|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Глава 6. Интересно разобраться, что же представляет из себя порода с точки зрения генетика
Когда то, многие тысячелетия тому назад, существовал вид рода Canis, предок современных собак. Он представлял большую, самостоятельно развивающуюся популяцию. В большой популяции, близкой по своим характеристикам к идеальной, частота аллелей генов остается постоянной на протяжении многих поколений. Это положение носит название закона Харди – Вайнберга. Доказывается этот закон математически просто. Допустим, что в большой (идеальной) популяции один из генов представлен двумя аллелями: доминантным А и рецессивным а. Но в популяции уже есть особи генотипов АА, Аа и аа. Они появились в процессе смены поколений, предшествующих родительскому. Каковы же будут частоты генотипов у детей взятого нами родительского поколения. Таблица 10 - Частоты гамет
Или
1 – частота а = q=0.05; 2 – частота генотипов aa = q2=0.052=0.0025 Рисунок 2
Рисунок 3 Большие популяции диких животных всегда равновесны, и это равновесие поддерживается в течение многих поколений. В больших популяциях, как правило, сосуществует два или несколько аллелей одного гена, различающихся пропорциями частот, остающимися постоянными (и равновесными) в течение многих поколений. Наличие в равновесной популяции двух и более аллелей одного гена носит название полиморфизма. Все виды животных полиморфичны. Популяции диких, еще не одомашненных, собак были полиморфны. Но вот вид подвергся одомашниванию. Популяция разобщилась на отдельные группы со случайным набором разных аллелей одних генов. Породообразование началось с образования многих, очень небольших по количеству животных, племенных ядер со случайным набором аллелей. Такие племенные ядра по генетическим процессам напоминают развитие малых популяций в условиях изоляции. В малой популяции естественный отбор быстрее начинает устранять летальные и полулетальные аллели. Но в малой популяции увеличивается случайность в накоплении отдельных генотипов. Случайно одни аллели могут исчезнуть, а другие случайно сохраниться. При последующем увеличении численности этой популяции число случайно сохранившихся аллелей может быстро возрасти, подобное изменение генных частот в малой популяции в результате действия случайных факторов называют дрейфом генов или генетико-автоматическим процессом. Но в породообразование вмешался человек. Он уже по своему усмотрению начал убирать из породы одни аллели и размножать другие. Порода, таким образом, представляет искусственно созданную неравновесную популяцию, поддерживаемую человеком. Главное генетическое различие между породами собак состоит в том, что породы различаются по частотам аллелей одних и тех же генов, определяющих те или иные признаки. У каждой из пород собак есть все гены присущие виду. Но аллели этих генов могут быть разными, присущими конкретной породе. Так у всех собак с полной выраженностью окраса работает ген С, у шнауцеров его аллель cch переводит черно-подпалый окрас в «перец с солью». У всех собак сплошного окраса без белых пятен генотип содержит ген S, а пегие гончие несут его рецессивный аллель sp. Существует доминантный ген, контролирующий нормальное развитие конечностей, и его рецессивный аллель, определяющий коротконогость такс, бассетов, некоторых терьеров. Однако природа стремится к равновесности. И если бы однажды собаководы выпустили на улицу всех собак всех пород и дали бы им возможность свободно и равновероятно скрещиваться, то уже первое поколение, полученное от таких скрещиваний, образовало бы равновесную популяцию. Кстати, равновесными являются популяции свободно гуляющих кошек в больших городах. По зоологической номенклатуре порода может быть приравнена к подвиду. Все представители одной породы обладают большим сходством друг с другом. Породам свойственна константность – стойкость в наследовании характерных признаков. Однако и самые константные породы полной однородности не имеют, так как велика и изменчивость. С одной стороны, изменчивость представляет угрозу, так как при разведении и неудачном спаривании собак можно потерять все, что было достигнуто. С другой стороны, изменчивость – залог совершенствования породы, так как каждое поколение представляет собой материал для селекции. Достаточно посмотреть на фото доберманов, шнауцеров, эрделей начала ХХ века, чтобы увидеть как они изменились. Способность породных животных стойко передавать свои качества потомству объясняется тем, что многие гены у них уже перешли в гомозиготное состояние. Однако если слишком много генов перейдет в гомозиготное состояние, то в породе наступит постоянство признаков, и она утратит способность изменяться. Чрезмерная генетическая однородность останавливает развитие породы. В чистопородном разведении применяются следующие виды спариваний: В качестве методов разведения выступают: Большинство культурных пород собак, сложившихся в ХХ веке, упираются в очень ограниченное число родоначальников и выдающихся предков, на которых весьма тесно инбридировали. В собаководстве считается, что спаривание особей, у которых общий предок находится в шестом ряду родословной, можно считать аутбредным, так как его влияние минимально, хотя у части потомства и может проявиться в виде рецессивного признака. Спаривая относительно неродственных животных, руководитель разведения в какой-то мере может избавить себя от критики: «Вот надо же! Аутбредный подбор, а нежелательная аномалия выскочила». Слабое утешение. Ведь руководитель разведения знает, что в культурных молодых породах все собаки в той или иной степени родственники. При неродственных (аутбредных) спариваниях влияние предков на генотип родившегося потомка следующее: родившаяся особь имеет 100 % своих генов, из которых 50 % получено от отца и 50 % - от матери. Но в отцовские 50 % попало 25 % генов деда и 25 % генов бабушки по отцу. Дедовские 25 % включают в себя по 12.5 % генов прадеда и прабабки и так далее в глубь прошлых поколений. Это можно представить в виде следующей таблицы. Таблица 11 - Особь имеет 100 % своих генов, из них получено от каждого предка
Неродственное спаривание более понятно широкой публике по сравнению с родственным, ведь всем известно, что супруги не должны быть родственниками друг другу. Однако в истории человечества имеются примеры близкородственных браков в течение нескольких поколений. Не смотря на это, браки двоюродных родственников считаются уже опасными для потомства. Известно, что при неродственных спариваниях домашних животных с большей вероятностью можно получить здоровое, жизнеспособное потомство. Замечено даже, что потомки от неродственных браков легче приспосабливаются к изменяющимся условиям жизни; они более умны, проявляют большую склонность к проявлению элементарного рассудка. Однако в пометах, полученных от аутбредных сочетаний, наблюдается значительный разброс потомков по фенотипу, что не нравится заводчикам, стремящимся к единообразию и сходству выпускаемых ими собак. Кроме того, заводчик, систематически работающий только при помощи аутбридинга, очень скоро обнаруживает, что ему не хватает высококлассных кобелей, чужекровных для его сук. Использование же кобелей среднего качества, только ради сохранения аутбридинга, как правило резко ухудшает качество потомства. Поэтому родословные собак, полученных аутбредно в ряде поколений, часто напоминают слоеный пирог, в котором собраны клички всех известных в стране производителей. В качестве примера приведу родословную, типичную для породы эрдельтерьер в 1970-х годах.
Рисунок 4 У миттельшнауцеров положение иное. Первый помет шнауцеров в СССР родился в 1979 году от голландской суки Элске в. д. Кемпваарт и австралийского кобеля Урангелине Беовульфа. Вскоре последовали импорты из ГДР, ФРГ, Чехословакии, Польши, Англии, Швеции, Финляндии. И эти производители поочередно перекрывали друг друга, представляя классическую картину бессистемного разведения по стране в целом. Постоянное спаривание с чужекровными производителями позволяет сохранить в потомстве высокий уровень гетерозиготности, и благодаря ей высокий уровень изменчивости. Это выгодно разведенцу, так как селекция оказывается результативной. С другой стороны гетерозиготные производители оказываются не препотентными, а это уже мешает работе разведенца. Ограниченное число выдающихся особей – хороших производителей – также вынуждает разведенца прибегать к родственным спариваниям, то есть к инбридингу. Главное генетическое последствие инбридинга заключается в том, что у животных, полученных в результате родственного спаривания, большое число пар генов становятся гомозиготными, независимо от типа действия этих генов, доминантного или рецессивного. Инбридинг вскрывает наследственные качества родительской пары. Он не увеличивает число рецессивных аллелей, а переводит их в гомозиготное состояние, заставляя таким образом проявляться в фенотипе. Применяя инбридинг, особенно тесный, нужно быть готовым к выбраковке большого числа потомков. В нескольких первых инбредных поколениях инбридинг как бы взламывает наследственность потомства, которое становится разнотипным, так как выщепляются все новые гомозиготы по рецессивам, резко меняющие фенотип. Так все тесные инбридинги, осуществленные в породе эрдельтерьер в 1970 – 1980 годы, выявили лишь отрицательную наследственность. В линиях, чистых от вредных рецессивных генов, инбридинг особого вреда не приносит. Однако, при возрастании степени гомозиготности, при многократном повторении инбредных спариваний у потомков снижается уровень развития признаков, связанных с приспособляемостью и жизнестойкостью. Увеличивается смертность эмбрионов и щенков, снижается плодовитость, ухудшаются материнские качества сук, замедляется рост и физическое развитие потомства. Этот процесс носит название инбредной депрессии. Не лучшим образом влияет инбридинг и на нервно-психическую конституцию животных, причем особенно страдает элементарная рассудочная деятельность, закрепляются нежелательные поведенческие черты. В целом нервная система слабеет. При помощи инбридинга могут быть закреплены экстерьерные особенности выдающегося производителя у потомства. Но так как при инбридинге переходят в гомозиготное состояние разные гены, то закрепляются не только положительные, но и нежелательные признаки. Существует еще одна причина, из-за которой не следует увлекаться многократно повторяемыми тесными инбридингами, переводящими значительную часть генов в гомозиготное состояние. Еще в 1903 году Иогансен обнаружил, что при постоянном инбридинге в ряде поколений образуются так называемые чистые линии по доминантным и рецессивным генам, а количество гетерозигот уменьшается, и составляет На основании этих наблюдений Иогансен вывел закон о бесполезности селекции в чистых гомозиготных линиях и об эффективности селекции только в генетически разнородных, гетерозиготных популяциях. Таким образом, чтобы получить препотентного производителя, стабильно передающего свои качества потомству нам нужен инбридинг. Но чтобы эффективно произвести отбор среди полученного потомства и иметь возможность при помощи отбора выявить лучшие генотипы необходимо, чтобы потомство оставалось достаточно гетерозиготным. А для того чтобы иметь возможность проводить селекцию в длинном ряду поколений необходимо, чтобы падение гетерозиготности и возрастание гомозиготности было медленным. Отсюда гомозиготность и гетерозиготность необходимо измерять. Самый простой способ обозначения, но не измерения, уровня инбридинга предложен Шапоружем. Он нашел широкое применение в собаководстве. Чтобы собака считалась инбредной необходимо, чтобы общий предок встречался как на отцовской, так и на материнской половине родословной. По методу Шапоружа общего предка записывают римскими цифрами, обозначающими вертикальные ряды родословной, в которых встречается его кличка. Причем первым записывается вертикальный ряд из родословной матери.
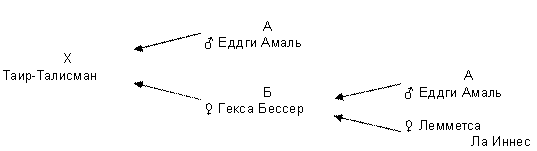
Рисунок 5 - Таир-Талисман заинбридирован на Еддги Амаля II – I
Рисунок 6 – Орлон заинбридирован комплексно на Арко IV - III, Уайта IV - III, Аликс IV – IV Как было сказано выше, общий предок должен встречаться в родословной изучаемого животного (пробанда) как со стороны отца, так и со стороны матери. Если предок встречается несколько раз только со стороны отца или только со стороны матери, то такое животное неинбредно. Например, в данной родословной заинбридированы родители Бублика. Его отец Таир-Талисман на Еддги Амаля II – I и мать Джульетта Аймон заинбридирована на Вирджинию Дарви III – II, но не сам Бублик.
Рисунок 7 – Бублик свободен от инбридинга Способ записи инбридингов по Шапоружу лишь приблизителен; он не позволяет судить о действительной степени тесноты инбридинга, сравнивать уровни инбридинга у разных животных, вычислять уровни инбридинга для групп животных и в целом по популяции. Глава 7. На вероятный уровень повышения гомозиготности, то есть на вероятный процент генов, перешедших в гомозиготное состояние, по сравнению с процентом гомозиготных генов, имеющих место во всей популяции животных, указывает коэффициент Райта. Коэффициент Райта изменяется от нуля, что соответствует полной гетерозиготности, до полной, теоретически возможной гомозиготности, равной единице или 100 %.
Рисунок 10 - а) вот их родословная; б) а так выглядит стрелочная диаграмма
Рисунок 11 - а) родословная; б) стрелочная диаграмма
Рисунок 12 - В этой родословной пробанд Х является результатом двух последовательных скрещиваний с кобелем А; у предков Х два акта инбридинга: первый - вязка А с его дочерью Б (путь Х - Б), второй - вязка А с его дочерью Д (путь Х - Б - Д)
Рисунок 13 - Пробанд Х - продукт двойного инбридинга (на В и на Г) Таблица 12 - Вспомогательная таблица значений 0.5, возведенной в степень
В случае если особь, на которую производится инбридинг, сама инбредна, то формула коэффициента Райта приобретает следующий вид:
Рисунок 14 - а) родословная; б)стрелочная диаграмма А теперь рассмотрим родословную, в которой осуществлялись бессистемные инбридинги, как это часто случается в собаководстве.
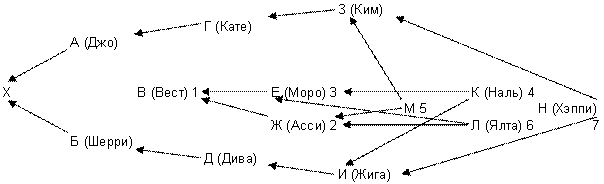
Рисунок 15 - Родословная гипотетическая, все клички взяты произвольно Для того, чтобы стрелочная диаграмма была менее громоздкой, заменим клички буквами.
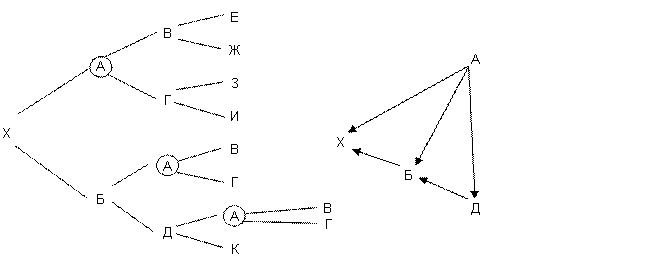
Сначала составим список общих со стороны отца и со стороны матери предков. В родословной их 7. Вест В, Асси Ж, Моро Е, Джой М, Ялта Л, Наль К и Хэппи Н. Затем нужно определить число разных путей, соединяющих одного и того же предка с пробандом со стороны отца и со стороны матери. В IV колене родословной чаще всего встречается Ялта Л. Общее число выходов на нее может быть: Выходов на Л (Ялту) всего девять. Но только три из них удовлетворяют правилу, что на пути инбридинга каждая кличка должна встречаться только один раз. Это ЖВАХБДЕ, ЖГАХБВЕ и ЖГАХБДЕ. Часть общих предков может иметь лишь один путь инбридинга, как например Вест, а другие и несколько, как Ялта (Л). Таблица 13
Примечание – Вест сам заинбридирован на Ялту (Л), и это усиливает гомозиготность Рэма. Если FX (степень гомозиготности) меняется от 0 до 1 (или от 0 до 100 %), то степень гетерозиготности представляет собой 1? FX. Ни того, ни другого практически не бывает. Но нужно стремиться, чтобы в ряду поколений гетерозиготность уменьшалась, а гомозиготность увеличивалась медленно. Для более быстрых расчетов можно пользоваться следующей таблицей. Важно только учитывать все встреченные в родословной инбридинги. Таблица 14 - Значение коэффициентов инбридинга в процентах
Таблица 15 - Классификация тесноты инбридинга
При отдаленных инбридингах, конечно, могут проявиться в фенотипе потомка скрытые рецессивные гены, но особого значения для целенаправленного применения отдаленные инбридинги не имеют. Глава 8. Коэффициент генетического сходства (родства) Иногда интересно знать степень родства между двумя особями, то есть выяснить, насколько вероятно, что эти две особи обладают одинаковыми копиями генов. Например, имеется родственник известного производителя, находящегося в другом питомнике. Если окажется, что родственник и производитель имеют высокий коэффициент родства (генетического сходства), то можно использовать на племя этого родственника. Кроме того, при спаривании двух родственных особей, чем теснее связаны родством две спариваемые особи, тем больше они должны иметь общих генов, и тем большая вероятность, что они передадут их потомству, потомок будет гомозиготным по этим генам. Чем больше общих генов имеют родители вследствие родства, тем более гомозиготным будет их потомство. Коэффициент генетического сходства между животными X и Y рассчитывается по формуле: Если особи X и Y и их общий предок не инбредны, то формула приобретает более простой вид: RXY=? 0.5n Вот несколько примеров: Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 890; Нарушение авторского права страницы

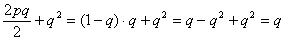
 или по 50 % от отца и матери
или по 50 % от отца и матери
 или по 25 % от каждого деда и каждой бабушки
или по 25 % от каждого деда и каждой бабушки
 или по 12.5 % от каждого прадеда и каждой прабабки
или по 12.5 % от каждого прадеда и каждой прабабки
 или по 6.3 % от каждого прапрадеда и каждой прапрабабки
или по 6.3 % от каждого прапрадеда и каждой прапрабабки
 или по 3.15 % от каждого предка
или по 3.15 % от каждого предка
 или по 1.6 % от каждого предка
или по 1.6 % от каждого предка
 или по 0.8 % от каждого предка
или по 0.8 % от каждого предка
 или по 0.4 % от каждого предка
или по 0.4 % от каждого предка
 или по 0.2 % от каждого предка
или по 0.2 % от каждого предка
 или по 0.1 % от каждого предка
или по 0.1 % от каждого предка
 или по 0.05 % от каждого предка
или по 0.05 % от каждого предка
 , где n – число поколений. Он обнаружил, что в чистых гомозиготных линиях, при постоянном и неизменном генотипе, качество фенотипа зависит от случайных причин, не связанных с генетикой.
, где n – число поколений. Он обнаружил, что в чистых гомозиготных линиях, при постоянном и неизменном генотипе, качество фенотипа зависит от случайных причин, не связанных с генетикой. ,
, 





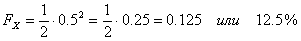


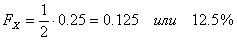
 ,
,