|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Получение энергии путём субстратного фосфорилирования. Брожение.
Субстратным фосфорилированием называется процесс бескислородного присоединения к молекуле субстрата остатков фосфорной кислоты с последующим их переносом на АДФ, в результате чего образуется АТФ. Субстратное фосфорилирование составляет сущность гликолиза, кетодезоксифосфоглюконатного и пентозофосфатного путей. Все реакции субстратного фосфорилирования локализованы в цитоплазме бактериальной клетки.
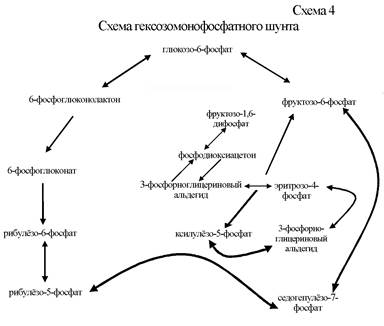
Гликолиз (путь Эмдена-Мейерхофа-Парнаса). Это наиболее примитивный способ извлечения энергии, поскольку из субстрата в результате его анаэробного преобразования извлекается лишь незначительная доля той химической энергии, которая в нём содержится. При гликолизе донором электронов является углевод, акцептором электронов – какая-нибудь органическая молекула, образующаяся в процессе последующего брожения. Синтез АТФ при гликолизе осуществляется на уровне субстрата. В результате расщепления 1 молекулы глюкозы при гликолизе расходуется 2 и синтезируются 4 молекулы АТФ (схема 3). Общий полезный выход составляет 2 молекулы АТФ и 2 молекулы НАД*Н2. Ключевым продуктом гликолиза выступает пировиноградная кислота (пируват), которая в зависимости от метаболических особенностей конкретных микроорганизмов может подвергаться дальнейшим ферментативным преобразованиям – брожению. Конечный продукт брожения является восстановленным соединением, акцептировавшим электроны, выделяется из клеток прокариот в окружающую среду и накапливается в ней в значительных количествах. Преобладающий продукт брожения определяет его вид. Молочнокислое брожение. Гомоферментативное молочнокислое брожение вызывают бактерии родов Streptococcus, Lactobacterium, Bifidobacterium. При этом виде брожения акцептором электронов служит пируват. Конечный продукт – молочная кислота. Бактерии родов Escherichia, Proteus, Salmonella, Shigella способны вызывать гетероферментативное молочнокислое брожение. При нём, кроме молочной кислоты, в меньших количествах образуются уксусная, муравьиная, янтарная кислоты, этанол. Пропионовокислое брожение вызывают бактерии родов Clostridium, Propionobacterium, Corynebacterium, некоторые виды из родов Neisseria, Veillonella. Конечными продуктами брожения являются пропионовая, уксусная, янтарная кислоты. Маслянокислое брожение характерно для бактерий родов Butyribacterium, Sarcina, Clostridium, Neisseria. При данном виде брожения образуются масляная кислота, бутанол, ацетон, изопропанол, уксусная кислота, этанол. Бутилен-гликолевое брожение свойственно некоторым видам бактерий родов Aeromonas, Bacillus, Enterobacter. Конечные продукты брожения: ацетон, бутилен-гликоль, молочная кислота, муравьиная кислота. Определение в среде ацетона (ацетилметилкарбинола) в реакции Фогеса-Проскауэра используется при идентификации Enterobacter sp. Спиртовое брожение способны вызывать бактерии родов Sarcina, Erwinia. У многих клостридий и энтеробактерий также среди продуктов брожения обнаруживается этиловый спирт. К альтернативным путям катаболизма углеводов относятся пентозофосфатный и кетодезоксифосфоглюконатный пути. Пентозофосфатный путь возник для обеспечения прокариот пентозами (схема 4). Суммарно весь процесс можно представить в виде следующего уравнения: глюкозо-6-фосфат + 2 НАДФ+ = рибозо-5-фосфат + СО2 + 2 НАДФ*Н2. Образовавшийся рибозо-5-фосфат используется прокариотами в синтезе нуклеиновых кислот и пентозосодержащих коферментов; моле
кулы НАДФ*Н2 потребляются в восстановительных биосинтетических процессах. Ни на одном из этапов пентозофосфатного пути АТФ не синтезируется. Пентозофосфатный путь присущ большинству микроорганизмов. Он в 2 точках может смыкаться с гликолизом и, следовательно, оба этих пути взаимно дополняют друг друга при необходимости образования таких ключевых веществ, как глюкозо-6-фосфат и 3-фосфорноглицериновый альдегид, которые могут переключить пентозофосфатный путь на гликолитический. Пентозофосфатный путь позволяет прокариотам сбраживать пентозы. Кетодезоксифосфоглюконатный путь (КДФГ-путь, Энтнера-Дудорова). КДФГ-путь функционирует у довольно широкого круга прокариот, главным образом у грамотрицательных (энтеробактерии, псевдомонады, спириллы и др.) (схема 5). У анаэробов он встречается довольно редко. Путь имеет важное значение при катаболизме глюконовой, мангнановой, гексуроновых кислот и их производных. Конечными продуктами КДФГ-пути являются пировиноградная кислота и 3-фосфорноглицериновый альдегид. При разложении молекулы глюкозы в КДФГ-пути образуется 1 молекула АТФ и по 1 молекуле НАДФ*Н2 и НАД*Н2. КДФГ-путь имеет несколько точек сопряжения с гликолизом (глюкозо-6-фосфат, 3-фосфорноглицериновый альдегид, пирофиноградная кислота) и пентозофосфатным путём. КДФГ-путь является наиболее коротким путём получения пировиноградной кислоты. Прокариоты,
расщепляющие глюкозу КДФГ-путём, не имеют ферментов, приводящих к бродильным процессам. Анаэробное дыхание, или субстратное фосфорилирование, сопровождающееся брожением конечных продуктов гликолиза - в 9-18 раз менее выгодный способ получения энергии по сравнению с окислительным фосфорилированием. Популярное:
|
Последнее изменение этой страницы: 2016-07-14; Просмотров: 1776; Нарушение авторского права страницы