|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
РЕГУЛЯЦИЯЭКСПРЕССИИ ГЕНОВ У ПРО- И ЭУКАРИОТ
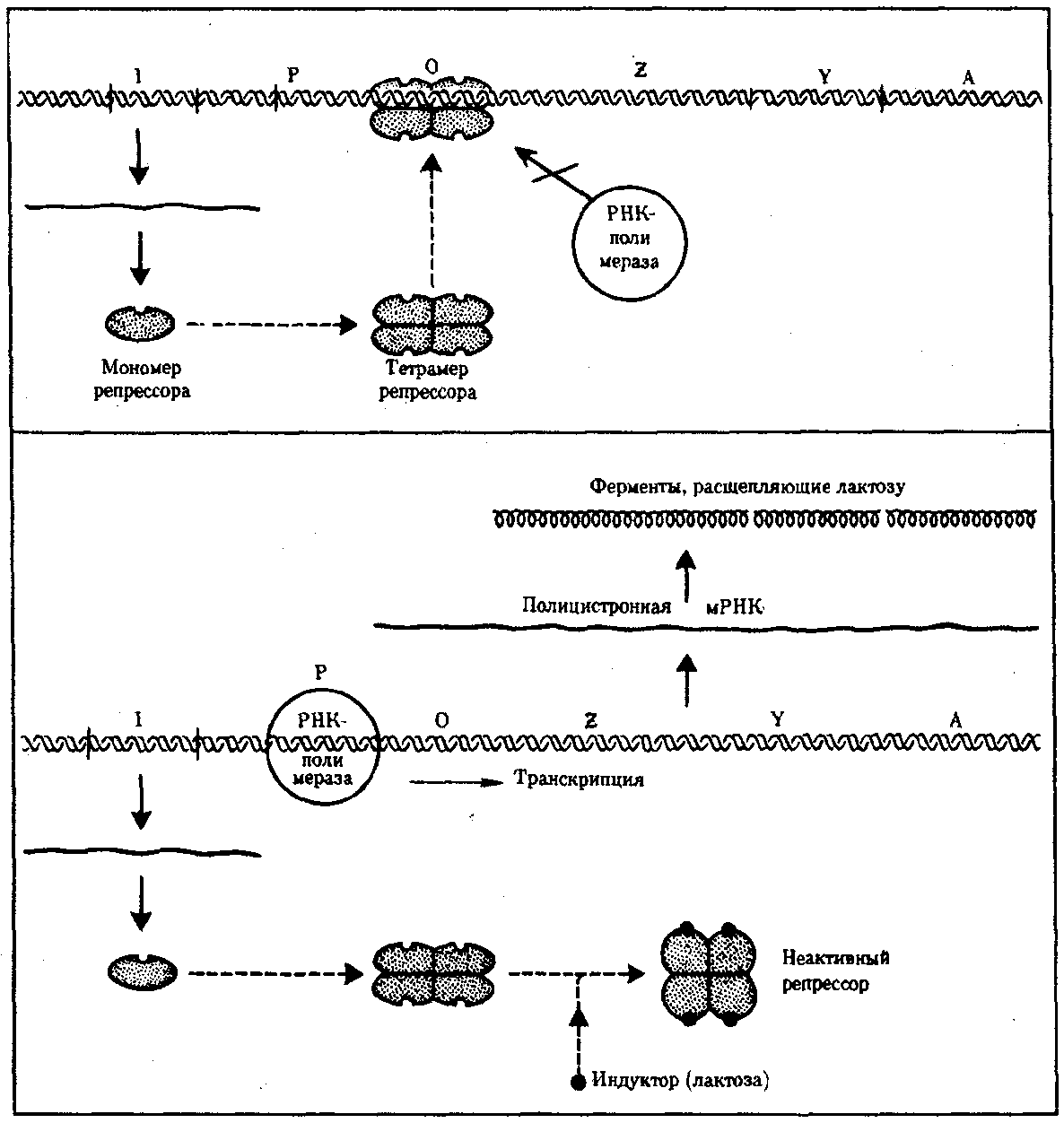
Генетические механизмы экспрессии генов были изучены у микроорганизмов французскими генетиками Ф. Жакобом и Ж. Моно. Один или несколько структурных генов, расположенных в бактериальной или вирусной «хромосоме» рядом с группой регуляторных генов, представляют вместе единицу генетической регуляции – оперон. Он включает структурные гены, несущие информацию о структуре белков, регуляторные гены, управляющие работой структурных. К регуляторным генам относят: промотор, оператор, терминатор. Промотор находится в начале каждого оперона. Это посадочная площадка для РНК‑ полимеразы (специфическая последовательность нуклеотидов ДНК, которую фермент узнаёт благодаря химическому сродству). Оператор управляет транскрипцией и имеет сродство с белком-регулятором. Терминатор включает стоп-кодоны, заканчивающие синтез и-РНК. Принципы работы оперона прокариот рассмотрим на примере работы оперона кишечной палочки (E. coli), на примере негативной регуляции (или негативного контроля) лактозного оперона у бактерии кишечной палочки E.coli (рис.26). При отсутствии в среде, на которой выращиваются бактерии, сахара лактозы активный белок-репрессор, синтезируемый геном-регулятором (I), взаимодействует с оператором (О), препятствуя соединению РНК-полимеразы с промотором (Р) и транскрипции структурных генов Z, Y, А. Появление в среде лактозы инактивирует репрессор, он не соединяется с оператором, РНК-полимераза взаимодействует с промотором и осуществляет транскрипцию полицистронной мРНК.
Рис. 26. Лактозный оперон Е. Со li
Последняя обеспечивает синтез сразу всех ферментов, участвующих в метаболизме лактозы. Уменьшение содержания лактозы в результате ее ферментативного расщепления приводит к восстановлению способности репрессора соединяться с оператором и прекращению транскрипции генов Z, Y, А. Таким образом, регуляция экспрессии генов, организованных у прокариот в опероны, является координированной. Синтез полицистронной мРНК обеспечивает одинаковый уровень синтеза всех ферментов, участвующих в биохимическом процессе. В связи с особенностями организации отдельных генов эукариот и генома в целом регуляция генной активности у них характеризуется некоторыми отличиями по сравнению с прокариотами. У эукариот не установлено оперонной организации генов. Гены, определяющие синтез ферментов одной цепи биохимических реакций, могут быть рассеяны в геноме и, очевидно, не имеют, как у прокариот, единой регулирующей системы (ген-регулятор, оператор, промотор). В связи с этим синтезируемые мРНК у эукариот моноцистронны, т.е. являются матрицами для отдельных пептидных цепей. В настоящее время механизмы регуляции и координации активности эукариотических генов интенсивно изучаются. Установлено, что их функционирование несомненно подчиняется регуляторным воздействиям, однако регуляция транскрипции у эукариот является комбинационной, т.е. активность каждого гена регулируется большим спектром генов-регуляторов (рис. 27).
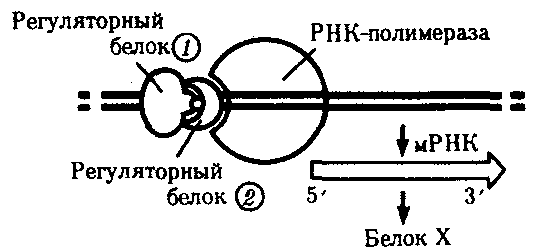
Рис. 27. Регуляция экспрессии гена, кодирующего белок Х у эукариот, двумя регуляторными белками
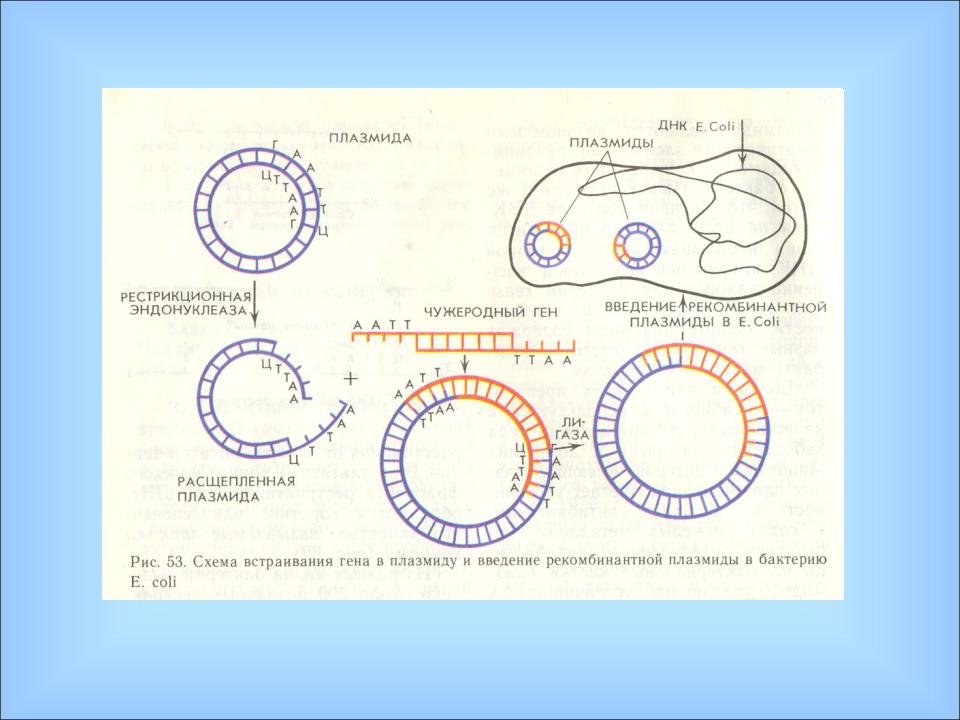
Как и у прокариот, регуляция транскрипции у эукариот опосредована ДНК - белковыми взаимодействиями. Белки позитивной и негативной регуляции связываются со специфическими областями ДНК и стимулируют, или ингибируют, транскрипцию. Группу таких белков называют транскрипционными факторами. Индукция генной экспрессии может быть обусловлена влиянием факторов внешней среды: температуры, света, а также действием факторов внутренней среды, таких как гормоны. У многоклеточных эукариот один тип клеток может подавать сигнал другому путем секреции гормонов. Гормоны циркулируют по телу. Разные гормоны проникают в разные клетки, вступая в контакт с соответствующими целевыми клетками и затем инициируют серию событий, которые регулируют экспрессию определенных тканеспецифических генов. В эмбриональный период развития особей гормоны являются одним из факторов дифференцировки клеток. Схематично генная экспрессия у эукариот может регулироваться на нескольких уровнях: 1. Регуляция уровня компактизации молекулы ДНК в виде эухроматина или гетерохроматина. На этом уровне работают ферменты, модифицирующие белковые молекулы гистонов, что позволяет осуществлять доступ ферментов транскрипции к определенным генам. В настоящее время этому уровню придается все большее значение. 2. Первичный контроль инициации транскрипции с участием регуляторных элементов ДНК и белковых факторов. 3. Регуляция формирования матричной РНК - процессинг транскрипта в зрелую мРНК. Различные формы мРНК обычно получаются из одногои того же гена путем альтернативного сплайсинга. 4. Контроль с участием особых молекул малых интерферирующихРНК (миРНК от англ. small interfering RNA или siРНК), способных комплементарно связываться с подавляемым геном или его транскриптом - интерференционной РНК. РНК-интерференция — это способ управления активностью генов посредством коротких двухцепочечных РНК, их назвали малые интерферирующие РНК (миРНК от англ. small interfering RNA или siРНК). При этом происходит выборочная деградация или ингибирование трансляции определенных мРНК, комплементарных миРНК. МиРНК являются короткими двуспиральными молекулами РНК, состоящими из 21-23 пары оснований. Попадая в клетку молекулы миРНК дают сигнал работе ряда ферментов, которые разрезают комплементарную одноцепочечную мРНК на отдельные фрагменты и удаляют из нее подлежащие ликвидации участки. Содержащаяся в этих участках информация не передается рибосомам. РНК– интерференция может разрушать генетический материал попадающих в клетку вирусов, уничтожает подвижные элементы его генома и участвует в регуляции экспрессии функциональных генов, блокируя их работу. Тем самым осуществляется особый вид внутриклеточного иммунитета с помощью молекул РНК. Также миРНК могут связываться с комплементарными участками ДНК и изменять структуру хроматина. 5. Контроль на уровне трансляции путем редактирования мРНК. 6. Контроль на посттрансляционном уровне. Трансляция – это механизм, с помощью которого последовательность нуклеотидов в молекулах м-РНК переводится в специфическую последовательность аминокислот в полипептидной цепи. Трансляция складывается из трех стадий: 1. Инициация – начало синтеза полипептида. 2. Элонгация – удлинение полипептида. 3. Терминация – окончание синтеза полипептида. На этапе инициации меньшая субъединица рибосомы узнает стартовый кодон АУГ м-РНК и прикрепляется к ней. АУГ занимает первую позицию. После этого присоединяется большая субъединица рибосомы и в ней начинается собственно синтез белка. К большой субъединице подходит т-РНК с аминокислотой. В молекуле т-РНК одна ее часть присоединяет аминокислоту, а другая – антикодон, спаривается с колоном м-РНК, определяющим эту аминокислоту. В т-РНК, антикодон которой комплементарен кодону АУГ м-РНК приносит аминокислоту метианин и останавливается в аминоацильном участке. В большой субъединице есть: 1 – аминоацильный участок 2 – пептидильный участок Стадия инициации контролируется факторами инициации. Вторая стадия – элонгации начинается с перемещения т-РНК с метианином в пептидильный участок, а на ее место приходит другая т -РНК с новой аминокислотой. Между двумя аминокислотами образуется пептидная связь. Тем временем рибосома продвигается вдоль м-РНК, на рибосоме оказывается новый кодон, к которому вскоре присоединяется своим антикодоном соответствующая т-РНК. Все это повторяется многократно, до тех пор, пока рибосома не дойдет до одного из терминальных кодонов (УАА, УАГ, УГА). Этим кодонам нет антикодона, нет аминокислоты. Наступает фаза терминации. К одной молекуле м-РНК прикрепляется обычно много рибосом и образуются полисомы. В процессе трансляции наряду с факторами инициации и элонгации важное значение имеет фермент, связывающий т-РНК с аминоацильным участком – аминоацил т-РНКаза. ГЕННАЯ ИНЖЕНЕРИЯ На основании достижений молекулярной биологии, биохимии и генетики в последние десятилетия интенсивно развивается новое направление в генетике - генная инженерия, целью которой является конструирование генетических структур по заранее намеченному плану, создание организмов с новой генетической программой путем переноса генетической информации из одного организма в другой. Методы генной инженерии были разработаны в 60-70-х годах XX века. Они включают следующие основные этапы: 1. получение генетического материала (выделение природных генов или химический их синтез): 2. включение этих генов в автономно реплицирующуюся генетическую структуру (векторную молекулу) и создание рекомбинантной ДНК; 3. введение рекомбинантных молекул ДНК в клетку-реципиент и включение ее в хромосомный аппарат; 4. отбор трансформированных клеток, в геном которых включен переносимый ген. В настоящее время применяют несколько способов получения генов для пересадки. Если полностью расшифрована последовательность нуклеотидов, то ген может быть синтезирован химическим путем. Впервые искусственный ген аланиновой т-РНК. состоящий из 77 нуклеотидов, был синтезирован индийским ученым Г. Корана (1970г.). В 1976 г. был синтезирован ген тирозиновой т-РНК. состоящий из структурной и регуляторной частей (промотор и терминатор), который при введении в бактериальную клетку нормально функционировал. Однако химическим способом удается синтезировать только небольшие по размеру гены прокариот. Синтез сложных генов осуществляют с помощью процессов обратной транскрипции, в основе которых лежит метод ферментативного синтеза. Выделяют и-РНК. и на ней как на матрице, с помощью фермента ревертазы (обратной транскриптазы) синтезируется комплементарная ей нить ДНК, а затем ее реплицируют (получают комплементарную цепочку). Гены, синтезированные с помощью ревертазы, не имеют регуляторной части и промотора и, вследствие этого, они не могут функционировать в животных клетках. При переносе в бактерию к структурным генам присоединяют промотор микробной клетки, после чего транскриптон начинает работать. Полученные различными способами гены соединяются с векторными молекулами, которыми чаще служат плазмиды бактерий. Кроме плазмид в качестве вектора используются фаги и вирусы. Они передают генетическую информацию посредством трансакции. Кольцевая молекула ДНК плазмиды разрывается той же рестриктазой. что и выделенный ген. В области разрыва образуются липкие концы, комплементарные липким концам пересаживаемого гена. Фермент лигаза сшивает липкие концы гена и плазмиды. Получается рекомбинантная молекула ДНК которая обладает способностью проникать в клетку-реципиент. Комбинируя различные рестриктазы и лигазы. можно разрезать нить ДНК в разных местах и получать рекомбинантные молекулы (Рис.28, 29)
Рис. 28. Схема действия рестриктаз
Так как не во все клетки попадут рекомбинированные молекулы ДНК, то с помощью специальных методов (чаше всего на селективных питательных средах) проводят отбор трансформированных клеток (с перенесенным геном). В дальнейшем проводят клонирование - размножение клеток с рекомбинантной ДНК - и получают клон клеток с заданными свойствами. Методами генной инженерии получены клоны клеток кишечной палочки, способные продуцировать соматотропин и инсулин в промышленных масштабах. Обычно эти препараты получают из соответствующих желез животных. Преимущества препаратов, полученных методами генной инженерии заключается в возможности синтеза их в достаточных количествах, биохимически чистыми и абсолютно стерильными. Генная инженерия - это современное интенсивно развивающееся направление генетики. С использованием ее методов созданы растения, способные усваивать атмосферный азот, микроорганизмы, разрушающие углеводороды нефти и синтезирующие из них пищевые белки. Разработаны методы внесения генов патогенных вирусов в бактерии и приготовление из синтезированных ими белков противовирусных сывороток. В будущем генная инженерия поможет человечеству избавиться от ряда наследственных заболевании, посредством пересадки в зародыш недостающих или замены мутантных генов. В настоящее время накапливаются клонированные гены человека, некоторых животных и растений, т.е. банки генов. Объединение чужеродных генов в одной клетке чревато опасными последствиями. Плазмиды способны соединяться в любых комбинациях, независимо от видовых и иммунологических барьеров. Конструирование новых разновидностей болезнетворных бактерий, устойчивых к лекарственным препаратам, может привести к возникновению серьезных эпидемий.
Рис. 29. Схема встраивания гена в плазмиду и введение рекомбинантной плазмиды в бактерию
В 1973 г. была проведена первая международная конференция по предупреждению опасных последствий генной инженерии. Опыты на время были запрещены. В 1975 г. Р. Кертис получил мутант кишечной палочки, нежизнеспособный в естественных условиях в связи с нарушением синтеза оболочки. Неопасная для человека и животных бактерия может жить только в лабораторных условиях, и опыты по генной инженерии были продолжены. Все исследования по генной инженерии проводятся в специальных лабораториях, строжайше изолированных от окружающей среды, с обязательным соблюдением определенных мер безопасности. Будущее генной инженерии базируется на следующих достижениях молекулярной биологии: 1) возможность с помощью химических мутагенов вызывать специфические мутации в определенных генных локусах: 2) возможность переноса генетической информации неполовым путем у эукариот (трансформация или трансакция), что позволит проводить генную терапию заболеваний: 3) замена дефектных генов с использованием ДНК вирусов в качестве переносчиков: 4) включение в геном человека искусственно синтезированных генов. 3. Перспективы генной терапии у человека. Необходимо различать 2 цели генной терапии - коррекцию генетических дефектов в соматических клетках и коррекцию их в гаметах или на самых ранних стадиях развития зиготы. В настоящее время единственными клетками человека, которые можно использовать для переноса генов, являются клетки костного мозга или фибробласты. Эти клетки можно извлечь из организма, вырастить в культуре, перенести в них нужный ген и снова ввести пациенту. Наиболее перспективным является перенос нужных генов, связанный с использованием ретровирусов. Чтобы применять на практике методы генной инженерии, нужно быть уверенным в их безопасности. Например, человеческие онкогены по структуре отчасти гомологичны ретровирусам и при заражении клеток такими вирусами возможна их модификация и превращение в онкогены. В экспериментах на мышах проведена генная терапия на уровне зиготы: в оплодотворенные яйцеклетки мышей карликовой линии вводили гены гормона роста крыс. При этом часть потомков (6 из 41) достигли гигантских размеров. Очевидно, что вновь встроенные гены не подвергаются нормальной регуляции, так как не удается внедрить их в места обычной локализации в хромосоме. Встраивание происходит в случайном порядке и в некоторых случаях это вызывало у мышей-реципиентов серьезные нарушения (мутации) работы нормальных генов в участках встраивания. По мнению большинства медицинских генетиков, метод генной терапии не следует в обозримом будущем применять к оплодотворенным клеткам человека, так как слишком велика опасность изменения генетической конституции человека.
ТЕМА: ИЗМЕНЧИВОСТЬ
|
Последнее изменение этой страницы: 2020-02-16; Просмотров: 79; Нарушение авторского права страницы