
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Упругость полимерной цепи с исключенным объемом
Модуль упругости идеальной цепочки, растягиваемой за концы силой f: Упругость обусловлена тепловым движением, т.е. флуктуациями конформаций, а флуктуации в клубке имеют макроскопический характер. Отсюда следует, что упругое поведение клубка должно зависеть от его общего размера R~l•Ng (для идеальной цепи g=1/2, для цепи с исключенным объемом g=3/5), но не от каких-либо микроскопических параметров звеньев. Поскольку для реальных цепей с исключенным объемом g=3/5, видно, что h~N в степени больше единицы: h~Nr, (r> 1) Этот результат имеет простой физический смысл: в реальной цепи связи воспринимают не только внешнюю нагрузку, как в идеальной цепи, но еще и расталкивающее действие межзвенных контактов. Для реальной цепи g=3/5 получается нелинейная зависимость растяжения от силы h~f2/3, т.е. мы видим, что полимерный клубок с исключенным объемом ведет себя под действием внешней нагрузки совсем не так, как идеальная цепь. Анализируя эти рассуждения, можно отметить, что главным явилось предположение о том, что из всех характеристик клубка упругие свойства зависят лишь от его общего размера lNg, т.е. что этот размер является единственным характерным масштабом клубка. В этом предположении заключена так называемая гипотеза масштабной инвариантности, или скейлинга (scale – масштаб). Теории, основанные на этой гипотезе, называются скейлинговыми. Самым важным в скейлинговых теориях является учет флуктуаций плотности звеньев.
Осмотическое давление полимерного раствора Макромолекулы создают избыток давления со своей стороны от мелкопористой перегородки (давление растворителя с обеих сторон одинаково). Этот избыток и есть осмотическое давление. В разбавленном растворе (с< < c*) не перекрывающихся клубков (с – концентрация полимера в растворе, с* - концентрация полимера в клубке) осмотическое давление – это избыток давления за счет давления клубков на перегородку. Поскольку число клубков в единице объема пропорционально с/N, осмотическое давление: Когда концентрация полимерного раствора достигает с*, клубки начинают перепутываться и в полуразбавленном растворе при c> > c*, отталкивание звеньев дает больший вклад в давление, чем поступательное броуновское движение целых клубков. По теории Флори для осмотического давления получили бы p~kT•Bc2 (В – второй вириальный коэффициент, B=a3, a – размер звена). Согласно скейлинговым рассуждениям клубки имеют единственный характерный макроскопический размер – размер R клубка как целого. Тем самым есть лишь одна характерная концентрация N/R3~c* и поэтому осмотическое давление:
где f – функция, которую и требуется определить. В полуразбавленном растворе, т.е. при с/с*> > 1 было установлено: f(с/с*)~(с/с*)5/4, т.е. Иными словами, был получен существенно отличающийся от теории Флори результат. Причина расхождения заключается в том, что в теории Флори флуктуации концентрации звеньев никак не учитываются. Разбавленные и неразбавленные растворы полимерных клубков сильно флуктуируют, и пренебрежение флуктуациями для этих систем приводит к значительным ошибкам. Скейлинговые рассуждения есть, по сути дела, простой способ получить результаты точной флуктуационной теории. Эксперименты полностью подтвердили скейлинговую формулу p»с9/4 (а не p~kTBc2~kTa3c2, а – длина звена). С ростом концентрации различие этих результатов уменьшается. Это указывает на то, что по мере концентрирования роль флуктуаций уменьшается, в концентрированном растворе они вообще несущественны. Статистика линейных полимеров Степень полимеризации Проводя статистический анализ полимерных молекул, приходится определять, сколько содержится в образце молекул каждого сорта, или каково среднее число мономерных звеньев в полимерной молекуле, какова вероятность общей заданной конформации, каково среднее расстояние от одного конца молекулы до другого и тому подобные задачи. Этому и посвящен данный раздел. С точки зрения биофизики (и молекулярной биологии) ответы на эти вопросы будут во многом способствовать выяснению физико-химических свойств биомакромолекул и, в конечном итоге, особенностей их функционирования. Степенью полимеризации полимерной молекулы называется число структурных субъединиц, содержащихся в молекуле – х. Молекула со степенью полимеризации х называется х-мером. Определение годится и для разветвленных полимеров. В простом линейном полимере идентичны все звенья, кроме концевых. Молекулярная масса Мх х-мера является суммой молекулярных масс всех х звеньев. При больших х (х> 100) концевыми эффектами можно пренебречь и считать, что все звенья имеют молекулярную массу Мо, тогда:
Мх = хМо,
где Мо – константа для полимера данного типа (не зависит от х), поэтому термины молекулярная масса и степень полимеризации в какой-то степени идентичны.
Распределение по молекулярным массам Степень полимеризации полимеров даже одного типа (как искусственных, так и природных) может колебаться в широких пределах. Распределение по молекулярным массам описывает частоту нахождения данной степени полимеризации. Наиболее обычным является задание изменения мольной доли х-мера (Хх) как функции х, т.е. Хх = f(х) Подобное уравнение называется функцией распределения. Хх – доля от общего числа молекул, приходящихся на х-мер. Например, функция распределения для поликонденсации, при которой не изменяется реакционная способность функциональных групп в зависимости от х: OH-R-COOH Пусть в начальном состоянии Nо мономеров, т.е. по No функциональных групп каждого типа. Пусть полимеризация протекает до тех пор, пока не прореагирует доля р от всех функциональных групп. Число оставшихся функциональных групп каждого типа равно тогда No(1-p). Но это же число определяет и число независимых молекул, так как каждая молекула имеет концевые не прореагировавшие группы. Среднечисловая степень полимеризации
Для того, чтобы Предположим, что все молекулы в конечной смеси расположены друг за другом вдоль конечного отрезка. Пусть он состоит из No звеньев структуры. В р•No местах звенья будут соединены СО-О-связями, в остальных (1-р)No местах связей нет. В нашем конечном отрезке будут точки, где кончается одна молекула и начинается другая. Если реакционная способность функциональных групп не зависит от длины цепи, размещение этих точек будет случайным. Вычислим вероятность того, что данная структурная единица (звено) является частью х-мера. Чтобы вычислить эту вероятность, нужно сначала определить вероятность того, что выбранное звено является начальной частью х-мера. Эта вероятность равна вероятности того, что вдоль нашего отрезка расположатся (х+1) состояний в последовательности:
1 – несвязанное состояние; 2 – (х-1) СО-О-связей; 3 – следующее несвязанное состояние. Остальные состояния могут быть либо связями, либо промежутками.
Так как эти состояния являются независимыми, вероятность того, что мы имеем в данном месте начинающийся х-мер, является произведением вероятностей нужного нам события для каждого состояния. Эта вероятность равна р для имеющих связи и (1-р) для не имеющих связи состояний (так как доля связанных функциональных групп – это и есть мера вероятности того, что в данном состоянии есть связь). Таким образом, вероятность того, что выбранное звено является начальной частью х-мера, равна Для всех других, кроме интересующих нас (х+1) состояний вероятность равна 1, так как не имеет значения, связаны они или нет. (х+1) здесь, так как одно несвязанное состояние плюс (х-1)-связей плюс второе несвязанное состояние с другого конца х-мера. Но нам надо определить вероятность того, что данное звено является частью x-мера. Если это звено должно быть частью любого х-мера, то вдоль цепи имеется х состояний, с которых можно начать отсчет. Таким образом, полная вероятность того, что выбранное звено является частью х-мера, равна Вероятность того, что данная структурная единица (звено) входит в х-мер, эквивалентна доле структурных единиц, составляющих х-меры. Считая, что все звенья имеют одинаковую массу, эта вероятность эквивалентна также весовой доле х-мера Wx:
Это и есть распределение по массам (называют чаще весовым распределением). Оно показывает долю от общей массы, приходящуюся на х-мер. Определим теперь числовое распределение, т.е. долю от общего числа молекул, приходящуюся на х-мер. Общая масса равна NoMo/NA, где Мо – молекулярная масса структурной единицы –СО-R-О-, NA – число Авогадро. Масса всех молекул х-мера равна WxNoMo/NA. Масса каждой х-мерной молекулы равна хMo/NA и число х-мерных молекул равно WxNo/х. Мы отметили, что общее число всех молекул равно No(1-p), поэтому мольная доля х-мера может быть записана так:
Это и есть числовое распределение, т.е. доля от общего числа молекул, приходящаяся на х-мер (или мольная доля). Комбинируя полученные выражения для распределений с выражением для Сумма всех весовых (Wx) долей и всех мольных (Xx) долей равна единице.
Функции на этих рисунках не непрерывны (х – целочисленные), но при больших х можно эти разрывные функции заменить непрерывной функцией распределения:
где Приведенные распределения – это частный случай распределений, которые могут быть получены при синтезе полимеров. Тип распределения зависит от кинетики полимеризации.
Фракционирование полимеров Фракционирование гетерогенных полимеров производят: - используя их различия в растворимости, связанные с разными длинами цепи; - с помощью ультрафильтрования через градуированные мембраны; - с помощью хроматографии на колонках; - с помощью ультрацентрифугирования. В результате получают более гомогенные растворы полимеров, с более узким распределением.
Средние молекулярные массы Закономерности, по которым данные физические свойства зависят от молекулярной массы, также определяют способ усреднения, который должен быть применен в каждом случае. Таким образом, при рассмотрении различных физических свойств мы неизбежно должны применить различные средние молекулярные массы. Особенно важны среднечисловая и средневесовая, встречаются также средневискозиметрическая и «Z-средняя» молекулярные массы. Среднечисловая
Вместо числа молекул Ni мы можем использовать число молей ni, число молекул или молей в единице объема или мольную долю. Удобно пользоваться мольными долями Если смесь представляет собой чистый полимерный образец, в котором молекулы различаются только по степени полимеризации, то х для различных сортов молекул будет меняться от 1 до ¥ и их молекулярные массы будут равны Мх=хМо так, что:
Среднечисловая степень полимеризации определяется также:
где Nx – число х-мерных молекул или их количество в единице объема. Средневесовая молекулярная масса.
И если полимер состоит из одинаковых звеньев, то:
Средневесовая степень полимеризации:
где Wx – весовая доля. Общее определение для Так как масса i-го сорта молекул gi, очевидно, равна NiMi/NA, то:
Используя выражения для Wx и Хх, полученные нами ранее для:
и
Z-средний - Z+1-средний - Суммирование по всем возможностям i. Если образец полностью гомогенен, то во всех случаях
Разность вязкости полимерного раствора и соответствующего растворителя часто может быть пропорциональна произведению весовой концентрации и Мa, где a - константа (0, 5-2, 0=a). Для гетерогенного образца получают средневискозиметрический молекулярный вес
Глава 3. Физика белка. 3.1. Общая характеристика белков Белки - это сложные макромолекулярные структуры, нерегулярные полимеры, построенные из остатков аминокислот. В общем виде, в водном растворе при нейтральном рН, аминокислоты имеют вид типа:
Их структура и структура надмолекулярных комплексов определяется ковалентными химическими связями и слабыми взаимодействиями. Нативные белки никогда не являются статистическими клубками. Физики их считают апериодическими кристаллами сложной структуры. Их поведение зависит от положения и свойств всех элементов молекул, т.е. существенно зависит от той конформации, в которой находится белковая молекула. В организме человека ~105 видов белков. Это странно, т.к. число генов приблизительно несколько десятков тысяч (один белок кодируется одним геном). Изучая химический состав белков, мы узнаем, из каких химических элементов они состоят. Анализ показал: во всех белках примерно 50-55% углерода, 21-23% кислорода, 15-17% азота, 6-7% водорода, 0.3-2.5% серы. В составе больших белков обнаружены: фосфор, йод, железо и др. макро- и микроэлементы в разных количествах. Содержание основных химических элементов белка может различаться за исключением азота, т.к. концентрация азота постоянна и равна примерно 16%. Содержание азота в других органических веществах мало, поэтому предложили определять количество белка по входящему в его состав азоту. Зная, что 1г азота содержится в 6.25г белка, найденное количество азота умножают на 6.25. Для определения химической природы белка, его разделяют на мономеры и выясняют их химический состав. Расщепление белка на составные части производят с помощью гидролиза (кислотного или щелочного). Кислотный гидролиз – это длительное кипячение белка с сильными кислотами. Чаще всего кипятят белок с HCL при 100º С в течение 24 часов. 2-ой этап: хроматография – это разделение веществ, входящих в состав гидролизата. Аминокислотный состав определяют с помощью анализа радикалов (гораздо сложнее определить их последовательность). Функционирование белка связано с определенной конформацией. Нарушение в одном звене молекулы белка обязательно приведет к изменению её конформации. Денатурация – это необратимое разрушение белка. Ренатурация - это восстановление белка. Задачи физики белков. 1) изучение структуры белка и надмолекулярных комплексов. 2) исследование динамического поведения белков (в том числе изучение денатурации). 3) установление связи между первичной структурой и структурами белков более высокого порядка. 4) изучение взаимодействия белков с нуклеиновыми кислотами. 5) изучение физики возникновения и эволюции белков. 6) изучение физических аспектов белковой изомерии. 7) изучение методов получения белков с заданными свойствами 8) изучение физических механизмов, лежащих в основе различных биологических функций.
Функции белков Белки выполняют следующие функции: 1) транспортная: транспортировка и запасание кислорода, жиров, железа, меди и др. элементов; 2) гормональная: белки-гормоны регулируют обмен веществ. (Например, белки гипофиза, поджелудочной железы); 3) каталитическая: белки являются ферментами; 4) защитная (иммунная): белки являются антителами иммунных процессов – в гуморальном иммунитете участвуют γ -глобулины, в клеточном иммунитете участвуют лимфоциты, выполняющие свои функции благодаря белкам, расположенным на лимфоцитарных мембранах. 5) белки в составе крови способствуют ее свертываемости. 6) структурная (опорная): белки являются компонентами соединительной ткани (коллаген), кожи, волос (кератин), сосудов (эластин), хрящей. Вместе с углеводами белки входят в состав секретов (гликопротеиды). Они являются компонентами слизистых оболочек, связок, сухожилий. 7) сократительная (механохимическая): белки преобразуют энергию АТФ в механическую энергию. 8) сигнальная: обеспечение рецепции и передачи химических сигналов, участвуют в распространении нервных импульсов. 9) регуляторная (белки осуществляют обратную связь в метаболических цепях). 10) белки входят в состав рецепторных систем организма, перекодируя внешние сигналы в электрохимический импульс. 11) энергетическая (питательная): белки являются источниками питания для развития плода (казеин). 12) Многие белки являются токсинами, защищающими организм от вредных воздействий. 13) Белки поддерживают рН (буферы) и сохраняют онкотическое давление в клетках крови. Существует множество иных специальных функций белков. Гормональная функция. Каждый белок выполняет одну узкоспециальную функцию и только в некоторых случаях выполняет несколько взаимосвязанных функций. Гормон мозгового слоя надпочечников – адреналин поступает в кровь, повышая потребление кислорода, повышает артериальное давление; с другой стороны, он повышает уровень содержания сахара в крови и стимулирует обмен веществ. У холоднокровных животных адреналин – это медиатор нервной системы. Каталитическая функция. Биохимические реакции в организме зачастую идут в мягких условиях (при температуре примерно 40º С и рН, стремящемся к 7.0). Но в этих условиях скорости протекания большинства биохимических реакций малы, следовательно, чтобы в организме они имели место, эволюционно возникли биологические катализаторы – ферменты. Ферменты – это либо белки, либо комплексы белков с каким-либо ко-фактором (ион металла или какая-нибудь органическая молекула). Ферменты обладают высокой избирательностью действия, в основе которой лежит определенная конформация белковой молекулы. Любое отклонение будет приводить к уменьшению избирательности и, следовательно, к ухудшению катализа. Транспортная функция. Внутрь клетки должны поступать многочисленные вещества (и строительный материал, и энергия), а биомембраны построены по одному принципу: двойной слой липидов, в который погружены различные белки, при этом гидрофильные участки макромолекул располагаются на поверхности мембраны, а гидрофобный хвост белка находится в толще мембраны. Такая структура мембран непроницаема ни для крупных молекул (аминокислоты, сахара), ни для относительно малых (ионы металлов). Их проникновение идет с помощью транспортных белков. У многоклеточных организмов существует и другая система транспорта веществ. Это гемоглобин крови или сывороточный альбумин (обладающий способностью образовывать прочный комплекс с жирными кислотами, с гидрофобными аминокислотами (триптофан), со стероидными гормонами, с лекарственными препаратами (аспирин, пенициллин) и переносить их в различные органы), трансферрин обеспечивает перенос ионов железа, церуплазмин – ионы меди. Рецепторная функция. Для функционирования организма большое значение имеют белки-рецепторы, вмонтированные в плазматическую мембрану клеток, которые служат для восприятия и преобразования сигналов, поступающих в клетку (как от окружающей среды, так и от соседних клеток). Например, рецепторы ацетилхолина находятся в межнейронных контактах на мембране клеток, в том числе в коре головного мозга. Эти белки специфически взаимодействуют с ацетилхолином. Это взаимодействие приводит к такому изменению конформации белка, в результате которого становится возможной передача сигнала внутрь клетки. После получения и преобразования сигнала, нейромедиатор (ацетилхолин) удаляется, и клетка готовится к восприятию следующего сигнала. Чтобы он ушел, существует специальный фермент – ацетилхолин эстераза, катализирующий гидролиз ацетилхолина до ацетата и холина. Многие гормоны не проникают внутрь клетки-мишени, а связываются со специфическими рецепторами на поверхности этих клеток. Это связывание уже является сигналом для запуска в клетке физиологического процесса. Защитная функция. Иммунная система обладает способностью отвечать на появление в организме чужеродных частиц выработкой лимфоцитов, и с другой стороны, отвечать производством белков (иммуноглобулинов). В первом случае говорим о клеточном иммунитете, во втором – о гуморальном. В ходе клеточного иммунитета лимфоциты разрушают чужеродные клетки, патогенные бактерии, раковые клетки, вирусы; во втором случае В-лимфоциты вырабатывают иммуноглобулины или антитела, которые, связываясь с чужеродными молекулами (антигенами), связывают и снимают эффект их действия. Если в роли антигена выступает большая молекула, то антитело опознает не всю молекулу, а ее определенный участок и этот участок называется антигенной детерминантой. Т.к. иммуноглобулины взаимодействуют с небольшой частью антигена, это позволяет лимфоцитам производить антитела, которые узнают специфические небольшие молекулы даже синтетической природы. Согласно современным представлениям каждый тип иммуноглобулина, вырабатываемый В-лимфоцитами, произошедшими от одного общего предшественника, называется клоном. Структурная функция. Наряду с белками, выполняющими специализированные функции, существуют белки, которые выполняют структурные функции (прочность, эластичность и др. свойства отдельных тканей организма: коллаген – основной белковый компонент соединительной ткани (у млекопитающих он составляет до 25% массы белков)). Он синтезируется в фибробластах. Первоначально, он возникает в виде проколлагена, который проходит определенную обработку (окисление остатков пролина и лизина до гидроксипролина и гидроксилизина) в фибробластах. Коллаген – это три скрученные в спираль полипептидные цепи. Выходя из фибробластов, эти спирали объединяются в коллагеновые фибриллы (волокна) и образуют коллагеновые нити, которые можно наблюдать под микроскопом. Двигательные белки. Мышечные сокращения – это процесс, при котором происходит преобразование химической энергии, запасенной в пирофосфатных связях молекул АТФ, в механическую работу. Участвуют два белка актин и миозин. Миозин состоит из длинной нитевидной части (хвоста) и двух глобулярных головок. Общая длина одной молекулы порядка 1600 нм, на головки приходится ~200 нм. Выделяют миозин в виде гексомера, образованного двумя цепями с молекулярной массой 200 тыс. и четырьмя легкими цепями с молекулярной массой около 20 тыс. Тяжелые цепи закручены спирально одна вокруг другой и образуют хвост. На одном конце нити находятся глобулярные головки, ассоциированные с легкими цепями. На головках миозина находятся два функциональных центра: 1-ый – каталитический (осуществляет расщепление пирофасфатной связи АТФ), 2-ой центр обеспечивает миозину способность специфически связываться с другим мышечным белком – актином. Актин – это глобулярный белок, с молекулярной массой 42 тыс (G-актин в глобулярном виде). G-актин обладает способностью полимеризоваться с образованием длинной структуры F-актина. В такой форме он может взаимодействовать с головкой миозина, и это взаимодействие сильно зависит от наличия АТФ. При высокой концентрации АТФ комплекс актина и миозина разрушается. После того, как под действием миозиновой атефазы происходит гидролиз АТФ, комплекс снова восстанавливается. Этот процесс часто наблюдают в растворе, содержащем оба белка. Без АТФ − это вязкое, высокомолекулярное соединение, добавление АТФ сразу уменьшает вязкость, а по мере разрушения АТФ опять происходит комплексообразование (конденсация). Эти превращения важны при мышечных сокращениях. Аминокислоты Аминокислоты – это органические кислоты, у которых атом водорода у α -углеродного атома замещен на аминогруппу:
Все аминокислоты, кроме глицина, имеют хиральный атом Cα и существуют в виде двух энантиомеров (2-х оптических изомеров). В настоящее время в состав всех изученных белков входят только L-аминокислоты. Биологический смысл этого заключён в том, что если это была бы смесь изомеров, то было бы бесконечно много диастереоизомеров.
Структура аминокислот в L и D формах.
D-аминокислоты широко распространены в живой природе. Они входят в состав олигопептидов. Из 20 основных α -аминокислот строятся белки. Некоторые аминокислоты образуются из этих 20-ти аминокислотных остатков, но в составе белковой молекулы. Например, при окислении двух сблизившихся остатков цистеина за счёт возникновения связи − S− S− образуется диаминодикарбоновая кислота – цистин (R=SH). Сшивка S–S может возникать как между различными полипептидными цепями, так и внутри одной цепи. Фосфорилирование остатков серина:
Такие ферменты как трипсин, химотрипсин, холинэстераза активны благодаря ОН группе, которая становится неактивной после фосфорилирования. ОН группа находится в активном центре фермента. Аминокислоты в водном растворе существуют в ионизированном состоянии за счет диссоциации амино- и карбоксильных групп. Они могут существовать как кислоты (доноры) или как основания (акцепторы протонов). Природные аминокислоты делят на 3 группы: 1) алифатические (ациклические соединения жирного ряда); 2) циклические; 3) иминокислотный остаток− пролин. Ациклические: 1) аминокислотные остатки с углеводородными хвостами (аланин, валин, глицин, лейцин, изолейцин); 2) аминокислотные остатки с гидроксильной группой (серин, треонин); 3) аминокислотные остатки с кислотным остатком (аспарагиновая и глутаминовая кислоты); 4) аминокислотные остатки с группой СОNH2 (аспарагин и глутамин); 5) аминокислотные остатки с аминогруппой (аргинин и лизин) (т.е. R содержит дополнительную аминогруппу); 6) аминокислотные остатки, содержащие серу (метионин, цистеин). Глицин участвует в образовании нуклеиновых и желчных кислот, в образовании гема, необходим для обезвреживания в печени токсичных продуктов. Аланин используется организмом в различных процессах обмена углеводов. Серин входит в состав различных ферментов, липопротеинов. Заменимая аминокислота. Треонин – незаменимая аминокислота, участвует в биосинтезе белка. Цистеин – заменимая аминокислота, которая легко окисляется, защищая организм от веществ с высокой окислительной способностью. Метионин – незаменимая аминокислота. Характеризуется высокой подвижностью. Она содержит СН3 группу и участвует в синтезе холина, кератина, адреналина. Валин, лейцин и изолейцин активно участвуют в обмен веществ, в организме не синтезируются. Аспарагин и глутамин входят в состав тормозных медиаторов нервной систем. Циклические: Аминокислотные остатки, содержащие π -электронные циклы, − гистидин, тирозин, триптофан, фенилаланин. Гистидин:
Незаменимая аминокислота, входит в состав большинства белков. Тирозин:
Заменимая аминокислота (в организме человека образуется при окислении фенилаланина). Это активная часть гормона щитовидной железы. Триптофан:
Незаменимая аминокислота (суточная потребность для человека − 0, 25г). Много триптофана в γ -глобулине, в белке фибриногене (основном компоненте системы свёртывания крови). Фенилаланин:
Незаменимая аминокислота (суточная потребность для человека − 1, 1г). Входит в состав почти всех белков (яичный белок, гемоглобин и т.д.). Иминокислота − пролин:
Заменимая кислота (в организме человека синтезируется из глутаминовой кислоты). Входит в состав почти всех белков. При её биохимическом окислении образуется оксипролин. Помимо основных аминокислотных остатков встречаются несколько производных уже названных аминокислот: оксилизин, оксипролин, и др. Найдены пиролизин, фосфосерин, тироксин и др. редкие аминокислоты.
Первичная структура Цепочка аминокислотных остатков в белковой молекуле возникает при образовании пептидной связи:
Белки имеют молекулярную массу от 6000 до нескольких миллионов. Для того чтобы установить первичную структуру (последовательность аминокислотных остатков), надо определить количество цепей, разделить их и определить аминокислоты в цепях. Число цепей можно узнать по количеству N и C концов. Для определения N-концевой аминокислоты предложен ряд методов, в частности метод Сенгера: 1) реакции амино- и оксигрупп с 2, 4-динитрофторбензолом; 2) полный гидролиз; 3) хроматография для определения аминокислоты. Для определения природы С-концевой аминокислоты чаще всего пользуются ферментативным путем. Все четыре атома пептидной группировки лежат в одной плоскости. Это следует из рентгеноструктурного анализа. Было показано, что пептидная связь вследствие сопряжения С=О и N-атома, на р-орбитали которого находится неподеленная пара электронов, не может рассматриваться как ординарная и вращение практически отсутствует. По этой же причине хиральный атом Сα и карбонильный атом группировки С=О и N-Cα находятся в одной плоскости, но в этой же плоскости и О, и Н. Следовательно, все атомы находятся в одной плоскости. Геометрически полипептидную цепочку можно рассматривать как образованную плоскими фрагментами структуру (по шесть атомов в одной плоскости). Возникают торсионные углы: угол ψ между Сα и С и угол φ между N и Сα.. Выражение для потенциальной энергии внутреннего вращения выглядит следующим образом:
U(ψ, φ )=U0/2*(1-cos3φ )+U0/2*(1-cos3ψ )+ Первичная структура определена генетически; она определена составом и порядком следования аминокислот. Реакция поликонденсации сопровождается выделением воды.
Вторичная структура Фрагменты пространственной структуры любого биополимера, имеющие периодическое строение рассматриваются как элементы вторичной структуры. Под вторичной структурой белка подразумевают конфигурацию полипептидной цепи, т.е. способ свертывания, скручивания полипептидной цепи в спиральную или какую-либо другую конформацию. Различают две основные конфигурации: α -спирали и β -структуры. Спиральная структура белка (α -спираль). |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 478; Нарушение авторского права страницы
 и
и  ~
~  (
(  ).
). , N> > 1 (число звеньев).
, N> > 1 (число звеньев). ,
,  .
. H-[-O-R-CO-]x-OH + H2O
H-[-O-R-CO-]x-OH + H2O определяется как среднее число мономерных звеньев в молекуле и равна:
определяется как среднее число мономерных звеньев в молекуле и равна: 
 или
или  (х-1, так как х звеньев связаны х-1 связью).
(х-1, так как х звеньев связаны х-1 связью). . Так как молекула может состоять из одного звена, то эта же вероятность может быть и вероятностью того, что этот х-мер является долей от общего числа х-меров.
. Так как молекула может состоять из одного звена, то эта же вероятность может быть и вероятностью того, что этот х-мер является долей от общего числа х-меров. (надо учитывать выделение воды при поликонденсации).
(надо учитывать выделение воды при поликонденсации).

 ;
; 
 ;
;  ,
,  - бесконечно малая доля веса полимера, имеющего степень полимеризации между х и х+dх. Преимущество функций такого рода в том, что х можно интегрировать в конечных пределах.
- бесконечно малая доля веса полимера, имеющего степень полимеризации между х и х+dх. Преимущество функций такого рода в том, что х можно интегрировать в конечных пределах. получается путем суммирования количества молекул каждого сорта, умноженных на их молекулярную массу, и деления на общее число молекул. Для любой смеси молекул:
получается путем суммирования количества молекул каждого сорта, умноженных на их молекулярную массу, и деления на общее число молекул. Для любой смеси молекул:  , где Ni – число молекул сорта i, имеющихся в смеси, Mi – их молекулярная масса.
, где Ni – число молекул сорта i, имеющихся в смеси, Mi – их молекулярная масса. и
и  .
.
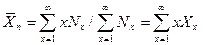 ,
,  получают путем сложения числа граммов gi полимера, молекулы которого имеют молекулярную массу Mi, умноженного на эту молекулярную массу, и деления на общую массу полимера в граммах. Можно использовать также концентрацию г/мл или весовую долю Wi:
получают путем сложения числа граммов gi полимера, молекулы которого имеют молекулярную массу Mi, умноженного на эту молекулярную массу, и деления на общую массу полимера в граммах. Можно использовать также концентрацию г/мл или весовую долю Wi: 
 ,
, 
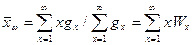 ,
, 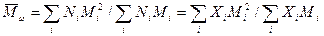
 ,
, 
 . При р®1
. При р®1 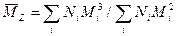

 =
= 
 .
.
 =
= 










 +Uкул
+Uкул