|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Факторы, влияющие на продолжительность жизни.
1) Экологические факторы. Экологические факторы, влияющие на продолжительность жизни, делятся на две группы: абиотические факторы и биологические факторы. Абиотическими факторами называются факторы неживой природы, к которым относятся: лучистая энергия, освещенность, температура, влажность воздуха, атмосферное давление, магнитное поле Земли, ионизирующее излучение, рельеф местности. В настоящее время активно изучается влияние на продолжительность жизни таких факторов, как температура (Helfand, Rogina, 2003; Huang et al., 2004), качество пищи (Helfand, Rogina 2003;Cheng et al., 2005; Partridge et al., 2005), ионизирующая радиация (Moskalev et al., 2008) Под биологическими факторами, влияющими на продолжительность жизни, подразумевают факторы живой природы, например: патогенные микроорганизмы (вирусы, бактерии, грибы, простейшие), хищники и т.д. (Стожанов, 2007). Например, показано, что при добавление в пищу гриба шиитаке Lentinula edodes увеличивается средняя продолжительность жизни Drosophila melanogaster (Матюшкова и др., 2010). 2) Генетические факторы. Генотип оказывает прямое влияние на жизнеспособность всех систем организма и на продолжительность его жизни. В последние годы, достигнут большой прогресс в понимании генетического контроля продолжительности жизни. Установлено, что в регуляции стрессоустойчивости организма, скорости его старения и продолжительности жизни участвуют различные гены. Многие из них вовлечены в процессы развития и роста клеток, метаболизма и репродукции, транскрипции и трансляции, и в другие жизненно важные процессы в клетке (McElwee et al., 2004; Lamitina, Strange, 2005; Carter, Brunet, 2007).Например, мутации в генах протеинкиназ PI 3 K, PKB, SGK-1, TOR или сверхэкспрессия фосфотазы PTEN, блокирующей каскад этих киназ, продлевают жизнь модельным объектам (Broughton et al., 2005; Solari et al, 2005). А деацетилазы белков семейства Sir2/ SIRT 1 в ответ на стрессовые воздействия подавляют проапоптозную функцию транскрипционных факторов p53 и FOXO, а также репрессируют гены, контролирующие участие эндоплазмотической сети в стресс-ответе, способствуя выживаемости клетки и увеличению продолжительности жизни (Rogina et al., 2004; Tissenbaum et al., 2006; Viswanathan et al.2005). А организм, в котором экспрессируется другая деацетилаза, Rpd3/ HDAC, напротив, способствует старению, а мутация ее гена продлевает жизнь (Rogina, 2002). В последнее время появились данные о влиянии генов белков теплового шока (Hsps, Heat Shock Proteins) на продолжительность жизни различных модельных организмов (Morrow et al., 2004; Klose et al., 2005; Poirier et al., 2008; Vanhooren et al., 2008). Продолжительность жизни может сильно отличаться в разных популяциях и у разных особей одного вида. Считается, что генетический контроль продолжительности жизни эволюционно консервативен, что делает целесообразным проведение исследований на модельных объектах, в том числе на дрозофиле, большинство генов которой имеет ортологи у других высших эукариот.
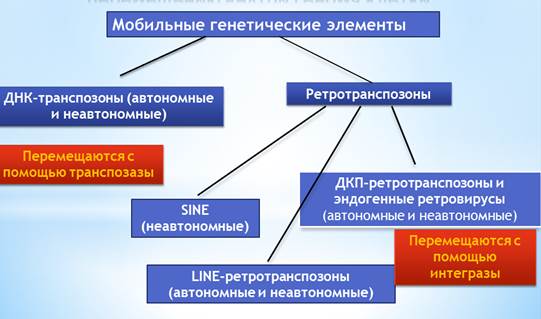
1.3. Drosophila melanogaster как модельный объект. Drosophila melanogaster, иначе плодовая, или уксусная, муха, принадлежит к семейству Drosophilidae из отряда Diptera. Это маленькая мушка, величиной около 3 мм. D . melanogaster является идеальной моделью для исследования разных вопросов генетики, так как обладает следующими свойствами: 1. Имеет малый срок развития от яйца до взрослой мухи. 2.Имаго имеет относительно не большую продолжительность жизни (при температуре 25° С около двух месяцев). 3. Обладает большой плодовитостью. 4. Имеет всего четыре хромосомы. 5. Имеет большое количество генов - ортологов с другими высшими эукариотами. На продолжительность жизни D . melanogaster влияют условия содержания: температура, влажность, пища, плотность популяции, наличие в питательной среде дрожжей. В некоторых опытах D . melanogaster доживала до 153 дней. Были получены разнообразные мутантные формы D . melanogaster обладающие меньшей или большей продолжительностью жизни по сравнению с диким типом (Медведев, 1965). На продолжительность жизни D . melanogaster влияют различные генетические факторы. Например, было показано влияние генов, кодирующих белки, необходимые для биосинтеза катехоламинов и передачи нервного импульса в нейронах, и генов, кодирующих транскрипционные факторы РНК-полимеразы II, которые участвуют в контроле развития и функционирования мотонейронов, на продолжительность жизни D . melanogaster (Рощина, 2007). Значительную часть генома D . melanogaster составляют мобильные генетические элементы (МГЭ). МГЭ могут существенно влиять на генетический материал хозяина. Перемещаясь внутри генома, они могут вызывать различные мутации и хромосомные аберрации (McClintock, 1953). Поскольку, МГЭ в результате своей активности могут нарушить работу различных генов, они могут рассматриваться как потенциальный фактор, влияющий на продолжительность жизни. 1.4. Мобильные генетические элементы Drosophila melanogaster и ретровирус gypsy . 1.4.1. Мобильные генетические элементы. Мобильные генетические элементы (МГЭ) представляют собой последовательности ДНК, способные перемещаться по геному. МГЭ имеют широкое распространение и составляют большую часть геномной ДНК многих изученных организмов. Впервые мобильные элементы были обнаружены в 1940-х годах американской исследовательницей Барбарой МакКлинток при изучении локуса или гена кукурузы, вызывавшего повышенные частоты хромосомных перестроек, индуцируя мутантный фенотип (McClintock, 1953). В конце 60-х годов М. Грин обнаружил высокомутабильный аллель гена white D . melanogaster, который с частотой 1 х 10–3 давал реверсии к норме, а также мутировал к промежуточным по фенотипу (цвет глаз) алеллям (Green, 1969). А в конце 1970-х у D . melanogaster были открыты разные семейства мобильных элементов (Rubin et al.,1976; Finnegan et al.,1978; Георгиев, Гвоздев, 1980).К настоящему времени известно множество генов, причинной мутации в которых является транспозиции МГЭ (Голубовский, 2009). МГЭ могут играть важную роль в функционировании клетки. Некоторые МГЭ, такие как HETA и TART стали незаменимыми в поддержание теломер у дрозофилы. SINE и LINE элементы участвуют в процессах репарации двунитевых разрывов ДНК у дрожжей (Kidwell М.J., Lisch D.,1997). МГЭ можно классифицировать по способу перемещения и по структуре. Выделяют элементы способные самостоятельно вырезаться и перемещаться, их называют автономные элементы; и те, которые не способны к самостоятельному перемещению – неавтономные. МГЭ разделяют по механизму транспозиции (рис.1.1): 1. Мобильные генетические элементы, перемещающиеся за счет фермента транспозазы. Транспозаза «вырезает» элемент из одной части генома и встраивает его в другую. Такие элементы называются транспозоны. Они в свою очередь разделяют на МГЭ с короткими инвертированными повторами (P элемент, hobo) и с длинными инвертированными повторами (FB-элементы). 2. Мобильные генетические элементы, использующие РНК-интермедиат. Сначала происходит транскрипция последовательности элемента, затем с помощью фермента РНК – зависимой - ДНК - полимеразы происходит синтез комплементарной ДНК. Потом интеграза встраивает новую копию элемента в геном. Такие элементы называются ретротранспозонами. Их также разделяют на две подгруппы: ретротранспозоны с длинными концевыми повторами (ДКП- ретротранспозоны), примером которых являются: copia, МДГ1, МДГ3, gypsy, и LINE элементы (поли-А-ретротранспозоны, не содержащие ДКП).
Рис.1.1. Классификация мобильных элементов. Объяснения в тексте. |
Последнее изменение этой страницы: 2019-04-21; Просмотров: 221; Нарушение авторского права страницы