|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Онтогенез — основа филогенеза
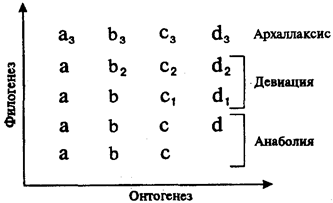
Опираясь только на основной биогенетический закон, невозможно объяснить процесс эволюции: бесконечное повторение пройденного само по себе не рождает нового. Так как жизнь существует на Земле благодаря смене поколений конкретных организмов, эволюция ее протекает благодаря изменениям, происходящим в их онтогенезах. Эти изменения сводятся к тому, что конкретные онтогенезы отклоняются от пути, проложенного предковыми формами, и приобретают новые черты. К таким отклонениям относятся, например, ценогенезы — приспособления, возникающие у зародышей или личинок и адаптирующие их к особенностям среды обитания. У взрослых организмов ценогенезы не сохраняются. Примерами ценогенезов являются роговые образования во рту личинок бесхвостых земноводных, облегчающие им питание растительной пищей. В процессе метаморфоза у лягушонка они исчезают и пищеварительная система перестраивается для питания насекомыми и червями. К ценогенезам у амниот относят зародышевые оболочки, желточный мешок и аллантоис, а у плацентарных млекопитающих и человека — еще и плаценту с пуповиной. Ценогенезы, проявляясь только на ранних стадиях онтогенеза, не изменяют типа организации взрослого организма, но обеспечивают более высокую вероятность выживания потомства. Они могут сопровождаться при этом уменьшением плодовитости и удлинением зародышевого или личиночного периода, благодаря чему организм в постэмбриональном или постличиночном периоде развития оказывается более зрелым и активным. Возникнув и оказавшись полезными, ценогенезы будут воспроизводиться в последующих поколениях. Так, амнион, появившийся впервые у предков пресмыкающихся в каменноугольном периоде палеозойской эры, воспроизводится у всех позвоночных, развивающихся на суше, как у яйцекладущих — пресмыкающихся и птиц, так и у плацентарных млекопитающих. Другой тип филогенетически значимых преобразований филогенеза — филэмбриогенезы. Они представляют собой отклонения от онтогенеза, характерного для предков, проявляющиеся в эмбриогенезе, но имеющие адаптивное значение у взрослых форм. Так, закладки волосяного покрова появляются у млекопитающих на очень ранних стадиях эмбрионального развития, но сам волосяной покров имеет значение только у взрослых организмов. Такие изменения онтогенеза, будучи полезными, закрепляются естественным отбором и воспроизводятся в последующих поколениях. В основе этих изменений лежат те же механизмы, которые обусловливают врожденные пороки развития: нарушение пролиферации клеток, их перемещения, адгезии, гибели или дифференцировки (см. § 8.2 и 9.3). Однако от пороков их так же, как и ценогенезы, отличает адаптивная ценность, т.е. полезность и закрепленность естественным отбором в филогенезе. В зависимости от того, на каких этапах эмбриогенеза и морфогенеза конкретных структур возникают изменения развития, имеющие значение филэмбриогенезов, различают три их типа. 1. Анаболии, или надставки, возникают после того, как орган практически завершил свое развитие, и выражаются в добавлении дополнительных стадий, изменяющих конечный результат. К анаболиям относят такие явления, как приобретение специфической формы тела камбалой лишь после того, как из икринки вылупляется малек, неотличимый от других рыб, а также появление изгибов позвоночника, сращение швов в мозговом черепе, окончательное перераспределение кровеносных сосудов в организме млекопитающих и человека. 2. Девиации — уклонения, возникающие в процессе морфогенеза органа. Примером может являться развитие сердца в онтогенезе млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и трехкамерное строение, но стадия формирования неполной перегородки, характерной для пресмыкающихся, вытесняется развитием перегородки, построенной и расположенной иначе и характерной только для млекопитающих (см. § 14.4). В развитии легких у млекопитающих также обнаруживается рекапитуляция ранних стадий предков, позднее морфогенез идет по-новому (см. разд. 14.3.4).
Рис. 13.9. Преобразования онто- и филогенеза в связи с возникающими филэмбриогенезами
Буквами обозначены этапы онтогенеза, цифрами — филэмбриогенетические преобразования
3. Архаллаксисы — изменения, обнаруживающиеся на уровне зачатков и выражающиеся в нарушении их расчленения, ранних дифференцировок или в появлении принципиально новых закладок. Классическим примером архаллаксиса является развитие волос у млекопитающих, закладка которых наступает на очень ранних стадиях развития и с самого начала отличается от закладок других придатков кожи позвоночных (см. § 14.1). По типу архаллаксиса возникают хорда у примитивных бесчерепных, хрящевой позвоночник у хрящевых рыб (см. разд. 14.2.1.1), развиваются нефроны вторичной почки у пресмыкающихся (см. разд. 14.5.1). Ясно, что при эволюции за счет анаболии в онтогенезах потомков полностью реализуется основной биогенетический закон, т.е. происходят рекапитуляции всех предковых стадий развития. При девиациях ранние предковые стадии рекапитулируют, а более поздние заменяются развитием в новом направлении. Архаллаксисы полностью не допускают рекапитуляции в развитии данных структур, изменяя сами их зачатки. Если сопоставить схему филэмбриогенезов с таблицей К. Бэра (рис. 13.9), иллюстрирующей закон зародышевого сходства, то станет понятно, что Бэр уже был очень близок к открытию филэмбриогенезов, но отсутствие эволюционной идеи в его рассуждениях не позволило более чем на 100 лет опередить научную мысль. В эволюции онтогенеза наиболее часто встречаются анаболии как филэмбриогенезы, лишь в малой степени изменяющие целостный процесс развития. Девиации как нарушения морфогенетического процесса в эмбриогенезе часто отметаются естественным отбором и встречаются поэтому значительно реже. Наиболее редко в эволюции проявляются архаллаксисы в связи с тем, что они изменяют весь ход эмбриогенеза, и если такие изменения затрагивают зачатки жизненно важных органов или органов, имеющих значение эмбриональных организационных центров (см. разд. 8.2.6), то часто они оказываются несовместимыми с жизнью. В одной и той же филогенетической группе эволюция в разных системах органов может происходить за счет разных филэмбриогенезов. Так, в онтогенезе млекопитающих прослеживаются все этапы развития осевого скелета в подтипе позвоночных (анаболии), в развитии сердца рекапитулируют лишь ранние стадии (девиация), а в развитии придатков кожи рекапитуляции вообще отсутствуют (архаллаксис). Знание типов филэмбриогенезов в эволюции систем органов хордовых необходимо врачу для прогнозирования возможности возникновения у плодов и новорожденных врожденных пороков развития атавистической природы (см. разд. 13.3.4). Действительно, если в системе органов, эволюционирующей путем анаболии и девиаций, возможны атавистические пороки развития за счет рекапитуляции предковых состояний, то в случае архаллаксисов это исключается полностью. Кроме ценогенезов и филэмбриогенезов в эволюции онтогенеза могут обнаруживаться еще и отклонения времени закладки органов — гетерохронии — и места их развития — гетеротопии. Как первые, так и вторые приводят к изменению взаимосоответствия развивающихся структур и проходят жесткий контроль естественного отбора. Сохраняются лишь те гетерохронии и гетеротопии, которые оказываются полезными. Примерами таких адаптивных гетерохронии являются сдвиги во времени закладок наиболее жизненно важных органов в группах, эволюционирующих по типу арогенеза. Так, у млекопитающих, и в особенности у человека, дифференцировка переднего мозга существенно опережает развитие других его отделов. Гетеротопии приводят к формированию новых пространственных и функциональных связей между органами, обеспечивая в дальнейшем их совместную эволюцию. Так, сердце, располагающееся у рыб под глоткой, обеспечивает эффективное поступление крови в жаберные артерии для газообмена. Перемещаясь в загрудинную область у наземных позвоночных, оно развивается и функционирует уже в едином комплексе с новыми органами дыхания — легкими, выполняя и здесь в первую очередь функцию доставки крови к дыхательной системе для газообмена. Гетерохронии и гетеротопии в зависимости от того, на каких стадиях эмбриогенеза и морфогенеза органов они проявляются, могут быть расценены как филэмбриогенезы разных типов. Так, перемещение зачатков головного мозга, приводящее к его изгибу, характерному для амниот, и проявляющееся на начальных этапах его дифференцировки, является архаллаксисом, а гетеротопия семенника у человека из брюшной полости через паховый канал в мошонку, наблюдающаяся в конце эмбриогенеза после окончательного его формирования, — типичная анаболия. Иногда процессы гетеротопии, одинаковые по результатам, могут являться филэмбриогенезами разных типов. Например, у различных классов позвоночных очень часто встречается перемещение поясов конечностей. У многих групп рыб, ведущих придонный образ жизни, брюшные плавники (задние конечности) располагаются кпереди от грудных, а у млекопитающих и человека плечевой пояс и передние конечности в дефинитивном состоянии находятся значительно каудальнее места их первоначальной закладки. В связи с этим иннервация плечевого пояса у них осуществляется нервами, связанными не с грудными, а с шейными сегментами спинного мозга. У упомянутых выше рыб брюшные плавники иннервируются нервами не задних туловищных, а передних сегментов, расположенных кпереди от центров иннервации грудных плавников. Это свидетельствует о гетеротопии закладки плавников уже на стадии самых ранних зачатков, в то время как перемещение переднего пояса конечностей у человека происходит на более поздних этапах, когда иннервация их уже полностью осуществлена. Очевидно, в первом случае гетеротопия представляет собой архаллаксис, в то время как во втором — анаболию. Ценогенезы, филэмбриогенезы, а также гетеротопии и гетерохронии, оказавшись полезными, закрепляются в потомстве и воспроизводятся в последующих поколениях до тех пор, пока новые адаптивные изменения онтогенеза не вытеснят их, заменив собой. Благодаря этому онтогенез не только кратко повторяет эволюционный путь, пройденный предками, но и прокладывает новые направления филогенеза в будущем. ОБЩИЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ОРГАНОВ
Организм, или особь, — отдельное живое существо, в процессе онтогенеза проявляющее все свойства живого. Постоянное взаимодействие особи с окружающей средой в виде организованных потоков энергии и вещества поддерживает ее целостность и развитие. В структурном отношении организм представляет собой интегрированную иерархическую систему, построенную из клеток, тканей, органов и систем, обеспечивающих его жизнедеятельность. Подробнее остановимся на органах и системах жизнеобеспечения. Органом называют исторически сложившуюся специализированную систему тканей, характеризующуюся отграниченностью, постоянством формы, локализации, внутренней конструкции путей кровообращения и иннервации, развитием в онтогенезе и специфическими функциями. Строение органов часто очень сложно. Большинство из них полифункционально, т.е. выполняет одновременно несколько функций. В то же время в реализации какой-либо сложной функции могут участвовать различные органы. Группу сходных по происхождению органов, объединяющихся для выполнения сложной функции, называют системой (кровеносная, выделительная и др.). Если одну и ту же функцию выполняет группа органов разного происхождения, ее называют аппаратом. Примером служит дыхательный аппарат, состоящий как из органов собственно дыхания, так и из элементов скелета и мышечной системы, обеспечивающих дыхательные движения. В процессе онтогенеза происходит развитие, а часто и замена одних органов другими. Органы зрелого организма называют дефинитивными; органы, развивающиеся и функционирующие только в зародышевом или личиночном развитии, — провизорными. Примерами провизорных органов являются жабры личинок земноводных, первичная почка и зародышевые оболочки высших позвоночных животных (амниот). В историческом развитии преобразования органов могут иметь прогрессивный или регрессивный характер. В первом случае органы увеличиваются в размерах и становятся более сложными по своему строению, во втором — уменьшаются в размерах, а их строение упрощается. Если у двух организмов, находящихся на разных уровнях организации, обнаруживаются органы, которые построены по единому плану, расположены в одинаковом месте и развиваются сходным образом из одинаковых эмбриональных зачатков, то это свидетельствует о родстве данных организмов. Такие органы называют гомологичными. Гомологичные органы часто выполняют одну и ту же функцию (например, сердце рыбы, земноводного, пресмыкающегося и млекопитающего), но в процессе эволюции функции могут и меняться (например, передних конечностей рыб и земноводных, пресмыкающихся и птиц). При обитании неродственных организмов в одинаковых средах у них могут возникать сходные приспособления, которые проявляются в возникновении аналогичных органов. Аналогичные органы выполняют одинаковые функции, строение же их, местоположение и развитие резко различны. Примерами таких органов являются крылья насекомых и птиц, конечности и челюстной аппарат членистоногих и позвоночных. Строение органов строго соответствует выполняемым ими функциям. При этом в исторических преобразованиях органов изменение функций непременно сопровождается и изменением морфологических характеристик органа. Дифференциация и интеграция В эволюции органов
Основным принципом эволюции органических структур является принцип дифференциации. Дифференциация представляет собой разделение однородной структуры на обособленные части, которые в силу различного положения, связей с другими органами и различных функций приобретают специфическое строение. Таким образом, усложнение структуры всегда связано с усложнением функций и специализацией отдельных частей. Дифференцированная структура выполняет несколько функций, и строение ее сложно. Примером филогенетической дифференциации может являться эволюция кровеносной системы в типе хордовых. Так, у представителей подтипа бесчерепных она построена очень просто: один круг кровообращения, отсутствие сердца и капилляров в системе жаберных артерий. В надклассе рыб имеются двухкамерное сердце и жаберные капилляры. У земноводных впервые появляется разделение кровеносной системы на два круга кровообращения, а сердце становится трехкамерным. Максимальная дифференциация характерна для кровеносной системы млекопитающих, сердце которых четырехкамерное, а в сосудах достигается полное разобщение венозного и артериального кровотоков. Отдельные части дифференцирующейся, ранее однородной структуры, специализируясь на выполнении одной функции, становятся функционально все более зависимыми от других частей данной структуры и от организма в целом. Такое функциональное соподчинение отдельных компонентов системы в целостном организме называют интеграцией. Четырехкамерное сердце млекопитающих представляет собой пример высокоинтегрированной структуры: каждый отдел выполняет лишь свою специальную функцию, не имеющую никакого смысла в отрыве от функций других отделов. Поэтому сердце снабжено автономной системой функциональной регуляции в виде парасимпатического атриовентрикулярного нервного узла и при этом строго подчинено нейрогуморальной системе регуляции организма в целом. Таким образом, одновременно с дифференциацией наблюдается и подчинение частей целостной системе организма, т.е. процесс интергации. 13.3.2. Закономерности морфофункциональных преобразований органов
В основе филогенетических преобразований органов лежит их полифункциональность и способность к количественным изменениям функций. Практически все органы выполняют не одну, а несколько функций, причем среди них всегда выделяется главная, а остальные второстепенны. Строение такого полифункционального органа обязательно соответствует главной функции. Так, рука человека может использоваться для лазания по деревьям, плавания, даже хождения. Но основной ее функцией является трудовая деятельность. В связи с этим и строение руки в максимальной степени соответствует функции труда. Один из основных принципов эволюции органов — принцип расширения и смены функций. Расширение функций сопровождает обычно профессивное развитие органа, который по мере дифференциации выполняет все новые функции. Так, парные плавники рыб, возникшие как пассивные органы, поддерживающие тело в воде в горизонтальном положении, с приобретением собственной мускулатуры и прогрессивным расчленением становятся еще и активными рулями глубины и поступательного движения. У придонных рыб они обеспечивают также их передвижение по дну. С переходом позвоночных на сушу к перечисленным функциям конечностей добавились хождение по Земле, лазание, бегание и др. Расширение функций сопровождается специализацией, благодаря которой главной функцией становится одна из бывших ранее второстепенными. Бывшая главной функция преобразуется во второстепенную и может впоследствии даже исчезнуть. Орган при этом меняется таким образом, что его строение становится максимально соответствующим выполнению главной функции. Так, переход предков ластоногих и китообразных к водному образу жизни привел к преобразованию их парных конечностей в ласты, практически утратившие способность обеспечивать передвижение по суше. Жизнь ленивцев, представителей отряда неполнозубых, на деревьях привела к формированию у них крючкообразных конечностей, с помощью которых возможно лишь медленное перемещение по веткам в подвешенном состоянии с почти полной утратой способности движения по земле. Нередко функции, выполняемые органами, могут измениться кардинально. Так, плавательный пузырь рыб, будучи гидростатическим органом, у кистеперых рыб становится дополнительным органом дыхания, а у земноводных он преобразуется в легкое, и основной функцией его становится дыхательная. У пресмыкающихся и млекопитающих, ведущих наземный образ жизни, легкие выполняют только дыхательную функцию, но первичная функция плавательного пузыря сохраняется за легкими у крокодилов, ластоногих и китообразных, ведущих водный образ жизни, а также у наземных форм во время плавания. В других случаях видоизменения органов в связи со сменой их функций столь велики, что выполнение ими функций, бывших ранее главными, становится невозможным. Так, передние жаберные дуги предков хрящевых рыб преобразовались в челюсти, а у наземных позвоночных они стали выполнять функции звукопроводящего аппарата, превратившись в слуховые косточки (см. разд. 14.2.1). Участие их в пищеварении и дыхании стало невозможным. В прогрессивной эволюции органов очень важным является принцип активации функций. Он наиболее часто реализуется на начальных этапах эволюции органов в том случае, когда малоактивный орган начинает активно выполнять функции, существенно при этом преобразуясь. Так, крайне малоподвижные парные плавники хрящевых рыб становятся активными органами движения уже у костистых. Более часто в филогенезе наблюдается интенсификация функций, являющаяся следующим этапом эволюции органов после активации. Благодаря этому орган обычно увеличивается в размерах, претерпевает внутреннюю дифференцировку, гистологическое строение его усложняется, нередко наблюдается многократное повторение одноименных структурных элементов, или полимеризация структуры. Примером является усложнение структуры легких в ряду наземных позвоночных за счет ветвления бронхов, появления ацинусов и альвеол на фоне постоянной интенсификации его функций. Высокая степень дифференцировки может сопровождаться уменьшением количества одинаковых органов, выполняющих одну и ту же функцию, или их олигомеризацией. Это явление наблюдается, к примеру, в эволюции артериальных жаберных дуг, которые закладываются у хрящевых рыб в количестве 6—7 пар, у костных рыб их становится 4 пары, а у млекопитающих и человека сохраняются в дефинитивном состоянии лишь части 3, 4 и 6-й пар (см. § 14.4). Иногда в процессе интенсификации функций наблюдается тканевая субституция органа — замещение одной ткани другой, более соответствующей выполнению данной функции. Так, хрящевой скелет хрящевых рыб сменяется на костный у более высокоорганизованных классов позвоночных. В противоположность интенсификации и активации ослабление функций ведет в филогенезе к упрощению строения органа и его редукции, вплоть до полного исчезновения. Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 891; Нарушение авторского права страницы