|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Восприятие электрических и электромагнитных полей
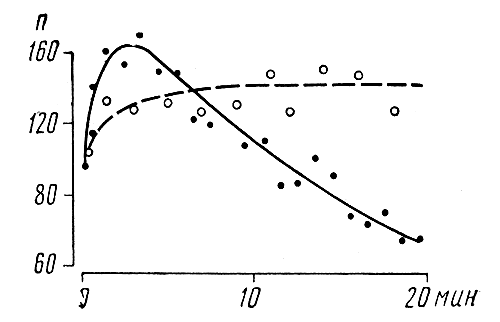
Реакции на переменные искусственно генерируемые ЭП. Действие ЭП на пчелиные семьи и группы пчел, находящихся вне жилища, стимулирует их возбуждение. Его уровень зависит от напряженности и частотно-временной структуры поля. Первые признаки возбуждения пчел под действием ЭП удалось зарегистрировать акустическим методом, для чего измеряли изменение интенсивности звуков в диапазоне 400-550 Гц. Эта полоса взята потому, что именно ее интенсивность сильнее всего изменяется при возбуждении пчел (Е. Еськов 1970, 1972, 1977). По данным акустического анализа, максимум чувствительности пчелиной семьи к ЭП приходится примерно на 500 Гц. Семья, содержащая примерно 10 тыс. взрослых особей, реагирует на ЭП указанной частоты при напряженности 4-5 В/см (Здесь и в дальнейшем за величину напряженности принимается отношение амплитуды напряжения, подаваемого на пластины, к расстоянию между ними ). Порог чувствительности к ЭП полого снижается на область низких частот. Он уменьшается примерно вдвое на частоте 10 Гц, то есть для возбуждения пчелиной семьи ЭП такой частоты требуется, чтобы его напряженность составляла 8-10 В/см. Чувствительность пчел к ЭП резко падает с повышением его частоты: на частоте 5 кГц она примерно в 10 раз ниже, чем на 500 Гц. На группах пчел (по 200 особей в группе), содержавшихся в садках из органического стекла, проследили изменение их двигательной активности с повышением напряженности ЭП. Его частота составляла 500 Гц. Установлено, что повышение напряженности сверх пороговой в начале отражается на повышении двигательной активности. Двигательная активность растет до тех пор, пока напряженность электрического поля не достигнет 150-200 В/см. Дальнейшее повышение напряженности понижает двигательную активность. Это наблюдается как у старых, так и у молодых пчел. Важно подчеркнуть, что и у летних (опыты поставлены в летний период), и у зимних (опыты проведены зимой) пчел прослеживается аналогичная зависимость изменения уровня двигательной активности от напряжения электрического поля. Двигательная активность не имеет резкого спада непосредственно после отключения электрического поля. В частности, активность пчел очень слабо уменьшается в первые 30-60 с после отключения электрического поля. Поэтому импульсный режим подачи электрического поля с паузами 30-60 с можно использовать как эффективное средство стимуляции двигательной активности пчел. Сравнительной регистрацией двигательной активности, стимулируемой действием электрического поля, подаваемого в непрерывном и импульсном режиме, установлено следующее: высокая двигательная активность сохраняется при подаче электрического поля в импульсном режиме. При этом наибольшей эффективностью действия обладают пульсации, длительность которых и интервалов между ними составляет 20-30 с (рис. 9). Подтверждением изложенных данных по регистрации двигательной активности служат результаты расхода корма пчелами, подвергаемыми действию электрического поля, подаваемого в различном режиме. В частности, за 20 мин пчелы, находящиеся под действием непрерывного электрического поля (напряженность 200 В/см, частота 500 Гц), подаваемого в непрерывном режиме, расходовали в расчете на одну пчелу около 2, 5 мг корма. В импульсном режиме (длительность импульса и паузы по 30 с) - 3, 5 мг. Не стимулированные ЭП пчелы расходовали за то же время по 1, 3 мг корма. Эти опыты провели на пчелах, успокоившихся после того, как их поместили в садки, что очень важно, так как непосредственно после перемещения пчел в садки они очень сильно возбуждаются и расходуют в первые 30-60 мин большое количество энергии. Особенно велики их энергетические затраты в первые пять минут после перемещения в садки. За этот период масса каждой пчелы уменьшается примерно на 0, 75 мг. В дальнейшем по мере успокоения пчел она снижается до 0, 25-0, 12 мг на одну особь в течение пяти минут. Уровень возбуждения пчел под действием ЭП зависит от их количества. По данным анализа интенсивности дыхания не обнаружено, чтобы одна изолированная свободно передвигающаяся особь реагировала на ЭП. Максимальная напряженность поля, действию которого подвергали пчелу, составляла 500 В/см. Интенсивность дыхания двух пчел под действием ЭП такой же напряженности возрастала в 1, 5 (Cυ - 2, 8%), десяти - в 1, 8 (Cυ - 22%) раза. По изменению интенсивности звуков первые признаки возбуждения 100 пчел под действием ЭП частотой 300-600 Гц наблюдаются при напряженности 80-200 В/см, что более чем на порядок превосходит напряженность, необходимую для возбуждения семьи. Воздействие на пчелиные семьи ЭП, напряженность которых превосходит пороговые значения, влечет за собой изменения микроклимата в их гнездах. Небольшие изменения температуры (на 0, 1-0, 3° С) и концентрации углекислоты (на 0, 05-0, 1%) в улье наблюдается в летний период под действием низкочастотного ЭП напряженностью 15-18 В/см. Значительные изменения рассматриваемых показателей микроклимата улья происходят под действием полей, напряженность которых составляет десятки вольт на сантиметр. При этом чем выше напряженность, тем на большую величину возрастает температура и концентрация углекислого газа (рис. 10). Своеобразно изменяется температура и газовый состав в пчелином жилище в зависимости от длительности действия ЭП. Например, включение ЭП напряженностью 35 В/см (частота 500 Гц) на 1 мин увеличивает температуру на 1, 2° С, а концентрацию углекислого газа - на 0, 7%. Пятиминутная стимуляция семьи таким же полем вызывает повышение температуры на 2, 7° С и концентрации углекислого газа на 1, 7%. При действии этого стимула в течение 1 ч температура возрастает на 3, 6° С, а концентрация углекислоты - на 2%. Возвращение температуры и концентрации углекислого газа к исходному уровню начинается соответственно через 18-23 и 8-10 мин после включения ЭП. Это связано с резким увеличением числа пчел, занимающихся аэрацией жилища. Усиленной аэрацией центральной зоны гнезда того места, где находится пчелиный расплод, можно объяснить также и тот факт, что именно в этой части пчелиного жилища меньше всего возрастает температура при действии на семью ЭП относительно высокой напряженности (Е. Еськов, 1972). Важно отметить, что несмотря на явно выраженную тенденцию возвращения температуры и концентрации углекислоты в улье к исходному уровню еще в период длительного действия ЭП, пчелы, тем не менее, слабо адаптируются к его действию. На это указывает относительно высокий уровень интенсивности звуков в течение всего периода включения поля. Наибольшее возбуждение рабочих особей семьи под действием ЭП наблюдается в первые 4-7 мин после его выключения. Об этом свидетельствует максимальная интенсивность звуков в указанный период. Кратковременное воздействие на семью ЭП, подаваемого в импульсном режиме, по сравнению с непрерывным, действующим в течение равного промежутка времени, не стимулирует увеличения уровня возбуждения семьи. Напротив, импульсные поля слабее возбуждают семью. Так, под действием ЭП, подаваемом в непрерывном режиме в течение 1 мин (частота 300 Гц, напряженность 79 В/см), температура над гнездом подопытных семей поднималась в среднем на 2, 1±0, 4° С, а концентрация кислорода опускалась - на 0, 9±0, 14%. Этот же стимул, подаваемый в импульсном режиме со скважностью (отношение периода следования импульсов к их длительности), равной двум и длительностью импульсов около 30 мс, вызывал повышение температуры в среднем на 1, 2±0, 2° С и понижение концентрации кислорода на 0, 5±0, 1%. При скважности, равной десяти, эти показатели микроклимата улья изменялись соответственно всего на 0, 4(-)±0, 1°С и 0, 23+0, 07%. Это противоположно эффекту действия длительных периодов пульсаций ЭП, на двигательную активность пчел, находящихся в садках. Причина альтернативности влияния пульсаций ЭП в том и другом случаях связана со следующим. Первостепенную роль играет здесь длительность импульса ЭП. В опыте на пчелах, содержащихся в садках, она составляла 20-30 с, а при стимуляции семьи - примерно в тысячу раз меньше. За такой короткий интервал пчелы не успевали сильно возбудиться. К тому же напряженность ЭП при действии на семью была примерно в три раза ниже. Следовательно, длительность действия ЭП вместе с частотой и напряженностью определяет уровень возбуждения пчел. Влияние магнитных полей. Пчелы, как и другие живые существа, подвержены действию магнитного поля (МП) Земли. По форме оно подобно однородно намагниченной сфере, ось которой составляет с осью вращения земли угол 11, 5°. МП Земли складывается из двух различных по происхождению компонентов: постоянного поля, существование которого обусловлено магнетизмом самого земного шара, и переменного поля, порождаемого токами, протекающими в верхних проводящих слоях атмосферы или даже за ее пределами. Имеются сведения, согласно которым постоянные и медленно меняющиеся ЭМП влияют на поведение и физиологическое состояние различных животных (А. Пресман, 1968; В. Протасов, 1972; Ю. А. Холодов, 1975, и др.) и в том числе насекомых. В частности, замечено, что некоторые из них, находясь в спокойном состоянии, ориентируют ось своего тела параллельно или перпендикулярно по отношению силовых линий МП Земли. Такую ориентировку можно заметить преимущественно у малоподвижных насекомых (термитов, тараканов, сверчков), а также у летающих при приземлении. Их можно дезориентировать, компенсировав ЭМП Земли с помощью кольца Гельмгольца. Но указанная особенность поведения насекомых не всегда четко прослеживается. Ряд исследователей вообще ставят под сомнение магнитную ориентацию у насекомых. В отношении магнитной ориентации пчел также нет единого мнения. Причин для разногласий много. Большая их часть связана с методическими издержками экспериментов и наблюдений. Рассмотрим примеры некоторых случаев, когда пчелы, возможно, пользуются ЭМП Земли в качестве ориентира. В 1973 г. X. Мартин и М. Линдауер опубликовали статью, в которой описали результаты наблюдений Оемке (ученик М. Линдауера). В ней сообщалось, что на выбор пчелами направления сотов при постройке гнезда влияет МП Земли. По наблюдениям Оемке, рой строил соты в том же направлении, как они были размещены в материнской семье. Позже М. Нантидис (1978) провел серию наблюдений за строительством сот в специальных ульях из глины или цемента. Первый вид опрокинутого усеченного конуса, второй - эллипсоида. Крышки того и другого улья были картонными. Через 1-4 дня после заселения этих ульев пчелами крыши многократно заменяли, вынуждая тем самым пчел начинать новое строительство гнезда. Оказалось, что ориентация сот в большинстве случаев не совпадала с их направлением в материнской семье. Одна и та же семья при смене потолка в большинстве случаев меняла направление вновь отстраиваемых сот на угол от 23 до 102°. Это указывает также на то, что пчелы не принимают во внимание направление МП Земли, так как оно не менялось в этом опыте. В статье X. Мартина и М. Линдауера приводятся сведения о том, что если с помощью постоянного магнита или катушки Гельмгольца изменить магнитное поле вокруг улья, пчелы меняют направление отстраиваемых сот. В частности, сообщается, что указанным способом удалось изменить направление сот на 40°. Отмечается также, что сильное искажение МП Земли влияет на строительную деятельность пчел. Так, если перед началом строительных работ пчел поместить в улей, рядом с которым находится металлическая шина, то пчелы начинают приклеивать соты не к верхней, а к боковой стене улья. В настоящее время имеются убедительные сведения, указывающие на то, что ЭП Земли используется в системе средств ориентации пчел-танцовщиц. Предпосылкой к изучению этого послужило явление, обнаруженное К. Фришем (1948). Он заметил, что пчелы в процессе исполнения танца (см. гл. 4) на вертикальной поверхности сот допускают «ошибки» при указании направления к цели полета. Изучая возможные причины, побуждающие пчел «ошибаться», К. Фриш и М. Линдауер (1961) заметили, что пчелы тацующие на освещенной солнцем горизонтальной поверхности сот, не ошибаются. «Ошибка» и ее знак в танце на вертикальном соте изменяются в течение дня. При этом она зависит от того, как соориентирован улей: сот расположен в направлении север - юг или запад - восток. Причину «ошибки» танцовщиц вскрыли М. Линдауер и X. Мартин (1968, 1973). Они обнаружили связь между дневной динамикой изменений МП Земли и значениями «ошибки». Уже это одно свидетельствует о влиянии МП Земли на ориентацию танцовщиц. Но исследователи пошли дальше. Они провели серию опытов, в которых компенсировали или усиливали действие на танцовщиц ЭП Земли, используя катушку Гельмгольца. Оказалось, что «ошибка» исчезала в случае компенсации ЭП Земли и увеличивалась при его значительном усилении. В случае компенсации ЭП Земли танцовщицы начинали «правильно» указывать направление не сразу, а примерно через 30 мин. В течение указанного времени происходила «нормализация» танца. При отключении катушки Гельмгольца переход танца на обычный с характерными для данного периода времени «ошибками» проходил значительно быстрее Ч всего за 10-15 мин. К действию значительного усиления ЭП Земли, влекущего за собой увеличение «ошибки», танцовщицы постепенно адаптировались. Это выражалось в постепенном уменьшении «ошибки» при длительном действии сильного ЭП. Так, заметное уменьшение «ошибки», стимулируемой 9- или 13-кратным усилением ЭП Земли, наблюдали примерно через двое суток непрерывного действия этого стимула. Механизмы восприятия ЭМП. Один из механизмов восприятия ЭП основан на притяжении или отталкивании под действием кулоновских сил механорецепторных органов, реагирующих на смещение, давление, растяжение и т. п. Роль рецепторов колебаний заряженных тел выполняют органы, контролирующие колебания антенн. Об этом говорит следующее. Антенны мертвых и анестезированных пчел притягивают к наэлектризованному диэлектрику, несущему на себе заряд любого знака. Живые пчелы, напротив, отводят антенны от наэлектризованных тел, причем тем сильнее, чем ближе заряженное тело и выше его заряд. В отличие от этого приближаемое к пчелам незаряженное тело они «ощупывают» (прикасаются) антеннами. Антенны могут воспринимать также относительно быстрые изменения напряженности ЭП. Частота ЭП, вызывающая максимальное отклонение антенн незаряженной пчелы, составляет около 50 Гц. У пчелы, заряженной любым знаком, эта частота вдвое выше. Отмеченная особенность аналогична удвоению частоты колебаний незаряженного проводника по отношению к частоте воздействующего на него ЭП. Амплитуда колебаний антенн в ЭП одной и той же частоты связана с напряженностью поля. При его частоте 50 Гц и напряженности 700 В/см антенна незаряженной пчелы отклоняется примерно на 3°, при 950 В/см - на 6°, при 1200 В/см - на 12 °С и при 1350 В/см - на 18°. С увеличением заряда тела пчелы угол отклонения антенны под действием ЭП одной и той же частоты возрастает. В ЭП напряженностью 950 В/см антенны отклоняются на 3°, 10°, 15° и 18° с ростом заряда пчелы соответственно до 150, 400, 600 и 800 пКл. Итак, антенны пчелы воспринимают перемещения заряженных тел и колебаний ЭП высокой напряженности (порядка сотен вольт на 1 см). Однако выше отмечалось, что пчелиные семьи реагируют на низкочастотные ЭП, напряженность которых составляет всего единицы вольт на 1 см. По имеющимся сведениям, восприятие слабых ЭП связано с физиологическим действием наведенных токов, протекающих в местах соприкосновения пчел друг с другом или токопроводящими поверхностями. На это указывает прежде всего то, что пчел, находящихся в ЭП, раздражают тактильные контакты. Так же они ведут себя при прикосновении к различным токопроводящим телам, помещенным в ЭП вместе с пчелами. Сильнее всего они реагируют на прикосновение, если находятся на одной прямой, параллельной силовым линиям ЭП, что можно объяснить максимальной силой тока, протекающей в месте контакта. Приняв, что восприятие ЭП связано с действием наведенных токов, воспринимаемых пчелами при их контактировании, можно объяснить неравномерность частотной характеристики и, в частности, сильное падение чувствительности с ростом частоты. Дело здесь в том, что пчелы обладают максимальной чувствительностью к току низких частот. При регистрации величины тока, побуждающего пчел, наполняющих медовый зобик раствором сахара, отдергивать хоботок, установлено, что эту реакцию стимулирует постоянный ток величиной около 0, 1 мкА. На частотах 50, 200, 500, 1000 и 2500 Гц пчелы реагируют указанным образом, когда величина тока достигает соответственно 0, 20; 0, 24; 0, 4; 0, 75 и 2 мкА. Указанная тенденция падения чувствительности пчел к току (эти величины нельзя считать пороговыми, так как только сильная стимуляция побуждает пчел прекращать заполнение медового зобика) значительно опережает рост величины тока, наводимого в ЭП, с повышением его частоты в указанном диапазоне частот. Поэтому падение чувствительности пчел к ЭП, с повышением его частоты можно объяснить ростом разницы между падением чувствительности и величиной наводимого в них тока. Восприятие переменных ЭП возможно также непосредственно через воздействие на различные системы и прежде всего нервные структуры организма пчелы. Вероятно, немаловажная роль принадлежит особенностям строения центральной нервной системы, включающей в себя нервную цепочку с двумя грудными и пятью брюшными ганглиями. По-видимому, с этим связано то, что для возбуждения одной изолированной пчелы, продольная ось тела которой зафиксирована вдоль силовых линий требуется ЭП меньшей напряженности (около 200 В/см), чем в том случае, если пчела расположена перпендикулярно к силовым линиям ЭП. Механизмы восприятия ЭМП и МП пока не имеют убедительного объяснения. На этот счет имеется несколько рабочих гипотез. Г. Беккер и У. Спек (1964), изучавшие магнитную ориентацию у мух, высказали предположение, что в теле насекомого постоянно в определенном направлении течет ток. Источником тока служит батарея, состоящая из кутикулы и внутренних тканей. Этот ток взаимодействует с внешним ЭП, что воспринимается механорецепторами. В подтверждение этой гипотезы авторы приводят результаты опытов, в которых подвешенные на нити живые или мертвые мухи ориентировались (поворачивались) определенным образом в МП. В своих работах С. И. Думбадзе и соавт. (1969) обнаружили, что пчела обладает свойствами четко выраженного диаполя. Она, будучи подвешена на нити, подобно мухе в опытах Г. Беккера и У. Спека разворачивается в поле постоянного магнита. Магнитный диполь пчелы совпадает с продольной осью ее тела. Диполь отчлененной головы перпендикулярен оси тела. Полярное магнитное свойство пчел, как и мух, не теряется после высушивания насекомых. И. Непримеров, У. Ахмеров и А. Бильдюнович (1966), основываясь на том, что вода под действием магнитного поля изменяет свои свойства, связывают с этим изменения жизнедеятельности животных под действием МП, так как вода входит в состав их органов и тканей. Один из механизмов восприятия ЭП связывают с детектирующими свойствами мембран нервных клеток (К. Коул, 1956). А. Пресман отождествляет механизмы действия ЭМП на животных с влиянием природных и искусственно генерируемых электромагнитных полей на радиосвязь. Таким образом, у пчел не обнаружено специализированного рецептора ЭМП. Изменения напряженности ЭП заряженного тела пчелы воспринимают антеннами, подвергаемыми действию кулоновских сил. Перемещение антенн, по-видимому, воспринимается джонстоновым органом. Восприятие низкочастотных ЭП связано в основном с физиологическим действием наведенных токов, которые протекают в местах прикосновения пчел друг к другу или токопроводящими телами. Видимо, на этом же принципе основано восприятие пчелами приближения грозы, перед которой, как известно, пчелы прекращают летную деятельность. Восприятие ЭМП, вероятнее всего, связано с комплексом факторов, среди которых немаловажная роль принадлежит взаимодействию электрических процессов в теле пчелы с внешним ЭМП. Ориентация в магнитном поле пчел может быть объяснена тем, что они обладают свойствами диполя.
Поиск и заготовка пищи Одной из особенностей образа жизни общественных насекомых является систематическая заготовка пищи для личинок и остальных членов семьи. Доставленная в гнездо пища используется сразу или откладывается впрок. Запасы питательных веществ во многом определяют жизнеспособность семьи. Так, запас меда позволяет пчелам пережить зиму. В семье общественных насекомых не все ее члены в одинаковой мере занимаются поиском и доставкой в гнездо пищи; у видов с высоким уровнем общественной организации этим занимаются фуражиры-разведчики и мобилизуемые ими фуражиры-новички (В дальнейшем мы будем называть фуражирами тех насекомых, которые первыми нашли источник пищи или в данный момент мобилизуют на ее доставку других сборщиков-новичков ). Понятно, что чем больше насекомых собирает пищу, тем обильнее ее запасы в гнезде. Если источник пищи находится далеко, то количество пчел, занимающихся его доставкой, зависит от того, насколько точно и быстро фуражиры оповещают и мобилизуют новичков. В системе пищевой мобилизации пчелы используют большой и сложный комплекс различных сигналов. Чтобы разобраться в этом, рассмотрим эволюцию механизмов поиска и заготовки пищевых запасов видами различных представителей пчелиных, находящихся на различных уровнях общественной организации. Заготовка пищи шмелями Накопление в шмелином гнезде запасов меда и цветочной пыльцы связано с включением в процесс добывания пищи сравнительно большого числа рабочих особей. Это побудило многих этологов искать механизмы, регулирующие активность членов шмелиной семьи, связанную с добыванием пищи. В частности, А. Бриан (1952), занимавшийся этим вопросом, заметил, что иногда сборщики пыльцы, побывав в гнезде, но не освободившись от своей ноши, вылетают и кружатся некоторое время у входа в гнездо. Это напоминает сигнальное поведение безжалых пчел. Однако другие авторы, наблюдая за внутригнездовым поведением шмелей, посетивших медоносные растения или приманки с раствором сахара различной концентрации, не обнаружили, чтобы они специфическим поведением или акустическими сигналами пытались привлечь к себе внимание и оповестить других членов семьи об обнаруженном ими источнике пищи. Шмели-сборщики, возвратившись в гнездо, сразу же складывают свою ношу в предназначенные для этой цели восковые ячейки. Как исключение, передают иногда матке-основательнице (К. Мишенер, 1972). Этот трофический контакт проявляется в своеобразной форме: матка атакует появившегося в гнезде сборщика, который отрыгивает на дно гнезда часть содержимого медового зобика, и она забирает (слизывает) этот корм. Интересные и разносторонние исследования механизмов регулирования деятельности шмелиной семьи при добывании пищи провел У. Якобс-Иессен (1959). Автор приучил 11 рабочих земляных шмелей Bombus terrestris из семьи, состоящей примерно из 500 особей, посещать кормушку с 50%-ным раствором сахара на расстоянии 6, 6 м от гнезда. В течение шести дней меченные краской шмели посетили кормушку 1157 раз, но за все это время ни один новичок не появился у кормушки. Аналогичный результат получен в 10-дневном опыте на небольшой семье полевых шмелей В. agrorum, когда меченые рабочие особи посетили кормушку 463 раза. Особенно показательны результаты, полученные У. Якобс-Иессеном в опытах на шмелиной семье (250 взрослых особей), содержавшейся в вольере, которая имела запас углеводной пищи, полностью потребляемой в течение ночи. Это вынуждало шмелей ежедневно заниматься добыванием пищи. В качестве пищевой приманки использовали раствор сахара, которым наполнили цветки живокости. Эти цветки, посещаемые четырьмя мечеными сборщиками, выставляли в вольере примерно на один час, после чего на столько же убирали. В течение каждого из этих чередующихся периодов подсчитывали количество немеченых шмелей, вылетающих из гнезда. В результате за все время опыта немеченые шмели вылетали 206 раз. Из этого числа только 31% вылетов пришелся на время, когда в вольере находились цветы, посещаемые мечеными шмелями. Итак, большая часть шмелей вылетала из гнезда в то время, когда приманка была изъята из вольера и, следовательно, пищевые запасы семьи не пополнялись. Опыты У. Якобс-Иессена убеждают, что принос сборщиками корма не только не активизирует, а, напротив, тормозит активность других шмелей по добыванию пищи. Отсюда можно заключить, что активность шмелиной семьи, связанная с добыванием пищи, регулируется ее запасами в гнезде. Это подтверждают опыты А. Бриана (1952), который стимулировал вылеты шмелей, удаляя у них пищевые запасы и затормаживая летную активность, добавляя мед или раствор сахара в медовые ячейки. Запах доставляемой в гнездо пищевой ноши, как и сама ноша, не активизирует членов шмелиной семьи. К запаху ноши сборщиков безразличны не только новички, но и особи, ранее посещавшие этот источник пищи. Запах цветков, видимо, не запоминается и не принимается во внимание шмелями-сборщиками при повторных посещениях одного и того же растения. Местонахождение медоносов, вероятнее всего, шмели запоминают исключительно по форме, цвету и пространственному расположению визуальных ориентиров. Популярное:
|
Последнее изменение этой страницы: 2016-06-05; Просмотров: 799; Нарушение авторского права страницы