
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Электрофоретическая подвижность
Подвижность белка в изоэлектрической точке равна нулю. При сдвиге рН на две единицы в любую сторону, подвижность белка становится сравнима с подвижностью малых ионов. Она зависит также от ионной силы и температуры, поэтому о подвижности следует говорить, когда хорошо известны условия среды. Основной причиной различий подвижности является белковый заряд. Но нельзя все сводить только к заряду. Согласно теории Дебая-Хюккеля в растворе электролита каждый заряд экранирован диффузной атмосферой противоионов, плотность которой убывает с расстоянием по закону: ρ = ρ 0е-æ r, r – расстояние, æ = J = Подвижность сферических частиц, движущихся при электрофорезе, с точностью до коэффициента определяется формулой Смолуховского: U =
Рассмотрим два случая: 1) низкие ионные силы (разбавленный раствор электролита): J < < 0, 01. В этих растворах толщина дебаевской диффузной оболочки больше размеров частиц. При таких условиях:
Заряд глобулы пропорционален её поверхности: z = 4π a2α e, где α е – заряд единицы поверхности. Тогда электрофоретическая подвижность: U = 2) ионная сила J Толщина ионной атмосферы В этом случае подвижность не зависит от размера частицы, от молекулярной массы белка. Это ближе к реальным условиям электрофореза белков, который при не слишком низких ионных силах буферных растворов определяется зарядом, но не сильно чувствителен к молекулярным массам и размерам белков. Конформационные переходы у пептидов Различить ту или иную структуру белка (α, β или неупорядоченную) сложно. Равновесная конформация в белках обеспечивается взаимодействием боковых групп аминокислотных остатков друг с другом и с растворителем, а с другой стороны - водородными связями между пептидными группами. Роль водородных связей растет в неполярных растворителях и падает в сильнополярных. Сильнополярные растворители сами образуют водородные связи с CO и NH группами пептидных группировок. Если боковые группы заряжены одноименно, то упорядоченность исчезает, а если они не заряжены, то в зависимости от их взаимодействия друг с другом и с растворителем можно наблюдать α, β структуры и стат. клубок. При изменении температуры, рН, состава растворителя обнаруживаются конформационные переходы между этими тремя типами структур. Эти переходы кооперативны и происходят внутри каждой макромолекулы. Полилизин в заряженном состоянии не образует регулярных структур (β –спиралей) и ведет себя как линейный полиэлектролит. При рН > 10, 5 – разряжен и образует α -спираль, которая устойчива в воде при температуре ниже 20 0С. При нагревании до 200 – 350С молекулы полилизина переходят в состояние статического клубка. При температуре выше 350С наблюдается второй конформационный переход и возникают β -складчатые структуры. При отливке пленок из растворов, содержащих белки, макромолекулы белков сохраняют ту конформацию, которую они имели в этом растворителе. Структура и упаковка в твердой фазе этих молекул определяется свойствами растворителя, из которого полимер выделяется при испарении жидкости.
Метод Линдерштрема и Ланга Большинство методов дают суммарное число аминокислотных остатков в α -спиральных и β -структурных участках макромолекулы, а т.к их различить сложно, то используется понятие степени регулярности структуры белка. По скорости замены атомов водорода (Н) в NH-группах пептидной группировки на Н2 или Н3 удобно определять степень регулярности структуры белка. Н1 в NH-группах обменивается на дейтерий быстро, если полипептид имеет низкую молекулярную массу и если он неупорядочен, и наоборот. Скорость обмена зависит от условий эксперимента. Переход от быстрого обмена к медленному – признак образования внутримолекулярных водородных связей. В молекуле РНК-азы из 123 пептидных атомов водорода 70 обмениваются медленно, следовательно, степень регулярности равна 57%. Оказалось, что из этих 70 пептидных групп, стабилизированных водородными связями, 25 способны медленно обмениваться при 00С, еще 25 – при 380С и 20 не обмениваются до 600С. При нагревании возле 600С происходит разрыв водородных связей между пептидными группировками, α -спираль превращается в стат. клубок (денатурация).
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 548; Нарушение авторского права страницы
 =1/æ – средняя толщина ионной атмосферы.
=1/æ – средняя толщина ионной атмосферы.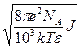 ,
, 
 – ионная сила, е – заряд электрона,
– ионная сила, е – заряд электрона,  − число Авогадро,
− число Авогадро,  − диэлектрическая проницаемость.
− диэлектрическая проницаемость. ,
,  - это вязкость среды,
- это вязкость среды,  - разность потенциала в двойном слое, окружающем частицу.
- разность потенциала в двойном слое, окружающем частицу. , где z - заряд белковой частицы,
, где z - заряд белковой частицы,  – ее радиус.
– ее радиус.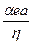 .
. 0, 1.
0, 1. 1/æ. В этих условиях:
1/æ. В этих условиях:  æ -1
æ -1  U =
U =  æ -1.
æ -1.