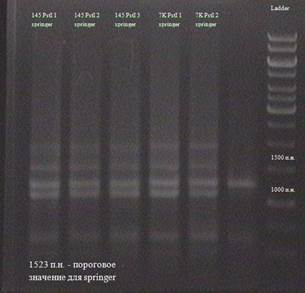
Рис. 3.14. Результаты анализа транспозиции по элементу springer, полученные с использованием рестриктазы PstI Как видно из рисунка 3.8. в линии SS(7K) присутствует ПЦР-фрагмент, размером около 8000 п.н., отсутствующий в линии MS(145). Это указывает на то, что элемент springer имеют транспозиционную активность в этих линиях.
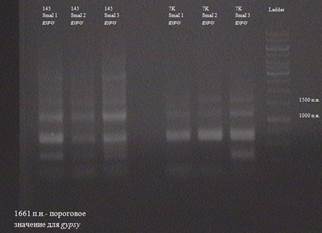
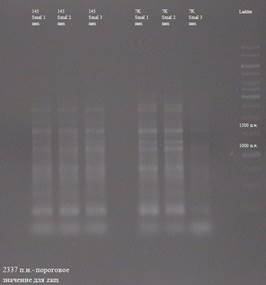
Рис. 3.15. Результаты анализа транспозиции по элементу gypsy, полученные с использованием рестриктазы SmaI Как видно из рисунка 3.9., в линии MS (145) присутствует ПЦР-фрагмент размером около 3000 п.н., отутствующий в линии SS (7K). Эти линии изогенны, но отличаются в наличие в линии MS(145) активной формы gypsy. Поэтому данный результат лишь подтверждает тот факт, что в линии SS (145) gypsy перемещается. Для остальных элементо (Idefix , Zam , 17,6, 297, opus , tirant , transpac , quasimodo) таким способом не получилось показать наличие транспозиционной активности в этих линиях. Однако, это не означает, что данные события не происходят, либо происходят крайне редко, и в рамках используемого подхода не обнаруживабтся. ВЫВОДЫ СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ ПРИЛОЖЕНИЕ 1 Результаты анализа транспозиции МГЭ (этап I ) В результатах присутствует термин «пороговое значение». Это означает, что анализировались ПЦР-фрагменты, имеющее значение большее, нежели пороговое, так как, как видно из рисунка 3.4. праймеры, были подобранны таким образом, что часть амплифицированной ДНК захватывала область элемента и именно эта область и является пороговым значением (измеряется в п.н.). На рисунках линия MS (145) обозначается как 145 линия, линия SS (7K) как 7K. Эксперимент ставился в 3-х проворностях. Результаты по элементу Idefix.

Результаты по элементу 17,6.
Результаты по элементу rover.
Результаты по элементу springer.
Результаты по элементу gypsy.
Результаты по элементу Zam .
ПРИЛОЖЕНИЕ 2. Продолжительность жизни каждой мухи.
Экологическая генетика. Экологическая генетика как наука образовалась во второй половине XX века. В 60-е годы Е.Б.Форд впервые сформулировал понятие «экологическая генетика» как генетику популяций животных в природных условиях (Ford, 1964). Экологическая генетика - междисциплинарная наука, вобравшая в себя закономерности и постулаты двух базовых дисциплин – экологии и генетики. Э. Геккель во второй половине XIX века определил экологию как науку об отношениях организмов с окружающей средой и с другими организмами (Гекель, 1866). Это определение охватывает не только область экологии, но и область смежных наук, в том числе и экологической генетики. Генетика, как определил ее У. Бэтсон в 1906 г.,- это наука о закономерностях наследственности и изменчивости. Таким образом, экологическая генетика - это область знания, использующая взаимовлияния генетических процессов и экологических отношений (Инге - Вечтомов, 2009). Область знаний, охватываемая экологической генетикой, представлена в табл.1.1. Табл. 1.1 Общая структура экологической генетики (Инге-Вечтомов и др., 1999).
Экологическая генетика опирается на мощную методологию генетического анализа и на методический арсенал экологии. Одним из вопросов экологической генетики является изучение генетического контроля устойчивости особей. Каждый организм в течение своей жизни подвергается воздействию неблагоприятных экологических факторов, таких как резкие перепады температуры, недостаток питания, гипероксия или гипоксия, поллютанты, ультрафиолетовое и ионизирующее излучение и др. Для перенесения неблагоприятных условий у живых организмов сформировались различные приспособления как на уровне индивида в течение онтогенеза (физиологическая адаптация), так и в ходе эволюции на уровне вида (генетическая адаптация). В последние годы, в связи с ухудшением состояния окружающей среды и возросшей на этом фоне заболеваемости человека, активно исследуются заболевания, развитию которых способствуют, с одной стороны, неблагоприятные факторы среды, а с другой стороны - неодинаковая реакция разных особей на данные факторы, обусловленная их генотипом. Было показано, что многие заболевания, например астма (Polonicov et al., 2009), колоректальный рак (Mala et al., 2009), папиллярный рак щитовидной железы (Siraj et al., 2008) и другие ассоциированы с некоторыми полиморфными вариантами генов ферментов трансформации ксенобиотиков. Биотрансформация ксенобиотиков – многостадийный процесс, и, соответственно, контролируется полигенно, в связи с чем, большое значение имеет исследование взаимодействия разных аллельных вариантов генов, участвующих в данном процессе. Важно отметить, что некоторые полиморфные варианты рассматриваемых генов могут иметь приспособительный характер. Например, частота аллеля 432L гена CYP1B1, выше у здоровых людей, по сравнению с людьми, имеющими бронхиальную астму. А отсутствие аллеля 462IV гена CYP1A1 приводит к увеличению вероятности возникновения астмы у курящих людей, тогда как у некурящих такого эффекта не наблюдается. Важно отметить, что один и тот же полиморфизм гена в разном генетическом окружении может по-разному влиять, оказывая как положительный, так и отрицательный эффект. Это объясняется взаимодействием генов. Например, взаимодействие аллеля Y113H гена EPHX1и аллеля V432L гена CYP1B1 приводит к увеличению вероятности развития заболевания (Polonicov et al., 2009). Другой яркий пример наследственной устойчивости к факторам окружающей среды – невосприимчивость отдельных индивидуумов к вирусу иммунодефицита человека (ВИЧ-1). Это обусловлено аллелем CCR5-Δ32 гена CC-CKR-5, в результате работы которого образуется нефункциональный белок – рецептор, вследствие чего, вирус иммунодефицита человека теряет способность связываться с рецептором и проникать в клетку (Liu et al., 1997; Djin-Ye Oh et al., 2008). Таким образом, генотип влияет на приспособленность организмов к окружающей среде. В борьбе за существование выживают более приспособленные особи. Менее приспособленные особи гибнут, либо не размножаются, либо производят на свет меньше полноценных потомков (Северцов, 2005). Характеристикой приспособленности может являться продолжительность жизни особи. Логично предположить, что чем выше продолжительность жизни, тем больше потомства может оставить особь. Продолжительность жизни является интегральным показателем стрессоустойчивости организма, так как она зависит от генотипа особи и от влияния экологических стрессоров (Петин, Сынзыныс, 1998).Поэтому, более узкой проблемой экологической генетики можно считать изучение влияния генотипа на продолжительность жизни модельных объектов. Продолжительность жизни. Продолжительность жизни - это мультифакторный признак, один из наиболее значимых количественных признаков организма. Старение с точки зрения биологии - это процесс постепенного угнетения основных функций организма, в том числе регенерационных и репродуктивных, вследствие чего организм становится менее приспособленным к условиям окружающей среды (теряется способность противостоять стрессам, болезням, травмам), что делает гибель организма неизбежной. Биология продолжительности жизни - это раздел биологии, занимающийся изучениемзакономерностей длительности жизни организмов, а также механизмов ее определяющих (Гаврилов и др.,1991). Среди наиболее актуальных ее проблем, выделяют: проблему наследуемости и изменчивости длительности жизни (Lints et al, 1989), проблему половых различий по срокам жизни (Hazzard, 1986), а также проблему изменения продолжительности жизни в процессе биологической эволюции (Wodhead, 1987). Таким образом, в задачи биологии продолжительности жизни входит выяснение причин индивидуальных различий по срокам жизни, а также причин межпопуляционных и межвидовых различий по этому признаку. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Последнее изменение этой страницы: 2019-04-21; Просмотров: 436; Нарушение авторского права страницы
 Денатурация ДНК: Т=95оС, 2 мин
Денатурация ДНК: Т=95оС, 2 мин