|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Химическая сигнализация и ориентация
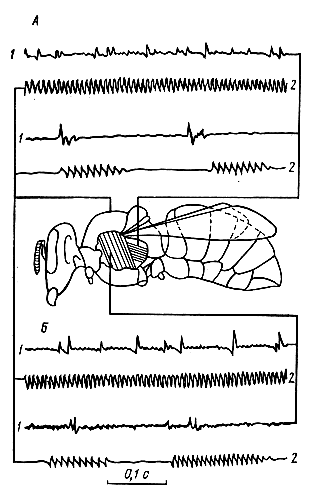
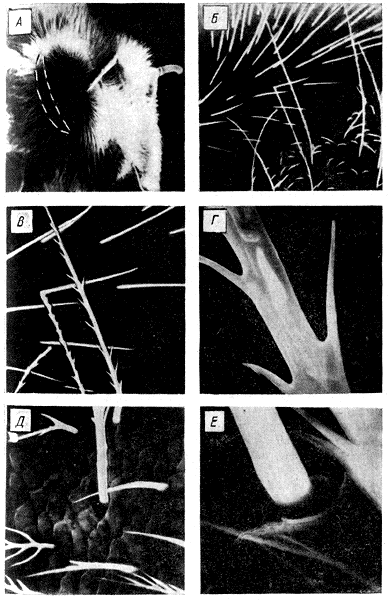
Химические вещества, используемые в системе сигнализации и ориентации пчел, можно разделить на две группы. К первой относятся те из них, которые выделяются самими пчелами, ко второй — природные пахучие вещества, входящие в состав продуктов питания и прополиса, а также выделения врагов пчел. И те, и другие используются пчелами для координации многих поведенческих актов, понимание которых невозможно без анализа роли химических сигналов. Поэтому они рассматриваются в различных главах книги. В настоящем разделе описаны лишь некоторые механизмы выделения пчелами пахучих веществ и работа хеморецепторов. Выделение пчелами пахучих веществ. Из специализированных пахучих желез хорошо изучена железа Насонова. Она имеется у рабочих пчел и матки. Железа располагается между 5 и 6-м спинными сегментами брюшка. Рабочие пчелы выпячивают железу, поднимая вверх брюшко. Это внешнее проявление функционирования железы. Многими исследователями замечено, что железа Насонова чаще всего функционирует у рабочих пчел в тех случаях, когда они посещают источники пищи, не обладающие запахом. Пчелы пользуются этой железой и в тех случаях, когда занимаются доставкой воды, запах которой может подавлять или активизировать функционирование железы. Так, Дж. Фри и Дж. Вильяме (1970) заметили, что 53—67% пчел, посещавших кормушку с дистиллированной водой, выставляли железу. При добавлении к воде пахучих веществ количество этих пчел уменьшалось до 5—11%. Секрет железы Насонова содержит гераниол, цитраль, гераниевую и неролиевую кислоты. Из названных химических компонентов наибольшей аттрактивностью (привлекательностью) для пчел обладают цис- и трансизомеры цитраля. Пахучие вещества, выделяемые насоновой железой, содержатся в нектаре и пыльце цветочных растений, которые посещают пчелы. Это позволяет предположить, что пища влияет на химическую структуру феромонов. С позиций изложенного можно объяснить явление, известное под названием «флорспециализация», выражающееся в том, что пчелы разных экологических популяций отдают предпочтение определенным видам растений из числа одновременно цветущих и выделяющих нектар и пыльцу. По-видимому, эта особенность пчел разных популяций сформировалась в процессе их длительной сопряженной эволюции с растениями. Отсюда для оптимального функционирования всего сложного комплекса физиологических процессов, протекающих в пчелиной семье, ее членам требуется нектар и пыльца определенных видов растений, характерных для условий, в которых формировалась экологическая популяция. Матка использует железу Насонова при вылетах из жилища. Пахучие выделения железы служат ориентиром для пчел, сопровождающих матку в период роения (см. гл. 1). Для обнаружения матки пчелы пользуются также выделением ее мандибулярных желез. Их выделения входят в комплекс пахучих веществ, привлекающих трутней к матке в брачный период. Их также возбуждают или привлекают выделения скоплений субэпидермальных клеток, расположенных на 2, 3 и 4-м тергитах брюшка (М. Реннер, М. Бауманн, 1964). Репелленты выделяет, как отмечалось выше, ядовитая железа. Вероятно, в число выделяемых ею репеллентных веществ входит изоамилацетат. Выделение мандибулярных желез рабочих пчел, по-видимому, также содержит репелленты. Они, по наблюдениям Д. Симпсон (1966), обладают отпугивающим воздействием на рабочих пчел. Хеморецепторы. Различные химические вещества воспринимаются хеморецепторами, которые функционально разделяются на обонятельные (ольфакторные) и вкусовые (контактные). Обонятельные реагируют на пахучие вещества, молекулы которых присутствуют в воздухе. Контактные — на вещества, растворенные в воде. Ольфакторные рецепторы находятся на восьми дистальных члениках антенн. На это указывают результаты электрофизиологических и поведенческих опытов. Пчелы с ампутированными восьмью дистальными члениками антенн не реагируют на пахучие вещества. Г. Демирчоглян и В. Непримян (1971) посредством регистрации электроантеннограмм при последовательной ампутации члеников антенн установили, что наибольшей чувствительностью обладают хеморецепторы, расположенные на шестом, седьмом и восьмом члениках. Морфологически и анатомически на антеннах выделяется семь типов сенсилл. Из них пока только плакоидные сенсиллы можно с уверенностью отнести к ольфакторным рецепторам. Разные группы этих сенсилл специализированы на восприятие определенных химических веществ. Однако то, что пчелы различают большое многообразие пахучих веществ, вероятно, связано с тем, что рабочие диапазоны различных рецепторов перекрываются. О невероятно высоком рабочем диапазоне хеморецепторного аппарата пчел говорят результаты работ Е. Вейреши (1971). В его опытах пчелы из 1816 предъявленных им парных сочетаний запахов различили 95, 5%. Плакоидные сенсиллы представляют собой овальные пластинки. Их диаметр достигает 12—14 мкм. Пластинки пронизаны порами, диаметр которых колеблется от 0, 01 до 0, 03 мкм. Каждая плакоидная сенсилла содержит от 12 до 18 рецепторных клеток. Число плакоидных сенсилл на антеннах пчел, маток и трутней неодинаково. У рабочих пчел на каждой антенне содержится от 3600 до 6000 сенсилл. У матки — около 3000, у трутня — по 30 тыс. плакоидных сенсилл. Увеличение числа сенсилл прямо коррелирует с числом афферентных импульсов, приходящих в центральную нервную систему, при одинаковой ольфакторной стимуляции. Это и приведенные выше сведения о различном количестве плакоидных сенсилл у разных представителей пчелиной семьи позволяет утверждать, что сильнее всего обоняние развито у трутней. Принято считать, что обоняние пчел развито сильнее, чем у человека. Это верно лишь для отдельных пахучих веществ. К ряду веществ обоняние пчел находится на том же уровне, что и у человека, а к некоторым — значительно ниже его (табл. 2). Контактные хеморецепторы пчелы контролируют в основном пригодность нектара, воды и других веществ для питания. Эти рецепторы находятся на ротовых частях (лабиальные пальпы), дистальных члениках ног (на лапках) и антеннах. Антеннальные контактные хеморецепторы представлены трихоидными и базиконическими сенсиллами. Вкусовые рецепторы стимулируют положительные или отрицательные реакции пчел на различные вещества. Так, горькие и кислые вещества вызывают у пчел отрицательную реакцию. Положительную пищевую реакцию стимулирует сахароза, глюкоза, мальтоза, фруктоза, трегалоза, мелецитоза, фукоза, а митилглюкозид и инозит. Привлекательность этих веществ неодинакова. Для ротовых рецепторов привлекательность глюкозы равна привлекательности мальтозы, фруктоза - равна глюкозе. Глюкоза привлекает пчел сильнее, чем фукоза, а фукоза превосходит галактозу. Лабиальные рецепторы пчел отличаются сравнительно низкой чувствительностью к концентрации углеводов. Пороговая концентрация сахара для этих рецепторов находится на уровне 0, 062—0, 125 М, что значительно ниже чувствительности контактных рецепторов, расположенных на антеннах. Последние реагируют на раствор сахарозы, концентрация которого составляет всего 0, 01—0, 07 М. Низкая чувствительность лабиальных рецепторов к сахарам объясняется тем, что пчелы приспособились потреблять высококонцентрированные растворы Сахаров. Положительная реакция пчел на растворы сахаров выражается в том, что они вытягивают хоботок и начинают забирать этот углеводный корм. Эту реакцию стимулируют контактные рецепторы, находящиеся на лапках и антеннах. Пороговая концентрация на одни и те же сахара может меняться в зависимости от физиологического состояния пчел более чем в десять раз. У голодных насекомых при прочих равных условиях вкусовые пороги значительно ниже, чем у сытых. Чувствительность вкусовых рецепторов пчел, как и ольфакторных, для одних веществ близка к вкусовой чувствительности человека, для других — сильно отличается от нее. Например, пчела начинает отличать дистиллированную воду от раствора хлористого натрия, когда его концентрация достигает 0, 24 М. Вкусовые рецепторы человека определяют наличие хлористого натрия при концентрации 0, 09 М. Пороги чувствительности для соляной кислоты у пчелы и человека одинаковы. Они составляют 0, 001 М. По чувствительности к хинину вкусовой анализатор человека примерно в 100 раз выше, чем у пчелы. Она реагирует на наличие хинина, когда его концентрация составляет всего 1-10-5. Приведенные значения чувствительности говорят о том, что она сильно варьирует в зависимости от химической структуры вещества. Хемоориентация. Расположение ольфакторных рецепторов на подвижных органах - антеннах обеспечивает возможность определять положение пахучей цели сидящим и летающим пчелам. Роль антенн в ольфакторной ориентации показана работами X. Мартина и М. Линдауера (1963, 1964, 1965). Они провели серию специальных опытов, в которых пчелы были приучены посещать ароматизированную подкормку. Способность пчел ориентироваться по запаху, к которому у них вырабатывали предварительно условный рефлекс, оценивали по выбираемому насекомыми направлению движения в У-образном лабиринте. Оказалось, что быстрее всего дорогу к цели находили интактные пчелы. Они во время поиска раздвигали в стороны антенны и двигали (скандировали) ими. После ампутации одной из антенн пчела намного медленее находила цель. Пчела с неподвижно закрепленными антеннами еще медленнее двигалась к цели. Это говорит о важности колебательных движений антенн во время поиска. Подтверждением сказанному служат результаты опыта, в котором одну из антенн удалили, а другую закрепляли на голове. Такая пчела двигалась к цели по зигзагообразной траектории. Различие в концентрации пахучего вещества, необходимое для распознания пчелой направления к нему, было определено в серии опытов X. Мартина. Для этого он надевал на антенны стеклянные капилляры, в дистальные концы которых помещали ароматическое вещество различной концентрации. По хемотропотаксической реакции (повороту на действие химического стимула) пчел было определено необходимое соотношение концентрации пахучего вещества, действующего на разные антенны. В этих опытах пчела поворачивалась, когда соотношение концентрации пахучих веществ достигало 2, 5. Следовательно, 2, 5-кратного градиента концентрации пахучего вещества достаточно, чтобы пчела могла обнаружить направление к пахучей цели. Акустическая связь Механизмы генерации звука. Генерация пчелами звуков основана на использовании летательного аппарата. По-разному используются и функционируют его элементы в зависимости от характера генерируемых звуков. Роль одного из элементов летательного аппарата — крыловых пластин была определена на основе экспериментов, в которых крылья укорачивали или совсем ампутировали. Оказывается, такие операции сильнее всего отражаются на интенсивности и частоте звуков, сопутствующих полету и вентилированию. Здесь по мере укорачивания крыльев наряду, со значительным ослаблением звуков происходит изменение соотношения интенсивностей их низкочастотного (150—500 Гц) и высокочастотного (5000—9000 Гц) диапазонов. Уменьшение крыльев наполовину влечет за собой усиление высокочастотного диапазона по отношению к низкочастотному примерно на 26 дБ. Различие их интенсивностей возрастает до 33—35 дБ при полной ампутации крыльев. При этом интенсивность низкочастотного диапазона уменьшается по сравнению с исходной примерно на 20 дБ. Изучение влияния крыловых пластин на структуру акустических сигналов, сопровождаемых едва заметными движениями крыльев в латеральной плоскости, проведено на фиксированных пчелах. Подопытные пчелы генерировали пульсирующие звуки, аналогичные по спектральным и временным характеристикам сигналам дальности. Спектр пульсирующих звуков, как и сигналов, используемых пчелами для указания расстояния до цели полета, укладывается в диапазоне 80—1000 Гц с максимумом интенсивности в области 200—400 Гц. Звук фиксированных пчел подобно сигналам дальности состоит из серии импульсов с частотой следования 20—26 Гц. Длительность одного импульса составляет 25—50 мс. Уменьшая и ампутируя крыловые пластины, определяли их роль в генерации пчелами пульсирующих звуков. Оказывается, крыловые пластины играют здесь значительно меньшую роль, чем в генерации звуков, сопровождающих полет или вентилирование. Об этом свидетельствует относительно небольшое падение интенсивности и меньшее изменение спектральной структуры пульсирующих звуков при уменьшении и ампутации крыловых пластин по сравнению с влиянием этих операций на характеристики звуков, генерируемых пчелами, машущими крыльями. Даже полная ампутация крыловых пластин снижает интенсивность пульсирующих звуков в диапазоне 200—400 Гц всего на 5—7 дБ. При этом различие между интенсивностями высокочастотных и низкочастотных составляющих увеличивается лишь на 9—10 дБ. Интересно отметить, что уменьшение и ампутация крыловых пластин не влияют на временную структуру пульсирующих звуков. Аналогичный результат получен в опытах с ампутацией крыльев у шмелей. Шмели, как и пчелы, могут издавать звуки, не разворачивая крыльев, но совершая ими небольшие колебательные движения. При этом амплитуда колебаний вершин крыльев у средних по величине рабочих особей садового шмеля Bombus hortorum достигает 1—2 мм и создается впечатление, что звук генерируется трением или вибрацией крыловых пластин. Однако данные спектрального анализа звуков, генерируемых интактными шмелями до и после ампутатации крыльев, опровергают это предположение. Действительно, даже полная ампутация крыловых пластин очень слабо влияет на спектральную характеристику звуков. Итак, роль крыловых пластин в генерации звуков двояка. С одной стороны, они издают звук, как и всякое тело, совершающее простые гармонические колебания. Основная частота этого звука определяется частотой взмаха крыла. С другой стороны, крыловые пластины выполняют роль резонаторов, усиливающих колебания, генерируемые тораксом. Анализ вибрации различных участков поверхности тела пчел, регистрируемых вибродатчиком в момент генерации звуков, показал, что сильнее других вибрируют хитиновые сегменты торакса, среди которых особенно велика амплитуда колебаний тергитов. Самые слабые колебания зарегистрированы на третьем стерпите. Одновременная регистрация вибрации торакса и звуковых сигналов позволила установить, что каждому вибрационному периоду соответствует один звуковой период, а спектральным анализом обнаружено совпадение между спектрами звуков и вибраций. Факторы, изменяющие структуру вибраций, в равной мере отражаются на характеристиках звуков, регистрируемых микрофоном. Например, уменьшение крыльев пчел, имитирующих полет, увеличивает основные частоты и звуков, и вибраций на 50—150 Гц, что связано с уменьшением нагрузки на мышцы непрямого действия, генерирующие вибрации торакса. Продольные и поперечные мышцы непрямого действия принимают неодинаковое участие в генерации различных звуков. На это указывают результаты одновременной регистрации потенциалов действия в соответствующих мышцах и возбуждаемых ими вибраций торакса. Оказывается, от продольных мышц отводится один импульс на 4, 5±0, 2 взмаха крыла, а от дорсовентральных — на 7, 1±0, 5 взмаха. Поскольку же частота взмахов крыла находится в прямой связи с количеством импульсов, генерируемых фибриллярными мышцами, изложенное свидетельствует о большей частоте сокращения продольных мышц, по сравнению с дорсовентральными в цикле одного взмаха крыльями. Способность пчел генерировать звуки с максимумом интенсивности на частотах до 600 Гц может быть объяснена тем, что торакс в этом случае вибрирует главным образом под действием продольных мышц, которые способны сокращаться в высоком ритме. Преимущественная роль продольных мышц подтверждается в данном случае тем, что в момент генерации этих звуков пчелы не машут крыльями, как при полете, а совершают ими едва заметные движения в латеральной плоскости. Пульсирующие звуки, издаваемые пчелами, также генерируются преимущественно продольными сжатиями торакса. Об этом свидетельствует то, что импульсы, генерируемые продольными мышцами, когда пчелы издают пульсирующие звуки, опережают или совпадают с началом вибрации торакса. Потенциалы действия, отводимые в этот период от поперечных мышц, чаще следуют после нескольких периодов вибрации торакса (рис. 5). Совпадение частоты движения крыльев в летательной плоскости с несущей частотой звуковых сигналов танцовщиц, обнаруженное X. Эшем (1964), еще раз подтверждает преимущество продольных мышц в издавании пульсирующих звуков. Важная роль в процессе и генерации вибраций торакса принадлежит подвижному сочленению его элементов и особенно наличию среднетергальной щели. При нарушении подвижности сочленений (например, скрепление их каплей клея) пчелы не в состоянии генерировать звук. На основании этого можно объяснить результаты опытов Дж. Симпсон (1964), в которых матка переставала петь в случае заклеивания первой пары грудных дыхалец. Дело здесь в том, что заклеивание названных мезотораксальных дыхалец приводит к жесткому закреплению прикрывающих их плейральных выростов проторакса, перемещающихся в период генерации пульсирующих звуков относительно сегментов мезоторакса. Торакс становится неподвижным и не может сжиматься и вибрировать. Генерирование звука, основанное у пчел на вибрации торакса крыловой мускулатурой непрямого действия, имеет некоторое сходство с принципом возбуждения колебаний тимбальных мембран у цикад. Но в отличие от пчел цикады издают звуки довольно стабильного спектрального состава на частотах от 1, 5 до 10 кГц. Спектры их звуков определяются в основном размерами тимбальных мембран, которые неодинаковы у разных видов Поэтому у цикад, имеющих тимбальные мембраны приблизительно одинакового размера, максимум энергии звуков сосредоточивается примерно в одном и том же диапазоне частот. Пчелы же, несмотря на малые размеры тела, способны генерировать низкочастотные звуки с максимумом энергии в диапазоне от 75 до 600 Гц. К тому же звуковой аппарат пчел обладает большими возможностями для широкой перестройки характеристик генерируемых сигналов. Восприятие звука. В системе акустической сигнализации в качестве линий акустического канала связи пчелы используют воздух или твердые тела. В принципе любой звуковой сигнал, независимо от способа генерации может передаваться по каждой из этих линий. Однако с точки зрения экономии энергетических затрат акустические сигналы целесообразно передавать, минуя переходы через среды с различной акустической проводимостью (воздух, твердые тела), что, по-видимому, повлияло в филогенезе на организацию системы акустической сигнализации пчел. Действительно, они обычно используют воздушную среду в тех случаях, когда звук генерируется без участия твердого опорного субстрата. Твердый субстрат как линию связи пчелы используют для передачи сигналов, генерируемых посредством воздействия на субстрат. Это достигается тем, что звукоиздающий орган контактирует (прижимается) к субстрату. Воспринимают пчелы сигналы, передаваемые по воздуху и через субстрат, двумя различными типами рецепторов. Роль рецепторов, воспринимающих колебания субстрата (виброрецепторы), выполняют субгенуальные органы, а распространяющихся через воздушную среду — специализированные фонорецепторы, расположенные на голове. Виброрецепторы. Их роль выполняют субгенуальные органы. Они располагаются в верхних частях голеней всех трех пар ног. Чувствительные элементы этих органов — длинные сколопидии - подвешены в двух точках. В одной из них клетки собраны, а в другой - свободно висят и поэтому выглядят как полураскрытый веер. Каждый субгенуальный орган содержит 48—62 сколопидия. Колебания субстрата, передаваемые через ногу сколопидиям, стимулируют их возбуждение. По данным электрофизиологических исследований X. Аутрума и В. Шнейдера (1948), чувствительность субгенуальных органов рабочих пчел наиболее высока в Диапазоне 1—3 кГц. Максимум чувствительности находится на частоте 2500 Гц и составляет около 13 нм. С понижением частоты чувствительность органа значительно падает и на частоте 400 Гц составляет 250—2000 нм. X. Ф. Литтл в поведенческих опытах обнаружил, что пчелы обладают сравнительно высокой чувствительностью к колебаниям субстрата в диапазоне 500—1000 Гц. Порог чувствительности в этом диапазоне частот составляет примерно 50 нм, что, как и частотная область максимальной чувствительности, несколько отличается от обнаруженной в изложенных выше электрофизиологических опытах. Различия обусловлены, видимо, неравнозначностью методов этих исследований. Фонорецепторы. Роль фонорецепторов у пчел выполняют волосковые сенсиллы, расположенные между фасеточными глазами и затылочным швом. Эти волоски дугообразно изогнуты или имеют волнистую форму. Их длина составляет 0, 640±0, 04 мм (Cυ — 27%). Поверхность волосков покрыта шиловидными выростами (остями). От вершины к основанию волосков количество остей постепенно уменьшается, а их размер увеличивается (рис. 6). Волосок колеблется под действием звуков за счет прогиба сочленованной мембраны. Под микроскопом это выглядит как «размыв» волоска, увеличивающийся от основания к вершине. Угол отклонения волосков от положения равновесия возрастает с увеличением интенсивности звука и зависит от его частоты. Высокочастотная граница звуковых колебаний интенсивностью до 80 дБ, отклоняющих волосок на один градус, находится между 200—400 Гц. С уменьшением частоты звука при неизменной интенсивности угол отклонения волосков увеличивается, а после перехода через некоторый максимум вновь уменьшается. Низкочастотный резонансный максимум наиболее высокочастотных волосков приходится на 125—180 Гц. В этой области они отклоняются на один градус под действием звуков интенсивностью около 64 дБ. Резонансный максимум самых низкочастотных волосков находится ниже 100 Гц. На этой частоте они отклоняются на один градус под действием звуков интенсивностью не более 55 дБ. На звуки, отклоняющие волосок, рецепторы отвечают серией нервных импульсов. Вместе с тем зарегистрированы незатухающие электрические колебания, повторяющие форму звуковой волны («микрофонный эффект»). Спонтанная импульсация в фонорецепторах пчел не обнаружена. Амплитуда импульсов, генерируемых фонорецепторами, составляет 765±31 (Cυ — 27%) мкВ, длительность их положительной части 1, 18±0, 06 (Cυ — 16%) мс. (рис. 7). Формула импульсов не зависит от структуры звуковых стимулов. Но количество импульсов в ответе, длительность интервалов между импульсами, скрытый период и скорость восстановления чувствительности рецепторов зависит от структуры звуковых стимулов. Количество импульсов, генерируемых фонорецепторами в ответ на звуковые стимулы равной длительности и интенсивности, наибольшее в зоне резонансного максимума волоска (см. рис 12). Волосковые сенсиллы, занимающие по своим резонансным свойствам некоторое промежуточное положение между высокочастотными и низкочастотными, при изменении частоты звуков длительностью около 100 мс со 150 до 350 Гц реагируют уменьшением количества импульсов в ответе с 7, 2±0, 4 (Cυ — 14%) до 2, 2±0, 2 (Cυ — 20%). Интенсивность звуков при прочих равных условиях сильно влияет на количество импульсов, генерируемых рецептором. Например, на 4-7 стимулов интенсивностью 70 дБ (длительность звуковых сигналов 100-150 мс, частота 200-300 Гц) рецепторы отвечают всего одним импульсом. На аналогичные по временной структуре и частоте звуки интенсивностью 83 дБ эти же рецепторы отвечают на каждый стимул 3-5 импульсами. Высокая чувствительность фонорецепторов с изменению интенсивности звука и их расположение по бокам головы указывают на наличие у пчел бинаурального слуха. Это подтверждается результатами поведеньческих опытов, в которых показано, что пчелы ориентируются на звук при низкой освещенности. На структуру электроотсвета рецептора влияет длительность звукового стимула. С увеличением продолжительности воздействия непрерывным звуковым сигналом количество импульсов, генерируемых рецепторов вплоть до длительности звуковой стимуляции, соответствующей периоду его адаптации, неуклонно возрастает (см. рис. 12). При этом в ответах на сигнал сравнительно большой длительности (более одной секунды) четко выражено снижение частоты импульсации, особенно в конце разряда. Небольшое влияние на количество импульсов в ответе рецептора оказывает температура, изменения которой укладываются в диапазоне колебаний внутриульевой и внешней, в период, когда пчелы ведут активный образ жизни. Так, на повышение температуры с 20-25° до 35-38 °С количество импульсов, генерируемых рецептором на звуковой стимул длительностью 240 мс (частота звука - 250 Гц, интенсивность - 76 дБ), возрастает соответственно с 3, 7±0, 2 (Cυ — 27%) до 4, 5±0, 3 (Cυ - 35%). Время адаптации рецепторов тесно связано с интенсивностью и временной структурой звуковых сигналов. К непрерывным звуковым раздражителям интенсивностью 75 дБ рецепторы адаптируются за 0, 6-1 с. С увеличением интенсивности звука до 85 дБ время адаптации возрастает в 2-3 раза. Время адаптации возрастает еще больше в случае стимуляции прерывистыми звуковыми сигналами. Так, на звуки, интенсивность которых близка к пороговой, длительностью 100 мс и частотой повторения 5 Гц, импульсный ответ рецептора практически не меняется в течение 4-7 с. Скорость восстановления чувствительности и продолжительность латентного периода ответа фонорецепторов находятся в обратной зависимости от интенсивности звуков. После адаптации к звукам интенсивностью около 75 дБ на частоте 200-300 Гц рецептор отвечает на повторную стимуляцию не раньше чем через 0, 3 с. Для получения ответа на сигнал интенсивностью 80 дБ эта пауза должна быть не менее 0, 6 с. Что же касается латентного периода, то его продолжительность уменьшается с 80±9 (Cυ - 35%) до 35±2 (Cυ - 20%) мс при увеличении интенсивности звукового воздействия частотой около 250 Гц с 73 до 83 дБ (см. рис. 7). Амплитуда электрических колебании «микрофонного эффекта» не имеет явно выраженного порога, находясь в прямой зависимости от интенсивности звукового стимула и в обратной от его частоты. В области 100-150 Гц амплитуда «микрофонного эффекта» максимальна и при интенсивности звука 80 дБ составляет 300-350 мкВ. С повышением частоты звука до 300-350 Гц его амплитуда уменьшается в 2-3 раза. Происхождение «микрофонного эффекта» и его роль в восприятии пчелами звуков не имеет пока убедительного объяснения. Относительно его происхождения можно предположить, что он, возможно, связан со смещениями в микроструктурах фонорецептора, вызываемых колебаниями волоска, Не исключено, что эти электрические колебания имеют отношение к трансформации механических колебаний волоска в нервное возбуждение. Своеобразен ответ фонорецептора на отгибание волоска. Он выражается в генерации разряда импульсов, длительность которого зависит от угловой скорости и конечной величины угла отклонения волоска. При отклонении волоска на 15-20° со скоростью примерно 100° в секунду рецептор генерирует 9±0, 5 (19, 5%) импульсов. С увеличением угла отклонения до 30-40° количество импульсов возрастает до 20±1 (Cυ - 25%). Длительность пауз между импульсами в ответах на такую стимуляцию составляет 13, 5±0, 4 (Cυ - 43%) мс. Специфической функциональной особенностью фонорецепторов пчел является то, что они отвечают генерацией импульсов только в период движения волоска - отклонения его от положения равновесия или возвращения в исходное положение. Важно также, что отогнутые волосковые сенсиллы, лишенные возможности колебаться под действием звука, не отвечают на звуковую стимуляцию. Популярное:
|
Последнее изменение этой страницы: 2016-06-05; Просмотров: 746; Нарушение авторского права страницы