|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ФУНКЦИИ МОЗГА: МЕХАНИЗМ ДВУХ ПРОЦЕССОВСтр 1 из 16Следующая ⇒
«Язык, с помощью которого передается информация [в мозге] ... не соответствует и не должен соответствовать тому языку, которым люди пользуются в общении друг с другом». Питтс и Мак-Наллок
Глава I СОСТОЯНИЯ И ОПЕРАТОРЫ ВВЕДЕНИЕ «Я вас люблю». В Париже была весна, и эти слова были произнесены с прелестным скандинавским акцентом. Это событие произошло на заседании ЮНЕСКО, посвященном проблемам изучения мозга и человеческого поведения. Слова эти были произнесены не белокурой красавицей, а маленьким блестящим металлическим устройством, которое держал в руках известный психолингвист. Устройство поразило всех нас простотой своей конструкции. Громкоговоритель регулировался с помощью только двух ручек. Одна изменяла состояние электронной цепи, которое моделировало напряжение голосовых связок; другая регулировала импульсы, генерируемые цепью и имитировавшие взрывные звуки воздушной струи, ударявшейся по связкам. Может ли это простое устройство иметь отношение к изучению человеком самого себя? Нельзя ли вызывать и контролировать все поведение с помощью столь же простого нейронного механизма? Не является ли нервнйя система «двухкнопочным» механизмом двойного процесса, в котором один процесс выражается в терминах нейроэлектрических состояний, а другой — в терминах особых пульсирующих операторов, воздействующих на эти состояния? То, что нервная система фактически действует посредством импульсов, хорошо известно. Существование нейроэлектрических состояний мозга также было установлено, но доказательства их существования были получены не сразу, и понимание их значения для изучения поведения приходило лишь постепенно даже в нейрофизиологии. Поэтому в первой главе рассматриваются данные, свидетельствующие о правдоподобии двухпроцессной модели функций мозга.. Чтобы понять, в чем заключаются функции мозга, мы должны вначале понять, из каких единиц состоит нервная система. Классический анализ определяет эти единицы как нейроны — клетки, которые структурно и функционально изолированы друг от друга* мембранными оболочками. Однако такое упрощенное представление об изолированном нейроне как единственном организаторе мозговых процессов вызывает затруднение при попытке охарактеризовать нейроэлектрические состояния как часть такого двойного механизма. В двух первых разделах дается беглый обзор основ нейронной теории и тех причин, по которым в настоящее время в нее должны быть внесены некоторые поправки. В конце XIX в. в нейробиологии возникла полемика о том, состоит ли мозговая ткань из единиц — клеток, подобно всем другим тканям тела. Результаты этой полемики были настолько убедительны, что нейрон и его способность действовать как единица, разряжающаяся электрическим потенциалом по закону «все или ничего», перестали быть предметом теоретического рассмотрения. Еще никто не «видел» нейрон мозговой ткани; никто не проследил полностью его протяженность и не показал, что он действительно изолирован от других нейронов (рис. 1-1). Каким же образом нейронная теория получила в дальнейшем всеобщее признание и почему мы вновь вернулись к этой проблеме сегодня? Спор вокруг нейронной теории достиг своей кульминации в конце XIX в. Непрерывность нервной ткани признавали такие известные нейроанатомы, как Билыповский и Гольджи; Валытейер и Рамон-и-Кахал рассматривали нейрон как независимый, но соприкасающийся с другими нейронами. Однако убедительный анализ был дан не этими нейроанатомами, а сэром Чарлзом Шеррингтоном в его лекциях в Йельском университете, опубликованных как классическая работа «Интегративная деятельность нервной системы». Ранее Шеррингтон совместно с Фостером предложил термин «синапсы» для обозначения предполагаемых связей между отдельными нервными клетками. Он внес большой теоретический вклад в нейронную теорию, противопоставив нейрофизиологические данные, основанные главным образом на изучении электрических явлений в нервных стволах, в& щопсихологичеспим *, базирующимся на изучении рефлекторной деятельности у «спинального животного» (нашедшей отражение в концепции рефлекторной дуги, см. гл. V). Шеррингтон высказал мысль, что парадоксальные расхождения результатов нейрофизиологических и поведенческих исследований могут получить объяснение, если обратиться к нейроанатомическому описанию синапсов. Основные расхождения состояли в том, что (1) в изолированном стволе сигнал, вызванный стимуляцией какой-нибудь его точки, передается в двух направлениях, тогда как проведение сигнала при осуществлении спинального рефлекса происходит в одном направ- * Автор употребляет термин «neurobehavioral», который далее будет переводиться уже прочно вошедшим в литературу термином «нейропсихологический». — Прим. ред. Рис. 1-1. Сканирующая электронная микрофотография, показывающая расположение нервных волокон в сетчатке Necturus'a. Волокна (дендриты) берут начало от внутреннего членика колбочки и распространяются в сторону наружного. Следует отметить, что точки контакта необязательно находятся на нервных окончаниях (Lewis, 1970).
лении, и (2) скорость проведения импульса по нервному стволу значительно выше, чем скорость ответа, получаемого в результате рефлекса. Помимо этих, было отмечено девять других расхождений. Большинство из них касалось степени соответствия между адресованным организму стимулом и полученным ответом: соответствие всегда было большим для нервного, чем для поведенческого ответа. Чтобы объяснить эти расхождения, синапсам приписывались такие предполагаемые функции, как пространственная конвергенция и временная суммация, утомляемость, облегчение и торможение. Следующее утверждение Шеррингтона дает представление о его концепции: «Наиболее характерными особенностями проведения в рефлекторных дугах по сравнению с нервными стволами являются: 1) меньшая скорость, если судить по измерениям латентного периода между нанесением раздражения и появлением конечного эффекта, причем это различие больше для слабых раздражителей, чем для сильных; 2) менее близкое соответствие между моментом прекращения раздражения и моментом прекращения конечного эффекта (т. е. имеет место отчетливое «последействие»); 3) менее близкое соответствие между ритмом раздражения и ритмом конечного эффекта; 4) менее близкое соответствие между градуальным увеличением интенсивности раздражения и градуальным увеличением интенсивности конечного эффекта; 5) значительное сопротивление к прохождению нервного импульса, которое, однако, без труда преодолевается последовательностью импульсов (временная суммация); 6) необратимость направления проведения по сравнению с обратимостью в нервных стволах; 7) утомляемость в противоположность сравнительной неутомляемости нервных стволов; 8) более значительная изменчивость пороговых величин раздражения по сравнению с нервными стволами; 9) рефрактерный период, » проторение», торможение и способность впадать в состояние шока в степени, неизвестной для нервных стволов; 10) большая зависимость от кровообращения и снабжения кислородом (Ферворн, Винтерштейн, Бейер и др.); 11) более выраженная восприимчивость к действию различных лекарств-анестетиков» (Шеррингтон, 1969, стр. 40—41). Йороче говоря, результаты электрофизиологического исследования нервных стволов и поведенческого изучения рефлекса не совпадали. Нервы казались простыми проводящими путями для импульсов; рефлекс же обнаружил такую сложность организации, которая не могла быть объяснена простой «проволочной» моделью. Поэтому Шеррингтон принял «нейронную теорию», высказав предположение, что «провода», образующие нервную систему, составляют не непрерывную сеть, а слегка отделены друг от друга и включают структуру, которую он назвал синапсом. Затем он наделил синапс всей той сложностью, которую необходимо принимать во внимание при объяснении рефлекторного поведения. Таким образом, свойства изучаемого поведения были отнесены за счет свойств соединений между нейронами, а не их проводниковой части (рис. 1-2 и 1-3). Теоретическая гипотеза Шеррингтона в значительной степени игнорировалась исследователями в области нейрофизиологии и нейропсихологии. Внимание было сосредоточено на существова- нии и свойствах синапсов. Эти свойства детально изучались с помощью электронной микроскопии и записи электрической и Рис. 1-2. Скопление нейронов в сером веществе спинного мозга обезьяны, иллюстрирующее обычное расположение тел нервных клеток и их аксонных и дендритных отростков. Большая часть пространства между телами клеток занята нервным сплетением, состоящим из этих отростков и множества входящих и разветвляющихся аксонных окончаний. Сплетение аксонов и дендритов, посредством которого главным образом осуществляется связь одного нейрона с другим, известно под названием «нейропиль». р — перикарион; а — аксон; d — дендрит; пр — нейропиль; л — ядро нейрона большого размера, вероятно мотонейрона. Парафиновый срез, 1, 5 |х, окраска по Бодиану (Х150). (Видимое пространство вокруг тел нервных клеток — артефакт в результате сморщивания ткани). (Bodian, 1967.) химической активности соединений. Однако интерпретация этих результатов почти не выходила за рамки вопроса: «Как осуществляется прохождение нервных импульсов через барьер синапса? » Ссылка на разряды, генерируемые клеткой, и на проводящие-
•двойства синаптических путей слишком часто рассматривалась как самоочевидная и достаточная. Немногие ученые придерживались тезиса Шеррингтона о том, что сложность поведения (и психологических процессов) следует объяснять сложностью организации соединительных (синаптических) механизмов в центральной нервной системе.
Рис. 1-3. Схема микроструктуры синаптических связей в коре. Частично перекрывающие друг друга окружности изображают область соединительных контактов между разветвлениями входных аксонов и кортикальных дендритов (Scheibel and Scheibel, см. Chow and Leiman, 1970). Цель данной книги состоит в том, чтобы восполнить этот пробел. В ней сделана попытка, исходя из идей Шеррингтона, решить проблему взаимоотношений между мозгом, сознанием и поведением, подобно тому как Шеррингтон решал проблему отношений между спинным мозгом и рефлексом. Проблемы, возникающие при изучении головного мозга, тоньше и сложнее, чем проблемы, связанные со спинным мозгом. Тем не менее можно уже теперь достигнуть значительных успехов в понимании этих проблем при осторожном использовании классического нейробиологического подхода Шеррингтона. Однако неврологи, физиологи и психологи часто его игнорируют, переходя от отчаяния к неопределенным надеждам, а порой даже вообще отрицают существование такой проблемы. Изучение отношений между мозгом, сознанием и поведение» может быть продуктивным только в том случае, если ставятся ограниченные проблемы. Каковы же эти проблемы? НЕКОТОРЫЕ НЕЙРОПСИХОЛОГИЧЕСКИЕ ФАКТЫ Хотя нейрофизиологический эксперимент основывался на нейронной теории, некоторые известные психологи неоднократно отмечали, что понимание нервной системы как функционирующей исключительно в виде набора дискретно проводящих единиц не согласуется с полученными ими экспериментальными данными. Эти исследователи объясняли свои наблюдения, прибегая скорее к той или иной модели поля, чем к квантовой, дискретной, вероятностной нейронной теории. Разногласия, коротко говоря, сводились к следующему: «Физиолог слепо верит в то, что, поскольку мозг состоит из нейронов, он способен только к такому возбуждению, которое является суммой возбуждений многих нейронов, и что эти центральные нейроны подчиняются тем же законам и возбуждаются при тех же условиях, что и периферические нейроны, которые были изучены экспериментально. Этому утверждению физиолога психолог иногда противопоставляет другое мнение, а именно, что организация центрального возбуждения протекает иначе, что оно определяется рамками получаемого индивидом опыта» (Boring, 1932, р. 32). Проблема возникает особенно тогда, когда проводятся нейропсихологические эксперименты, предполагающие разрушение мозга. Результаты этих экспериментов послужили основанием для защиты двух противоположных точек зрения. По мнению одних авторов, каждая кортикальная точка, каждая клетка или группа клеток специализируется на выполнении какой-нибудь одной функции. Интеграция, необходимая для объяснения поведенческих и психологических процессов, согласно этой точке зрения, достигается за счет врожденных или образующихся при жизни прочных ассоциативных связей между нейронами. Другая точка зрения, которой придерживается и автор этой книги, состоит в том, что в мозговой ткани возникают определенные важные для организации поведения и психологического процесса взаимодействия и эти взаимодействия нельзя рассматривать только как постоянные ассоциативные связи между нейронами. К. Лешли, первый сторонник этой точки зрения, построил свою аргументацию по трем линиям: 1) эквивалентность рецепторной, функции; 2) спонтанная реорганизация моторных реакций и 3) сохранение поведения после разрушения любой части мозга, в то время как лишь разрушение всей мозговой системы устраняет раз возникшее поведение. Остановимся на этих трех ли-4 пнях доказательства. Что касается эквивалентности рецепторной функции, то эксперименты показывают, что «абсолютные характеристики стимула ямеют сравнительно небольшое значение для поведения и что реакция определяется отношениями возбуждений, которые при возникновении в любой группе рецепторных клеток этой системы имеют одинаковый эффект». Например, «животное, тренированное в выборе большего из трех кругов, может сразу же положительно реагировать на поле с наиболее широкими линиями при сравнении трех полей с полосами различной ширины» (Lashley, 1960, 238—239). Подробные данные такого рода содержатся во второй части книги; сейчас же достаточно отметить, что такая проблема существует. Что касается спонтанной моторной реорганизации, то «результаты показывают, что, когда привычно используемые двигательные органы перестают функционировать вследствие их удаления или паралича, наблюдается непосредственное спонтанное использование других моторных систем, которые ранее не были связаны •с этой деятельностью или не использовались при ее осуществлении» (Lashley, 1960, р. 239). Например, основной рисунок мышечного напряжения во время.акта письма, когда человек держит карандаш в пальцах, сохраняется и тогда, когда он переходит к письму, зажав карандаш зубами. Каким образом мозг обеспечивает это, составляет содержание первых глав третьей части этой книги. Доказательство взаимозаменяемости частей функциональных систем организма в организации поведения и психологических процессов распространяется также и на центральные отделы мозга. Это доказательство содержится во второй части. Здесь же лам необходимо только отметить, что даже значительное повреждение мозга может оказать лишь небольшое влияние на то крайне сложное поведение, которое, казалось бы, должно быть особенно чувствительно к разрушению, если считать, что интеграция целиком зависит от наличия постоянных ассоциативных связей. Лешли (см. Beach и др., 1960) следующим образом резюмирует проблему: «Эти три линии доказательства показывают, что некоторые хорошо координированные виды деятельности, казалось бы зависящие от определенных областей коры, могут в известных пределах осуществляться любой частью коры. Это может происходить в результате наличия многих дублирующих рефлекторных путей, проходящих через кору, и такое объяснение, по-видимому, можно дать всем известным случаям сохранения функций после частичного разрушения определенных областей, имеющих отношение к этим функциям. Но оно не годится для объяснения фактов сенсорной и моторной эквивалентности. Эти факты говорят о том, что, если была выработана условная реакция (например, положительная реакция на определенную структуру зрительного сигнала), эта реакция будет вызываться возбуждением сенсорных клеток, которые раньше никогда не стимулировались таким способом. Точно так же однажды приобретенные двигательные акты (например, открывание задвижки ящика) могут быть сразу же выполнены двигательными органами, которые не включались ранее в выполнение этях актов» (р. 237—240) В утверждении Лешли сметаны по крайней мере два вопроса: взаимозаменяемость частей организма и перенос навыка, который? включает в себя проблему сходства и новизны. Тем не менее приведенные здесь высказывания Лешли говорят о необходимости согласования данных психологии с данными нейрофизиологии. Такое согласование предполагает более широкое использование фактов, касающихся работы мозга, чем то, которое возможна сейчас вследствие узкого толкования нейронной теории. Между тем окончательная точка зрения на функции мозга должна быть основана на классических нейрофизиологических данных. Поэтому остановимся на тех последних успехах нейрофизиологической техники, которые показывают, что такое согласование действительно возможно (Bullock, 1959). Этот путь позволяет провести различие между первичными структурами интранейронных нервных импульсов и первичными интернейронными структурами активности; такой путь значительно расширяет диапазон тех подходов, которые применяются для изучения функций мозга. МИКРОСТРУКТУРА СОЕДИНЕНИЙ Значение нейроэлектрических процессов, генерируемых в синапсе и позади него, стало очевидным для нейрофизиологов после того, как были получены определенные данные. В течение многих лет электрическая активность, отводимая от кожи черепа, рассматривалась как отражение совокупности нервных импульсов, генерируемых нейронами мозга, лежащими где-то ниже участка записи. Когда стала возможной одновременная регистрация электрической активности локальных участков мозговой ткани и их совокупности, это предположение стало вызывать сомнение (Purpura, 1958). Запись электрической активности нейрона при внутриклеточном отведении показывает, что даже при отсутствии широко распространяющихся нервных импульсов возникают ритмические изменения медленных потенциалов, и, когда их сравнивают с ритмами, одновременно записанными от группы нейронов той же самой точки, они совпадают (Creutzfeldt et al., 1966; Fugita and Sato, 1964; Elul and Adey, 1966; Morrell, 1967). Когда мембрана нервной клетки подвергается поляризации постоянным током, амплитуда внутриклеточных ритмов меняется и это изменение отражается в суммарной записи (Eccles, 1964, см. рис. 1-4). Таким образом, было получено серьезное доказательство того, что-главный вклад в электрическую активность, записываемую суммарно в виде электроэнцефалограммы (ЭЭГ), вносится медленно возникающими флуктуациями потенциалов нейронных мембран (рис. 1-5). Только тогда, когда активность большого числа единиц синхронизирована — когда их разряды вызываются вспышкой
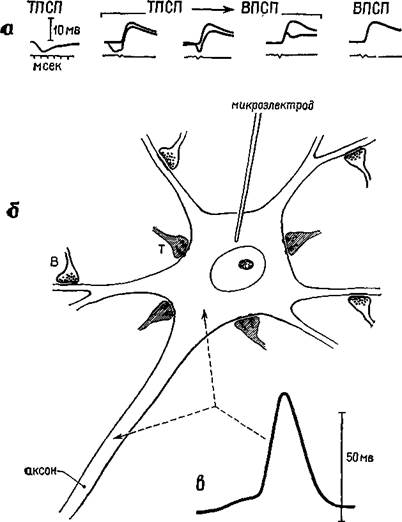
Рис. 1-4. Схема нейрона (б) с возбудительными (Б) и тормозными (Т) •синапсами и внутриклеточные записи (а) медленных постсинаптических потенциалов (ВПСП и ТПСП) — вверху. Следует отметить, что импульс, генерируемый на начальной части аксона, имеет амплитуду выше 50 мВ, тогда как амплитуда внутриклеточных записей имеет диапазон около 5 мВ. Только нервному импульсу аксона свойственно распространение (см. Eccles, 1967).
Рис. 1-5. Записи медленных дендритных потенциалов (справа) с указанием примерного уровня их регистрации в коре. Калибровка 0, 2 мВ; 10 мсек (Purpura, 1967). света или звуковым шелчком, — суммарная запись и импульсы, генерируемые отдельными нейронами, оказываются одинаковыми (рис. 1-7 и 1-8). В одном таком исследовании (Fox and O'Brien,. 1965) животному предъявлялось много световых вспышек (от 3000 до 5000). Когда разряды нейрона были усреднены для всего
Рис 1-6. Диаграмма связи между внутриклеточными записями медленных потенциалов и ЭЭГ, записанной с поверхности коры той же самой области мозга, для 500-секундного периода времени (Adey, 1967).
Рис 1-7 и 1-8. Отношения между синхронизацией волн ЭЗГ, нейронными разрядами и циркуляцией нейронной активности при одновременной записи четырьмя микроэлектродами, удаленными друг от друга на 100— 150 м- Внизу: двухмерная схема нейронных полей, показывающая нейронную активность каждой последовательной стадии (Verzeano et al., 1970). Рис. 1-9. Основные типы нейронов в центральной нервной системе млекопитающих в соответствии с выполняемой ими функцией, иерархическим уровнем и вероятной степенью разветвленности их отростков (Bodian, 1967). числа предъявлении, вероятность разряда одиночной клетки зрительной коры в период, следовавший за вспышкой (в течение 1 сек), приблизилась к волновой форме симультанно записанного суммарного потенциала. Таким образом, суммарный потенциал указывает на вероятность, с которой будет разряжаться нейрон после предъявления кратковременного стимула, способного синхронизировать группы нейронов. Однако при отсутствии столь отчетливо выраженной синхронизации суммарная запись отражает электрические изменения потенциала, возникающие в сплетении нервных волокон, которые представляют собой нервную ткань, но фактически не слособны вызывать спаиковые разряды нейронов (Li, Cullen, and Jasper, 1956; Creutzfeldt, 1961; Verzeano and Negishi, 1960; см. рис. 1-5 и 1-6). Эти экспериментальные данные заставляют фактически различать два типа нейроэлектрической активности: импульсные разряды нейрона, с одной стороны, и градуальные изменения медленного потенциала — с другой. Распространяются только нервные импульсы; градуальные изменения усиливаются и ослабевают в мозговой ткани локально, они чувствительны к различным
влияниям, таким, как локальная химическая среда, которая по своей природе не является строго нейронной. Различие между градуальными медленными потенциалами и нервными импульсами зависит частично от того, что амплитуда и скорость проведения нервного импульса пропорциональны диаметру нервного ствола, в котором возникает импульс. Таким образом, в нервных волокнах большого диаметра импульсы имеют, как правило, значительную амплитуду и быстро распространяются. В нервных волокнах с малым диаметром амплитуда электрического потенциала нервного импульса меньше (по крайней мере при экстраклеточном отведении) и скорость его проведения ниже. По мере удаления нервных волокон от тела клетки они утончаются и разветвляются; в результате этого импульсы становятся все более слабыми и замедленными, особенно на окончаниях волокон (рис. 1-9). Поэтому необходимы химические медиаторы, которые усиливали бы изменение потенциала, возникающее в нервных окончаниях, чтобы электрический сигнал мог распространить свое действие через структурные соединения между нейронами. Несмотря на химический усилитель, потенциалы, генерируемые позади синапса (названные постсипаптическими потенциалами), первоначально возникают как рудиментарные медленные градуальные изменения, подверженные влиянию химического посредника, создаваемого текущим по сосудам током крови и ненейронными (глиальными) клетками, окружающими волокна. Кроме того, более тонкие нервные волокна теряют толстое изолирующее покрытие (миелиновую оболочку), которая у крупных волокон препятствует взаимодействию импульсов. Медленные потенциалы, возникающие в таких тонких волокнах, как разветвления дендритов, следовательно, не встречают препятствий для реального локального взаимодействия., Упрощенная до некоторой степени картина (рис. 1-10) взаимоотношений между изменениями медленных потенциалов и нервными импульсами выглядит следующим образом: внутри нейрона электрохимические процессы ведут к деполяризации мембраны — возрастанию градуальных потенциалов, пока они не становятся нервными импульсами. Там, где соединяются нейроны, или где тонкие, немиелинизированные волокна формируют сплетение и образуют контакты относительно бесструктурным способом, названным эфаптическим (рис. 1-11), или где окончания волокон действительно образуют структурный контакт с другими нейронами посредством синапса (рис. 1-12), нервные импульсы постепенно затухают, то есть превращаются в небольшие медленные потенциалы, не отличимые от спонтанно генерированной локальной градуальной активности (см. гл. III). Короче говоря, эффективный иот/хшейронный механизм характеризуется тем, что возрастание градуальной активности приводит к появлению распространяющихся нервных импульсов. Рис. 1-10. Схема, показывающая отношения между градуальными медленными потенциалами, нервными импульсами и химической секрецией, которая действует через структурированные соединения между нейронами (синапсы) как химический возбудитель (Grundfest, 1967). Эффективный механизм соединений характеризуется противоположными чертами: нервные импульсы затухают, трансформируясь в локальные медленные потенциалы. Соединительный механизм в свою очередь влияет на интранейронную мембранную деполяризацию, но только после того, как реализуется возможность пространственных и временных взаимодействий между конфигурациями медленных потенциалов. Именно в этих взаимодействиях между конфигурациями и состоит специфическое значение медленных потенциалов. Такая точка зрения на активность соединений несколько отличается от широко распространенного понимания синаптической функции. В большинстве нейрофизиологических исследований синапса ставился вопрос: «Каким образом нервные импульсы или передаваемая ими информация преодолевают синаптическую щель? » Настоящий подход обращает внимание на медленные потенциалы, как таковые; в основе его лежит предположение, что медленные потенциалы образуют динамические структуры, которые выполняют какую-то функцию, помимо той роли, какую они играют в передаче импульсов. Согласно принятой здесь точке зрения, структуры медленных потенциалов «вычисляют» взаимодействие между соседними нейронами в пространстве и в какой-то
Рис. I—Id. Электрофизиологическое доказательство (представленное внутриклеточными записями двух нейронов) эфаптической передачи через неструктурированные контакты между нейронами, выявленные с помощью электронной микроскопии. Залпы электрических импульсов (В, В) и переменный ток (Г) воздействовали на одну клетку, а запись активности производилась от другой. Слабая стимуляция одной клетки (А) вдали от другой вызывала импульсы почта одновременно в обеих клетках (Grundiest, 1967).
Рис. 1-12. Синаптические контакты на пересечении двух нервных do-локон в нейропиле брюшного ганглия Aplesya Californica. Увеличение примерно в 35 000 (Lewis, 1970). мере их взаимодействие во времени скорее с помощью непрерывного (аналогового), чем дискретного (цифрового) механизма, работающего по принципу «все или ничего». Совокупность медленных потенциалов, в любой момент присутствующая на обширном участке, может быть описана как состояние, имеющее некоторую микроструктуру. Приход импульсов на эфаптические или синаптические соединения никогда не бывает одиночным. Аксонные окончания обычно многочисленны, ибо аксоны ветвятся на своих концах. Соединение между двумя нейронами может осуществляться с помощью большого количества синапсов, доходящего до 1000. Дендриты, уже по определению напоминающие деревья, обладают многочисленными тонкими волокнистыми ветвями, которые, пересекаясь, образуют множественные контакты между нейронами. Эти контакты у некоторых клеток (например, амакриновых клеток сетчатки) включают структурные синапсы (рис. 1-13). Таким образом, эфаптические и синаптические процессы, возникающие в местах соединений между нейронами, формируют определенную динамическую структуру, заключение о природе которой может быть сделано на основе зна-
Рис. 1-13. Зарисовка, показывающая многочисленные точки контакта (синаптического и эфаптического) между двумя нейронами мозга (Ralston, 1968). пия тонкой структуры мозга и его электрической активности. Можно, в частности, предположить, что эти динамические структуры создают волновые фронты. Это предположение становится особенно полезным при рассмотрении проблем эквивалентности (см. вторую часть книги). Это не значит, что микроструктуру медленного потенциала следует рассматривать в терминах механической волны, но иногда это сходство полезно иметь в виду. Ведь именно интерпретация динамических структур медленных потенциалов как образующих волновые фронты помогает представить себе то, что происходит. «Нейрон — это реле, действующее по закону «все или ничего». Импульс, достигающий синапса, вызывает очень небольшой и временный электрический эффект равный 0, 001 вольта и длящийся от 0, 01 до 0, 02 сек. Необходимо примерно в 10 раз большее возбуждение, чтобы вызвать разряд Нейрона. Так как требуется конвергенция многих импульсов на какой-нибудь «дин нейрон, чтобы вызвать его разряд, то цепи, состоящие из одиночных нейронов, не могут обеспечить распространение волны активности в коре.
Рис. 1-14. На этой диаграмме синаптические области коры изображены в виде кружков на плоскости. Движение процесса, которое может осуществляться во многих направлениях и характеризуется специфической нейронной структурой возбуждения, показано черными кружками для одного направления и темно-серыми для другого. Области с контурными кружками не активизируются ни одной из динамических структур. Следует отметить, что на пересечении этих двух направлений движения одни и те же области принимали бы участие в обоих процессах и поэтому каждая из них была бы изображена в виде очень большого серого кружка, а не как темно-серый или черный кружок, как показано на рисунке (Eccles, 1970). Распространение активности представляет собой, скорее, передний фронт движения во многих направлениях, когда параллельно активизированы многие клетки в каждом синаптическом звене цепи... (рис. 1-14). Мы сразу же получаем объяснение одного замечательного свойства нервной сети: каким образом два совершенно различных входных воздействия (одно на клетки А, и А2, другое — на клетки А3 и А4) могут быть переданы посредством одной и той же динамической структуры клеточных связей, противодействуя друг другу и достигая совершенно различных выходов (Г3 — Г4)... (рис. 1-15). Конечно, передача волнового фронта в коре происходит гораздо сложнее. При вовлечении 100 нейронов на каждой релейной станции передняя волна может захватывать более 100 000 нейронов в одну секунду. Такая волна обладает... богатыми потенциальными возможностями...» (Eccles, 4958, р. 4-7). Другой вывод следует из модели не распространяющейся стоячей волны, основанной на идеализированном представлении об анатомическом распределении связей между нейронами (Scholl, 1956). Все предположения, лежащие в основе этой модели, вполне
Рис. 1-15. Модель очень схематизированной нейронной сети, иллюстрирующая простейший случай проведения в системе с множеством направлений. На верхней и нижней диаграммах представлены нервные чзети с одной и той же анатомией. В колонках А, Б и В изображены синаптические контакты 12 клеток, клетки генерирующие импульс (со стрелками) — светло-серого цвета, молчащие клетки — черные. Предполагается, что клетка разряжается импульсом, если возбуждается двумя или больше синапсами (также светло-серого цвета). Таким образом, входное воздействие А]А2 вызывает разряд Г3Г4 (верхняя диаграмма), в то время как вход от А3А4 достигает выхода на Г1Г2 (нижняя диаграмма). Нейроны Бг, Б3, Вг, В3 активизируются в зоне пересечения этих двух входных воздействий. Серьезным недостатком этой диаграммы является то, что она игнорирует тормозные элементы (Eccles, 1968). разумны; 1) нейроны распределены в случайном порядке; 2) богатство связей между клетками уменьшается с расстоянием;. 3) пространственная структура распределения процессов в каждой клетке отличается тем, каким образом она передает возбуждение другим клеткам; 4) наблюдается затухание возбуждения во времени, то есть изменение потенциала в местах соединений имеет конечную длительность, и, наконец, 5) возбуждение само себя поддерживает. Последнее предположение при отсутствии информации о тормозных взаимодействиях говорит о том, что любое состояние равновесия будет неустойчивым и что «вскоре обнаружится, что-активность в одних местах уже прекратилась, а в других возникла. По этой причине всегда предпочтительнее активность,, имеющая определенную пространственную и временную организацию...» (Beurle, 1956). Детальные исследования электрической, а также анатомической структуры некоторых наиболее организованных мозговых образований в целом подтверждают обоснованность этих заключений. «Лучше всего вто можно понять, если представить себе, что кора мозжечка... непрерывно подвергается воздействиям вследствие возникновения микроволн; каждая волна — это небольшой гребень активации длиной 3 мм, она возникает от клетки Пуркинье, которая имеет на другом конце тормозной сток. Эти волны не распространяются, но, конкурируя и интерферируя друг с другом, они, конечно, сильно модифицируют структуру волновых форм; более того, такая интерференция ведет даже к укорочению-волны, длительность которой становится менее 100 мсек. Эта операция конкурирующего взаимодействия волновых структур должна быть ключом к разгадке действия нейронного механизма...» (Eccles, Ito and SzentagoLhai,. 1967, p. 342). Действительно, запись электрической активности, полученная от множества электродов, которая одновременно может быть представлена в пространстве (на топоскопе), говорит о возникновении какой-то волновой активности (Walter and Shiptom, 1951;. Lilly, 1949; Ливанов и Ананьев, 1955; Rernond, 1961). В каком отношении эта волновая форма соответствует нераспространяющейся стоячей волне, о которой говорил Берл (Beurle), остается еще исследовать. Важно в данном случае то, что описание нейронной активности медленных потенциалов как механической волны может помочь представить себе природу микроструктуры медленного потенциала или — более формально — послужить моделью, которая поможет дать более точную интерпретацию данных. Таким образом, подобный подход, состоящий в уподоблении медленного потенциала механической волне, хотя и не является необходимым для описания микроструктуры медленного потенциала, может в данном случае оказаться полезным (см. гл. VIII, рис. 1-16). В заключение скажем, что наиболее распространенные представления о функции мозга основаны на том, что нервные им- пульсы возникают в нейронах и по ним передаются. Хотя точные нейрофизиологические исследования иногда предостерегают против чрезмерного упрощения, эти привычные представления, как ни странно, игнорируют активность соединений за исключением тех случаев, когда она имеет отношение к передаче нервных импульсов. Согласно этим взглядам, первичная задача синапса (или дендрита) состоит в передаче (или генерации) импульсов.
Рис. 1-16. Диаграмма, показывающая зависимость ЭЭГ космонавта Ф. В. от высоты подъема в камере, имитировавшей полет «Джеминай». Соотношение было исследовано на протяжении 70-минутного периода времени, в течение которого было сделано 40 записей. Каждая из них длительностью 20 мин. Обратите внимание на характеристику волнового фронта диаграммы и сравните ее с диаграммой на рис. 1-6 (Adey, 1967). Популярное:
|
Последнее изменение этой страницы: 2016-03-17; Просмотров: 565; Нарушение авторского права страницы