|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Прежде всего, общая биологияСтр 1 из 25Следующая ⇒
Прежде всего, общая биология академик Академии медицинских наук СССР И.В. Давыдовский ВВЕДЕНИЕ БИОЛОГИЯ – НАУКА О ЖИВОЙ ПРИРОДЕ Биология (от лат. bios – жизнь, logos – учение) – наука, которая изучает жизнь как особую форму движения материи, законы ее существования и развития. Термин «биология» был впервые предложен французским естествоиспытателем Ж.-Б. Ламарком в 1802 г. для обозначения науки о жизни, как особом явлении природы. Биология зародилась в античное время (Гиппократ, Аристотель, Гален), однако свое название получила только в 1802 г., когда этот термин в современном его толковании был предложен французским ученым Ж.Б. Ламарком и немецким исследователем Г.Р. Тревиранусом. Первые сведения о живых существах человек начал собирать с техпор, как выделил себя из окружающей природы, осознавая свое отличие от ее объектов. Из сохранившихся литературных памятников известно, что уже древние индейцы, вавилоняне, египтяне и другие народы немало знали о растениях и животных. В XIV в. в Месопотамии систематизировались знания о растениях, подразделявшихся на деревья, овощи, лекарственные травы и т. п., а также о плотоядных и травоядных животных, что следует из дошедших до нас клинописных табличек тех времен. Изучение живой природы диктовалось двумя насущными потребностями человечества: потребностью познания растений и животных с целью удовлетворения своих нужд в растительной и животной пище и необходимостью познания тела человека с целью совершенствования древнейшего искусства врачевания. Основной задачей биологии является изучение общих закономерностей развития живой природы, раскрытие сущности жизни, систематизация живых существ. Предметом биологии являются строение и жизнедеятельность живых организмов, происхождение, развитие и распространение живых существ на Земле, их связи друг с другом и с неживой природой. В развитии биологии условно выделяют три основных этапа: Первый этап – разработка классификации и систематика живого. Большой вклад в развитие биологии внес Карл Линней, предложивший систему классификации животных и растений, Карл Максимович Бэр (1792–1876) в своих работах сформулировал основные положения теории гомологичных органов и закона зародышевого сходства, заложившие научные основы эмбриологии, в 1808 г. в работе «Философия зоологии» Жан Батист Ламарк поставил вопрос о причинах и механизмах эволюционных преобразований и изложил первую по времени теорию эволюции. Второй этап – становление эволюционного учения. На основе многочисленных наблюдений Ч. Дарвин опубликовал в 1859 г. свой основной труд «О происхождении видов путем естественного отбора или сохранении благоприятствуемых пород в борьбе за жизнь», в котором сформулировал основные положения теории эволюции, предложил механизмы эволюции и пути эволюционных преобразований организмов. Третий этап – развитие микробиологии. В XIX в. благодаря работам Луи Пастера (1822–895), Роберта Коха (1843–910), Ильи Ильича Мечникова в качестве самостоятельной науки оформилась микробиология. К концу XIX в. как отдельные науки выделились паразитология и экология. XX век начался с переоткрытия законов Грегора Менделя, что ознаменовало собой начало развития генетики как науки. В 40–50-е годы XX в. в биологии стали широко использоваться идеи и методы физики, химии, математики, кибернетики и других наук, а в качестве объектов исследования – микроорганизмы. В результате возникли и стали бурно развиваться как самостоятельные науки биофизика, биохимия, молекулярная биология, радиационная биология, бионика и др. Исследования в космосе способствовали зарождению и развитию космической биологии. В XX в. появилось направление прикладных исследований – биотехнология. В настоящее время условно можно выделить три направления в биологии. Первое направление – это классическая биология. Ее представляют ученые-натуралисты, изучающие многообразие живой природы. Они объективно наблюдают и анализируют все, что происходит в живой природе, изучают живые организмы и классифицируют их. Второе направление – это эволюционная биология. В настоящее время изучение эволюции живых организмов активно продолжается. Синтез генетики и эволюционной теории привел к созданию так называемой синтетической теории эволюции. Но и сейчас еще есть много нерешенных вопросов, ответы на которые ищут ученые-эволюционисты. Третье направление – физико-химическая биология, исследующая строение живых объектов при помощи современных физических и химических методов. Это быстро развивающееся направление биологии, важное как в теоретическом, так и в практическом отношении. Можно с уверенностью говорить, что в физико-химической биологии нас ждут новые открытия, которые позволят решить многие проблемы, стоящие перед человечеством. В настоящее время биологические знания используются во всех сферах человеческой деятельности: в промышленности и сельском хозяйстве, медицине и энергетике. Чрезвычайно важное значение имеют экологические исследования. Перед человечеством встала грандиозная задача – сохранение биосферы с целью поддержания условий существования и развития цивилизации. Без биологических знаний и специальных исследований решить ее невозможно. Современная биология представляет собой комплекс естественных наук, изучающих живую природу. Так, биология тесно связана с химией, физикой, математикой и другими науками. На стыке наук получили развитие биохимия, биофизика, физиология и другие. Биология является теоретической базой медицинской генетики, микробиологии, иммунологии, паразитологии, непосредственно связанных с медициной. Все биологические науки можно подразделить на следующие группы: - общебиологические (цитология, генетика, эволюционное учение и др.); - морфологические дисциплины (например: анатомия, гистология, патологическая анатомия); - физиологические (физиология растений, животных, нормальная физиология, патологическая физиология); - экологические (экология, биогеография, паразитология); - пограничные (биохимия, биофизика, молекулярная биология и др.). ОСНОВНЫЕ СВОЙСТВА ЖИВОГО Фундаментальные свойства живых организмов: саморегуляция, самообновление, самовоспроизведение. Живое характеризуется целым рядом важнейших признаков: 1. Обмен веществ, энергии и информации. Любой живой организм можно представить как открытую систему, поддерживающую непрерывный обмен веществ и энергии с окружающей средой. Основу обменных процессов составляют реакции пластического (анаболизм) и энергетического обмена (катаболизм). По типу анаболизма живые организмы подразделяют на автотрофные (фотосинтезирующие и хемосинтезирующие), гетеротрофные (сапротрофные и паразиты), миксотрофные. По типу катаболизма организмы бывают аэробными и анаэробными. 2. Структурная организация. Живое построено из тех же химических элементов, что и неживое, но характеризуется сложностью химических соединений, обусловленной определенной упорядоченностью на молекулярном уровне. Структурная организация – характерное свойство живого на всех уровнях его организации. Типичный пример упорядоченной структуры – хромосома (единство нуклеиновых кислот и белков). 3. Дискретность и целостность. Органический мир целостен, т.к. составляет систему взаимосвязанных частей, и в то же время он дискретен (лат. discretus – прерывистый). Органический мир состоит из отдельных единиц – организмов или особей. Каждый организм состоит из клеток, но функционирует как единое целое. 4. Репродукция – воспроизведение себе подобного. 5. Наследственность и изменчивость – важнейшие признаки живого, связанные с передачей потомству от родителей наследственных признаков организма и с возможностью их изменяться под влиянием факторов среды. 6. Рост и развитие – признаки организма, обеспечивающие увеличение размеров и развитие за счет деления клеток и их дифференцировки. 7. Раздражимость и движение. Признаки живого, благодаря которым организмы непрерывно контактируют с окружающей средой, другими организмами. У одноклеточных – в виде таксисов, у растений – в виде тропизмов, у высших животных – в виде рефлексов. 8. Внутренняя регуляция и гомеостаз. Любой организм, являясь открытой системой, сохраняет в тоже время постоянство своей внутренней среды (гомеостаз) благодаря нейрогуморальной регуляции гомеостаза. ПРОИСХОЖДЕНИЕ ЖИЗНИ
Существуют две главные гипотезы, по-разному объясняющие появление жизни на Земле. Согласно гипотезе панспермии, жизнь занесена из космоса либо в виде спор микроорганизмов, либо путем намеренного «заселения» планеты разумными пришельцами из других миров. Прямых свидетельств в пользу космического происхождения жизни нет. Космос, однако, наряду с вулканами мог быть источником низкомолекулярных органических соединений, раствор которых послужил средой для развития жизни. Современной наукой возраст Земли оценивается в 4, 5—4, 6 млрд. лет. Появление на планете первых водоемов, с которыми связывают зарождение жизни, отстоит от настоящего времени на 3, 8—4 млрд. лет. Полагают, что около 3, 8 млрд. лет назад жизнь могла стать определяющим фактором планетарного круговорота углерода. В породах вблизи местечка Фиг-Три (Южная Африка), имеющих возраст более 3, 5 млрд. лет, обнаружены бесспорные следы жизнедеятельности микроорганизмов. Таким образом, процесс образования примитивных живых существ шел относительно быстро. Ускорению процесса могло способствовать то, что простейшие органические вещества были из нескольких источников: абиогенно образующиеся в первичной атмосфере и в то же время поступающие с оседающей на поверхность планеты космической и вулканической пылью. Подсчитано, что Земля, проходя через пылевое облако в течение 1 млрд. лет, могла получить с космической пылью 10 млрд. т органического материала. Это всего в 300 раз меньше суммарной биомассы современных наземных организмов (3 • 1012 т). Вулкан за одно извержение выбрасывает до 1000 т органических веществ. Согласно второй гипотезе, жизнь возникла на Земле, когда сложилась благоприятная совокупность физических и химических условий, сделавших возможным абиогенное образование органических веществ из неорганических. В середине прошлого столетия Л. Пастер окончательно доказал невозможность самозарождения жизни в теперешних условиях. В 20-х годах текущего столетия биохимики А. И. Опарин и Дж. Холдейн предположили, что в условиях, имевших место на планете несколько миллиардов лет назад, образование живого вещества было возможно. К таким условиям они относили наличие атмосферы восстановительного типа, воды, источников энергии (в виде ультрафиолетового (УФ) и космического излучения, теплоты остывающей земной коры, вулканической деятельности, атмосферных электрических явлений, радиоактивного распада), приемлемой температуры, а также отсутствие других живых существ. Главные этапы на пути возникновения и развития жизни, по-видимому, состоят в: 1) образовании атмосферы из газов, которые могли бы служить «сырьем» для синтеза органических веществ (метана, оксида и диоксида углерода, аммиака, сероводорода, цианистых соединений), и паров воды; 2) абиогенном (т.е. происходящем без участия организмов) образовании простых органических веществ, в том числе мономеров биологических полимеров — аминокислот, Сахаров, азотистых оснований, АТФ и других мононуклеотидов; 3) полимеризации мономеров в биологические полимеры, прежде всего белки (полипептиды) и нуклеиновые кислоты (полинуклеотиды); 4) образовании предбиологических форм сложного химического состава — протобионтов, имеющих некоторые свойства живых существ; 5) возникновении простейших живых форм, имеющих всю совокупность главных свойств жизни, — примитивных клеток; 6) биологической эволюции возникших живых существ. Возможность абиогенного образования органических веществ, включая мономеры биологических полимеров, в условиях, бывших на Земле около 4 млрд. лет назад, доказана опытами химиков. В лабораторных условиях при пропускании электрических разрядов через различные газовые смеси, напоминающие примитивную атмосферу планеты, а также при использовании других источников энергии ученые получали среди продуктов реакций аминокислоты (аланин, глицин, аспарагиновую кислоту), янтарную, уксусную, молочную кислоты, мочевину, азотистые основания (аденин, гуанин), АДФ и АТФ. Низкомолекулярные органические соединения накапливались в водах первичного океана в виде первичного бульона или же адсорбировались на поверхности глинистых отложений. Последнее повышало концентрацию этих веществ, создавая тем самым лучшие условия для полимеризации. Возможность полимеризации низкомолекулярных соединений с образованием полипептидов и полинуклеотидов (определяющая следующий этап на пути возникновения жизни) непосредственно в первичном бульоне вызывает сомнения по термодинамическим соображениям. Водная среда благоприятствует реакции деполимеризации. Ученые предполагают, что образование полипептидов и полинуклеотидов могло происходить в пленке из низкомолекулярных органических соединений в безводной среде, например на склонах вулканических конусов, покрытых остывающей лавой. Это предположение находит подтверждение в опытах. Выдерживание в течение определенного времени при 130°С сухой смеси аминокислот в сосудах из кусков лавы приводило к образованию полипептидов. Образующиеся описанным образом биополимеры смывались ливневыми потоками в первичный бульон, что защищало их от разрушающего действия УФ-излучения, которое в то время из-за отсутствия в атмосфере планеты озонового слоя было очень жестким. По мере повышения концентрации полипептидов, полинуклеотидов и других органических соединений в первичном бульоне сложились условия для следующего этапа — самопроизвольного возникновения предбиологических форм сложного химического состава, или протобионтов. Предположительно они могли быть представлены коацерватами (А. И. Опарин) или микросферами (С. Фоке). Это коллоидные капли с уплотненным поверхностным слоем, имитирующим мембрану, содержимое которых составляли один или несколько видов биополимеров. Возможность образования в коллоидных растворах структур типа коацерватов или микросфер доказана опытным путем. При определенных условиях коацерваты проявляют некоторые общие свойства живых форм. Они способны до известной степени избирательно поглощать вещества из окружающего раствора. Часть продуктов химических реакций, проходящих в коацерватах с участием поглощаемых веществ, выделяется ими обратно в среду. Происходит процесс, напоминающий обмен веществ. Накапливая вещества, коацерваты увеличивают свой объем (рост). По достижении определенных размеров они распадаются на части, сохраняя при этом некоторые черты исходной химической организации (размножение). Поскольку устойчивость коацерватов различного химического состава различна, среди них происходит отбор. Перечисленные выше свойства ученые усматривают у протобионтов. Протобионты представляются как обособленные от окружающей среды, открытые макромолекулярные системы, возникавшие в первичном бульоне и способные к примитивным формам роста, размножения, обмена веществ и предбиологическому химическому отбору. Предбиологическая эволюция протобионтов осуществлялась в трех главных направлениях. Важное значение имело совершенствование каталитической (ферментной) функции белков. Один из путей, дающих требуемый результат, заключается, по-видимому, в образовании комплексов металлов с органическими молекулами. Так, включение железа в порфириновое кольцо Гемоглобина увеличивает его каталитическую активность в сравнении с активностью самого железа в растворе в 1000 раз. Развивалось такое свойство биологического катализа, как специфичность. Во-вторых, исключительная роль в эволюции протобионтов принадлежит приобретению полинуклеотидами способности к самовоспроизведению, что сделало возможным передачу информации от поколения к поколению, т.е. сохранение ее во времени. В основе этой способности лежит матричный синтез. Механизм матричного синтеза был использован также для переноса информации с полинуклеотидов на полипептиды. Третье главное направление эволюции протобионтов состояло в возникновении мембран. Отграничение от окружающей среды мембраной с избирательной проницаемостью превращает протобионт в устойчивый набор макромолекул, стабилизирует важные параметры обмена веществ на основе специфического катализа. Разделение функций хранения и пространственно-временной передачи информации, с одной стороны (нуклеиновые кислоты), и использование ее для организации специфических структуры и обмена веществ — с другой (белки); появление молекулярного механизма матричного синтеза биополимеров; освоение эффективных систем энергообеспечения жизнедеятельности (АТФ); образование типичной биологической мембраны — все это привело к возникновению живых существ, которые поначалу были представлены примитивными клетками. С момента появления клеток предбиологический химический отбор уступил место биологическому отбору. Дальнейшее развитие жизни шло согласно законам биологической эволюции. Переломным моментом на этом пути было возникновение клеток эукариотического типа, многоклеточных организмов, человека.
РАЗДЕЛ I. ЦИТОЛОГИЯ ЖИВОГО КЛЕТОЧНЫЙ (ЖИЗНЕННЫЙ) ЦИКЛ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ Жизненные формы организмов: 1. Доклеточные – царство вирусов. 2. Клеточные: прокариоты – царства бактерий и цианобактерий, эукариоты – царства растений, животных и грибов. Основными отличиями строения и жизнедеятельности прокариотических клеток от эукариотических клеток являются следующие (рис. 1):
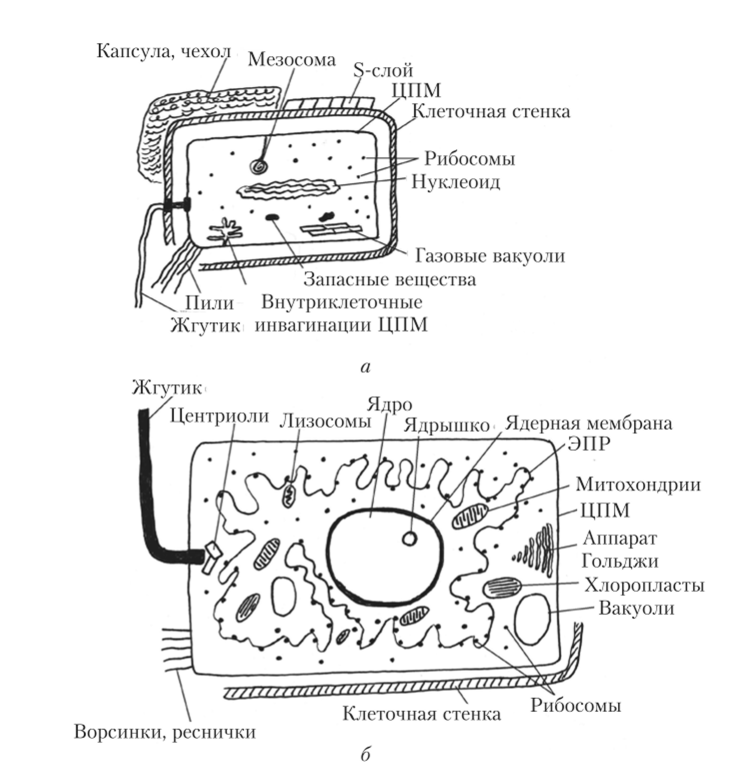
Рис. 1. Отличие эукариот от прокариот: а - клетка прокариот, б - клетка эукариот
1. Клетка прокариот не имеет оформленного (ограниченного мембраной) ядра, наследственная информация в ней содержится в кольцевой молекуле ДНК. ДНК не заблокирована белками, в первую очередь гистонами, поэтому все гены в ней активны, т. е. постоянно функционируют. У эукариотических клеток имеется оформленное ядро, а генетический аппарат представлен молекулами ДНК в комплексе с белками – гистонами, упаковывающими ДНК в компактные структуры и регулирующими активность ее генов. 2. Цитоплазма прокариотической и эукариотической клеток окружена мембраной (плазмолеммой), однако у бактерий, растений и грибов снаружи от плазмолеммы располагается клеточная стенка, образованная веществом полисахаридной природы – муреином (бактерии), целлюлозой (растения) или хитином (грибы). Клеточная оболочка животной клетки образована плазмолеммой, покрытой снаружи слоем гликокаликса. 3. В цитоплазме прокариотической клетки отсутствуют мембранные органеллы (митохондрии, пластиды, эндоплазматическая сеть, пластинчатый комплекс, лизосомы, пероксисомы), а ограниченное количество мембран представляет собой впячивания плазмолеммы внутрь цитоплазмы – мезосому. 4. Синтез белка осуществляется свободными рибосомами, имеющими меньший размер (70S), чем рибосомы эукариотнческих клеток (80S). Большая субъединица рибосомы прокариотической клетки содержит 2 молекулы рибосомной РНК (рРНК), тогда как субъединица рибосомы эукариотической клетки – 3 молекулы рРНК. 5. Специальные органеллы прокариотической клетки – жгутики устроены проще, чем жгутики эукариотической клетки: они лишены внутреннего каркаса из микротрубочек и микрофиламентов. 6. В цитоплазме многих прокариотических клеток имеются газовые вакуоли. 7. В прокариотических клетках отсутствует клеточный центр.
8. Прокариоты размножаются простым делением клетки, у эукариот имеет место половой процесс с образованием гамет. 9. У прокариотических клеток отсутствует амебоидное движение и внутриклеточные перемещения цитоплазмы. 10. Синтез АТФ осуществляется в прокариотических клетках на мембране плазмолеммы. Эукариотические клетки имеют обособленное ядро, наружную биологическую мембрану – плазмолемму и цитоплазму с органеллами и включениями.
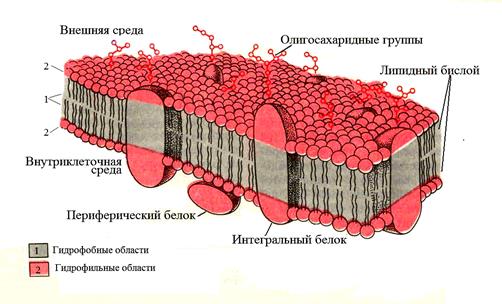
Цитоплазматическая мембрана Функции мембран: 1. Барьерная (отграничивающую); 2. Регуляторная (осуществляют регуляцию метаболических потоков); 3. Транспортную (обеспечение избирательной проницаемости веществ путем пассивного и активного транспорта); 3. С труктурная; 4. О бменная. Биологические мембраны построены в основном из липидов, белков и углеводов. Предложено несколько моделей строения цитоплазматических мембран (модель «сэндвича» - модель Даниели и Даусона, модель Ленарда и др.). По-видимому, в зависимости от функции существует несколько типов мембран. В настоящее время принята за основу жидкостно-мозаичная модель, предложенная Сингером-Николсоном в 1972 г. (рис. 3). Согласно этой модели в состав мембран входит бимолекулярный слой фосфолипидов, в который погружены молекулы белков.
Рис. 3. Модель Сингера-Николсона (жидкостно-мозаичная)
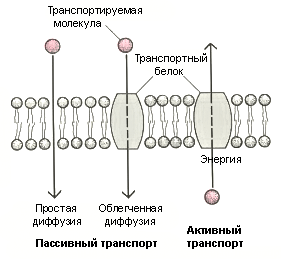
Фосфолипиды – соединения глицерина, жирных кислот и остатка фосфорной кислоты. Это водонерастворимые соединения, которые состоят из полярной (заряженной) головки (азот-содержащая группа) и двух длинных неполярных (незаряженных) хвостов (цепи жирных кислот). Молекулы липидов обращены друг к другу неполярными хвостами, а их полярные полюса (головки) остаются снаружи, образуя гидрофильные поверхности. В бимолекулярный слой липидов погружены белковые молекулы. Белки мембран можно разделить на три группы: периферические (наиболее слабо связаны с мембраной), погруженные (полуинтегральные) и пронизывающие (интегральные, трансмембранные), формирующие поры и каналы мембраны. В функциональном отношении белки мембран подразделяются на ферментативные, транспортные, структурные и регуляторные. На внешней поверхности плазматической мембраны белковые и липидные молекулы связаны с углеводными цепями (гликопротеиды и гликолипиды), образуя гликокаликс – рецепторный аппарат клетки. Гликопротеиды выполняют роль рецепторов, клетка приобретает способность специфически реагировать на воздействия извне. Так, взаимодействие гормона со «своим» рецептором снаружи вызывает изменение структуры интегрального белка, что приводит к запуску клеточного ответа. В частности, такой ответ может проявиться в образовании «каналов», по которым растворы некоторых веществ начинают поступать в клетку или выводятся из нее. Гликолипиды обеспечивают одну из важных функций мембраны – обеспечение межклеточных контактов. Под плазматической мембраной со стороны цитоплазмы имеются белковые фибриллы, формирующие опорно-сократительный аппарат клетки. У растительных клеток кнаружи от мембраны расположена плотная структура – клеточная оболочка, состоящая из полисахаридов (целлюлозы). Одно из важнейших свойств цитоплазмы связано со способностью пропускать в клетку или из нее различные вещества. Это необходимо для поддержания постоянства ее состава. Малые молекулы и ионы проходят через мембраны путем пассивного и активного транспорта. Пассивный транспорт происходит без затрат энергии путем свободной диффузии, облегченной диффузии и осмоса (рис. 4). Диффузия – транспорт молекул и ионов через мембрану из области с высокой в область с низкой их концентрацией, т.е. по градиенту концентрации. Если вещества хорошо растворимы в жирах, то они проникают в клетку путем простой диффузии (кислород, углекислый газ). Облегченная диффузия – транспорт веществ, нерастворимых в жирах и не проходящих сквозь поры, через ионные каналы с помощью белков переносчиков.
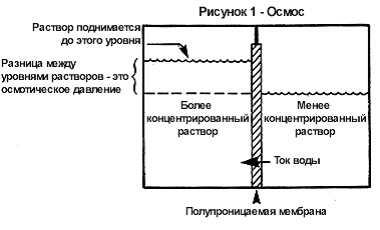
Рис. 4. Схема активного и пассивного транспорта веществ через мембрану Транспорт воды через полупроницаемые мембраны называется осмосом (рис. 5).
Рис. 5. Осмос
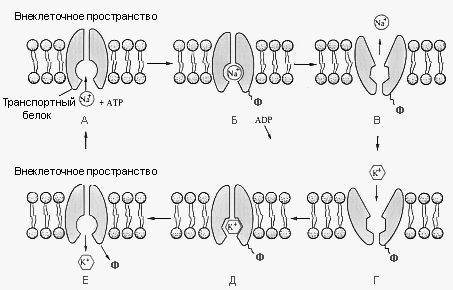
В цитоплазматической мембране присутствуют специальные каналы для транспортировки воды с растворенными в ней ионами и молекулами. В 1989 году американский ученый Питер. Это выделил мембранный белок, образующий водные каналы, и назвал аквапорином. В тканях человека было обнаружено 11 аналогов аквопорина, причем ряд из них может привести к появлению тех или иных заболеваний человека, например, к некоторым формам диабета и хронической сердечной недостаточности. Вода переходит из области с меньшей концентрацией солей в область, где их концентрация больше. Возникающее давление на полупроницаемую мембрану называют осмотическим. Напряженное состояние клеточной оболочки, создаваемое давлением внутриклеточной жидкости, называется тургором. Тургор обуславливается тремя факторами: внутренним осмотическим давлением клетки, которое вызывает напряжение клеточной оболочки, внешним осмотическим давлением, а также упругостью клеточной оболочки. Снижением тургора сопровождаются процессы обезвоживания, автолиза (распада), увядания и старения клеток. Активный транспорт веществ через мембрану осуществляется против градиента концентрации с затратой энергии АТФ и при участии белков-переносчиков. Так транспортируются аминокислоты, сахар, ионы калия, натрия, кальция и др. Примером активного транспорта может быть работа калий - натриевого насоса (рис. 6).
Рис. 6. Схема работы калий - натриевого насоса
Концентрация К+ внутри клетки в 10 – 20 раз выше, чем снаружи, а Na+– наоборот. Для поддержания данной концентрации происходит перенос трех ионов Na+ из клетки на каждые два иона К+ в клетку. В этом процессе участвует белок в мембране, выполняющий функцию фермента, расщепляющего АТФ с высвобождением энергии, необходимой для работы насоса. Перенос макромолекул и крупных частиц внутрь клетки осуществляется за счет эндоцитоза, а удаление из клетки - путем экзоцитоза (рис. 7).
Рис.7. Схема эндо- и экзоцитоза
При эндоцитозе мембрана образует впячивания или выросты, которые затем отшнуровываясь превращаются во внутриклеточные пузырьки, содержащие захваченный клеткой продукт. Этот процесс происходит с затратойэнергии АТФ. Различают два вида эндоцитоза – фагоцитоз (поглощение клеткой крупных частиц) и пиноцитоз (поглощение жидких веществ). Мембрана принимает участие в выведении веществ из клетки в процессе экзоцитоза. Таким способом из клетки выводятся гормоны, белки, жировые капли и др. Цитоплазма – внутреннее содержимое клетки без ядра, состоит из основного вещества, органелл и включений. Гиалоплазма (цитозоль) – основное вещество цитоплазмы, заполняющее пространство между клеточными органеллами. Гиалоплазма содержит около 90% воды и различные белки, аминокислоты, нуклеотиды, ионы неорганических соединений и др. Крупные молекулы белка образуют коллоидный раствор, который может переходить из золя (невязкое состояние) в гель (вязкий). В гиалоплазме протекают ферментативные реакции, метаболические процессы, синтез аминокислот, жирных кислот. Гиалоплазма содержит множество белковых нитей – филаментов, которые пронизывают цитоплазму и образуют цитоскелет. Включения - это относительно непостоянные (временные) компоненты цитоплазмы, которые не имеют мембраны и представляют собой продукты, подлежащие выведению из организма (секреторные (например, инсулин в клетках поджелудочной железы), экскреторные (например, мочевая и щавелевая кислоты)); запасные питательные вещества (гликоген, крахмал, белки, жиры, углеводы); пигменты (меланин, гемоглобин). Органеллы (органоиды) – постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции. В зависимости от функции различают органоиды общего и специального назначения. К органоидам специального назначения относятся микроворсинки, реснички, жгутики. Органеллы общего назначения делятся на немембранные (рибосомы, клеточный центр (центросома), микротрубочки, промежуточные филаменты, микрофиламенты) и мембранные. К одномембранным органеллам относятся эндоплазматическая сеть (ретикулум), аппарат Гольджи, лизосомы, пероксисомы, вакуоли. К двумембранным органеллам относятся митохондрии и пластиды растительных клеток. Органоиды специального назначения: реснички, жгутики и микроворсинки – органеллы передвижения. Представляют собой тонкие цилиндрические выросты цитоплазмы, покрытые плазматической мембраной. Жгутики отличаются от ресничек длиной. Микроворсинки формируются только на одной поверхности клетки.
РИБОСОМЫ Анаболическая (ассимиляция, пластический обмен) и катаболическая (диссимиляция, энергетический обмен) системы клетки неразрывно связаны, так как все процессы жизнедеятельности клетки немыслимы без энергии АТФ, которая, в свою очередь, не может образовываться без ферментных систем, строящихся в результате анаболических реакций. Также неразрывно связаны друг с другом потоки вещества и энергии, так как гетеротрофные клетки способны использовать только энергию, заключенную в сложных химических соединениях. К анаболической системе клетки относятся: рибосомы, эндоплазматический ретикулум, комплекс Гольджи.
КЛЕТОЧНЫЙ (ЖИЗНЕННЫЙ) ЦИКЛ Период времени от деления материнской клетки до собственного деления дочерней или ее гибели называют жизненным циклом (или клеточным). Клеточный цикл (КЦ) - э то период существования клетки от образования до следующего деления или гибели (рис. 9). В КЦ различают две фазы: автосинтетическую, или интерфазу (подготовка клетки к делению) включающую пресинтетический (G1, от англ. Gap – промежуток), синтетический – (S), постсинтетический – (G2) периоды, и деление клетки – митоз. Выход клетки из цикла обозначается стадией G0. Так называемый период покоя. G0 – фаза принятия решения. Будет ли клетка размножаться и через какое-то время такие клетки могут вновь вернуться, но чаще всего в многоклеточных организмах такие клетки теряют способность к размножению. Слабо дифференцированные клетки сохраняют способность к делению и продолжают следовать по G1, S, G2 периодам и заканчивается цикл делением (митозом, или мейозом). Пресинтетический (G1) период. В этот период клетки растут, синтезируют РНК, белки, накапливается энергия, но синтез ДНК отсутствует. Период характеризуется тем, что клетка содержит диплоидный набор хромосом (2n). Каждая хромосома образована одной хроматидой. Хромосомы деспирализованы. Длительность периода – 8–10 часов. Если количество ДНК, содержащееся в гаплоидном наборе хромосом человека обозначить " С", то в G1 количественная характеристика генетического материала: 2n 1 хроматида 2С.
Рис. 9. Жизненный цикл G1 – пресинтетический период, S – синтетический период, G2 – постинтетический период, G0 – период пролиферативного покоя Cинтетический (S) период. В клетке продолжается транскрипция РНК, синтезируются гистоновые белки, удваиваются центриоли клеточного центра. Основной процесс, который происходит в ядре – это репликация ДНК, в результате которого происходит удвоение генетического материала. Набор генетического материала 2n 2 хроматиды 4С (диплоидный набор двухроматидных хромосом – двунитевых хромосом). Длительность этого периода 4–8 часов. СПОСОБЫ РЕПРОДУКЦИИ Типы и виды деления клеток
ПРЯМОЕ | |||||
| МИТОЗ |
АМИТОЗ | ||||
| 1.Собственно митоз | по форме | по виду | |||
| 2. Эндомитоз | 1.Равномерный | 1.Генеративный | |||
| 3. Политения | 2.Неравномерный | 2.Реактивный | |||
| 4. Мейоз | 3.Фрагментация | 3.Дегенеративный | |||
| 4.Без деления цитоплазмы | |||||
В процессе митоза идет последовательная смена фаз: профаза, метафаза, анафаза, телофаза (рис. 10).
Профаза – происходит спирализация, укорочение, утолщение хроматиновых нитей. Наблюдается расхождение центриолей к полюсам. Начинают образовываться нити веретена деления. В конце – наблюдается разрушение ядрышка и ядерной оболочки. Количественная характеристика генетического материала: 2n 2 хроматиды 4С.
Метафаза – хромосомы в животных клетках располагаются в упорядоченном состоянии в области экватора. Образуется метафазная пластинка. Завершается образование нитей веретена деления. Нити веретена, которые крепятся к кинетохорам, называются хромосомными, а которые идут от полюса к полюсу – полюсными. Количественная характеристика генетического материала: 2n 2 хроматиды 4С .
Анафаза – хромосомные нити веретена сокращаются. К противоположным полюсам расходятся хроматиды, которые принято называть дочерними хромосомами. На каждом полюсе количественная характеристика генетического материала: 2n 1хроматида 2С.
Телофаза – дочерние хромосомы, разошедшиеся к полюсам, деспирализуются, теряют ясные очертания, вокруг них формируются ядерные оболочки, восстанавливается ядрышко. Клеточный центр теряет активность. Начинается цитокинез – деление цитоплазмы. Итогом деления является образование двух диплоидных клеток.
Деление в растительной и животной клетках происходит сходно. Но в клетках высших растений отсутствует клеточный центр. Цитотомия в животных клетках происходит путем перетяжки (образование борозды), которая, углубляясь, делит клетку на две части. В клетках растений формируется в центре срединная пластинка, которая затем растет к периферии.
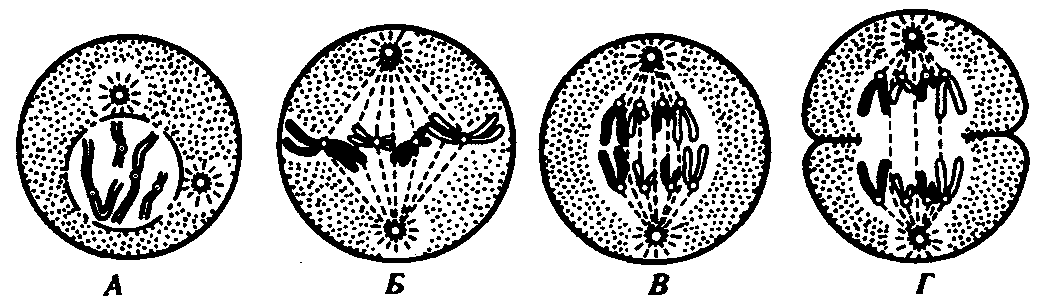
Рис. 10. Митоз в животной клетке.
А — профаза; Б — метафаза; В — анафаза; Г — телофаза
Мейоз I.
Профаза 1 имеет 5 стадий: лептотена, зиготена, пахитена, диплотена и диакинез. Количественная характеристика генетического материала: 2n 2 хроматиды 4С.
Лептотена (стадия тонких нитей) – хромосомы приобретают форму нитей, различимых в микроскоп. Хромосомы в это время состоят из двух хроматид, соединенных общим участком — центромерой. Это говорит о том, что удвоение хромосом, их редупликация, в основном произошла в интерфазе, предшествующей мейозу.
Последнее изменение этой страницы: 2020-02-16; Просмотров: 103; Нарушение авторского права страницы