|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Денатурация глобулярных белков
Денатурацию глобулярных белков вызывают: нагревание, воздействие ультразвуком, радиационное воздействие, рН, состав растворителя. Экспериментально было показано, что денатурацию вызывают слабополярные органические растворители. Их действие связано с контактами с гидрофобными группами белка. Было обнаружено, что денатурирующее действие спиртов на белки возрастает с увеличением размера алифатического радикала. Глобула имеет фиксированную пространственную структуру и переход “глобула клубок” отличен от перехода “спираль - клубок” потому, что он происходит в трехмерной системе. С другой стороны, кооперативность перехода (которая определяет величину изменения температуры (интервал Δ T, в котором происходит переход), а Δ T примерно одинаково при плавлении α -спирали и глобулярного белка) связана не только с взаимодействием ближнего порядка, но и дальнего. Показательны денатурационные превращения в зависимости от рН. При уменьшении рН уменьшается изменение энтальпии и температура перехода. Например, для химотрипсиногена:
Конформационные переходы белковых молекул связаны с разворачиванием полипептидной цепи и часто происходят малые, локальные и трудно наблюдаемые изменения, которые не сопровождаются сильными изменениями теплоемкости. Когда изменения теплоемкости при денатурации нативных белков велики, их связывают с нарушением гидрофобных взаимодействий. По существенным изменениям теплоемкости можно судить о характере перехода. Т.к. при денатурации глобулярных белков изменяется структура молекулы, то исследования могут способствовать установлению факторов, стабилизирующих белковую глобулу. Факторы: 1) В ходе денатурации белки характеризуются большей поворотной гибкостью цепи, и поэтому среднее изменение энтропии Δ 2) Изменение энтальпии Δ 3) Изменение свободной энергии Δ 4) Реакции денатурации белка сильно зависят от рН. Факторы, определяющие эту зависимость, делят на две группы: а) дальнодействие электростатического взаимодействия; б) аномальные группы и эффекты, связанные с титрованием. Многие белки содержат ионизированные группы, рК которых отличаются на несколько единиц от нормального значения рК для этих групп в развёрнутом состоянии. В ходе денатурации происходит нормализация рК этих групп, т.е. изменяется связывание протонов с этими группами. Это приводит к появлению титрационной составляющей изменения свободной энергии (Δ Fтитр.). Δ Fg = p(N р – доля пептидной цепи, развернутой в ходе денатурации, или степень разворачивания. N – число звеньев в молекуле.
3.31. Метод Тенфорда определения разности свободной энергии денатурированного и нативного белка по денатурации в растворе мочевины Для изменения свободной энергии денатурации белка в растворе мочевины можно записать: − Δ Fu = RTlnKu = RTln δ (Δ F) = Δ Fu - Δ FH δ (Δ F) = ni – число групп (звеньев) типа i в белке. α i – численный параметр, зависящий от степени доступности растворителю групп данного типа в нативной конформации. Δ Fi – вклад групп типа i в свободную энергию перехода. α i (пептидн.) α i (полярн.) α i (гидроф.) Δ Fi определяется из данных о растворимости аминокислот в воде и в растворе мочевины данной концентрации. Для концентрации мочевины (C Δ FH
3.32. Калориметрические измерения денатурационных изменений в белках Денатурацию рассматривают как мономолекулярную реакцию перехода из нативного состояния белка (N) в денатурированное (Д): N← → Д В калориметрии измеряют Δ Н или Δ Ср, сопровождающие эти переходы. В соответствии с формулой Киргоффа изменение энтальпии при плавлении: Δ Н = В обычных измерениях подводят теплоту к образцу: Чувствительность микрокалориметров высокая, примерно 10-7 Вт.
Δ Сpd – это разность теплоемкостей белка в нативном и денатурированном состояниях при температуре перехода Td. Δ Нден = где Δ Нd − молярная энтальпия перехода. Для энтропии: Δ Sден = Свободная энергия Гиббса для процесса денатурации (Δ Fден ) запишется в виде: Δ Fден = Δ Нден - T1 Δ Sден = Δ Нd
МИОГЛОБИН Миоглобин - сложный белок третьего уровня структурной организации. Состоит из одной полипептидной цепи (153 остатка). Третичная структура белка образована, главным образом, альфа-спиралями вторичной структуры, на которые приходится около 70 процентов АКО, остальные - на повороты и начальный и конечный участки. Белок содержит в себе так называемый ГЕМ- комплекс порфирина и иона железа в степени окисления +2.
Ион железа встроен в кольцо порфирина таким образом, что четыре координационные связи из шести затрачены на образование связей с атомами азота, еще одна связана с азотом имидазольного остатка ГИС полипептидной цепи (проксимальный Гистидин F8), а другая - также с имидазольным остатком другого ГИС (дистальный ГИС Е7). Молекула кислорода присоединяется между остатком дистального ГИС и железом. Изменения степени окисления железа при этом не происходит. Порфириновое кольцо (ГЕМ) частично погружено в молекулу белка. Молекула кислорода присоединяется к нему, входя как бы через открывающуюся дверцу. Пока остается неясным, дожидается молекула кислорода случайного открывания двери, или существует какой-то механизм, пропускающий кислород к нему.
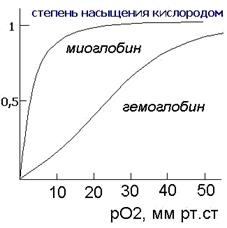
Миоглобин сосредоточен, главным образом, в мышцах и его главной функцией является хранение кислорода. Скорость насыщения миоглобина кислородом намного превышает таковую для гемоглобина. Миоглобин мало приспособлен для транспортировки кислорода из легких в ткани, поскольку скорость отдачи кислорода в тканях невелика (при давлении 1 мм рт. ст. примерно половина миоглобина все еще не отдает кислород). Вопросы транспортировки кислорода решаются при участии гемоглобина.
ГЕМОГЛОБИН Гемоглобин представляет собой белок четвертичной структуры, состоящий из двух пар субъединиц альфа- (141 АКО) и бета- (147 АКО- аминокислотных остатков). Субъединицы миоглобина и гемоглобина очень сходны между собой, как весьма сходна и третичная структура обоих белков. Главное отличие гемоглобина от миоглобина заключается в проявлении особого рода эффектов - кооперативных, влияющих на скорости присоединения- отсоединения молекул кислорода. Каждая молекула гемоглобина способна присоединять и переносить четыре молекулы кислорода, при этом кооперативность проявляется в том, что как присоединение, так и отсоединение каждой последующей молекулы кислорода облегчается в результате структурных изменений в конформации молекулы, которых у гемоглобина имеется две - оксигенированная и дезоксигенированная. Промежуточные состояния нестабильны. Предполагается следующий механизм кооперативного эффекта. Присоединение первой молекулы кислорода приводит тому, что атом железа смещается от своего места примерно на 0, 4-0, 6 ангстрем, вызывая изменения конформации субъединицы. Изменившаяся конформация по аллостерическому эффекту облегчает присоединение кислорода к другой субъединице и т.д. Это позволяет максимально ускорить процесс присоединения кислорода в легких (РО2 = 100 мм рт. ст.). При переносе оксигенированного гемоглобина в капилляры тканей (РО2 = 5 мм рт. ст.) отсоединение молекул кислорода протекает также быстро, по кооперативному эффекту. Известны, впрочем, и химические регуляторы скорости и полноты присоединения кислорода. К ним, в частности, относится 2, 3- дифосфоглицериновая кислота. Она облегчает присоединение кислорода у организмов, обитающих в высокогорных районах.
Транспорт газов
Большинство тканей для поддержания своего окислительного потенциала постоянно снабжаются молекулярным кислородом (О2). Из-за плохой растворимости О2 связывается и транспортируется в крови гемоглобином. Свойства Нb таковы, что он не только обеспечивает транспорт кислорода, но и создает благоприятные условия для связывания О2 в легких и передачи его тканям. Если фермент реагирует на эффекторы (субстрат, активатор или ингибитор) повышением или понижением активности благодаря конформационным изменениям, то в этом случае говорят об аллостерической регуляции. Аллостерические ферменты являются, как правило, олигомерами, состоящими из нескольких субъединиц, которые взаимно влияют друг на друга. Гемоглобинне является ферментом и имеет все признаки аллостерического белка. Он связывает и отдает кислород неизмененным. Его эффектором служит кислород, который как положительный гомотропный эффектор с повышением концентрации увеличивает константу связывания. Кривая насыщения гемоглобина О2имеет ярко выраженный сигмоидальный характер (2, кривая 2). Для сравнения приведена не сигмоидальная кривая насыщения мышечного белка миоглобина (2, кривая 1). Миоглобинпохож по структуре на гемоглобины, но, являясь мономером, не обнаруживает аллостерических свойств. Гемоглобин участвует также в транспорте диоксида углерода (СО2) от тканей к легким. Примерно 5% образующегося в тканях СО2 ковалентно связываются с N-концом гемоглобина и транспортируется как карбамино-НЬ. Около 90% СО2 превращается в более растворимый гидрокарбонат (НСОз-). В легких из него снова регенерируется СО2, который выводится легкими. Высокая концентрация СО2, существующая в тканях с интенсивным обменом веществ, увеличивает локальную концентрацию Н+ и снижает сродство гемоглобина к О2 (эффект Бора). Это ведет к усиленному освобождению О2 и вместе с тем к лучшему снабжению кислородом. Без катализатора равновесие между СО2 и НСОз- устанавливается относительно медленно. В эритроцитах эта реакция ускоряется карбонат-дегидратазой («карбоангидразой»), присутствующей в достаточно высокой концентрации.
ГУМОРАЛЬНЫЙ ИММУНИТЕТ ГАММАГЛОБУЛИНЫ Иммунитет – это комплекс реакций, направленных на защиту организма от инфекционных агентов и веществ, отличающихся биологическими (антигенными) свойствами. Иммунный ответ состоит из сложного ряда клеточных взаимодействий, активируемых попаданием в организм чужеродного антигенного материала. После обработки макрофагами, антиген предоставляется лимфоцитам, которые являются главными клетками исполнительного звена иммунной системы. Активация лимфоцита антигеном приводит к пролиферации и трансформации лимфоцитов. Существует два главных типа иммунного ответа: Клеточный иммунитет – это функция T-лимфоцитов; при клеточном иммунитете происходит образование эффекторных клеток – T-киллеров, способных уничтожать клетки, имеющие антигенную структуру, путем прямой цитотоксичности и путем синтеза определенных веществ, названных лимфокинами, которые участвуют в процессах взаимодействия клеток (макрофагов, T-клеток, B-клеток) при иммунном ответе. Кроме того, два подтипа T-клеток участвуют в регуляции иммунного ответа: T-хелперы усиливают иммунный ответ; T-супрессоры оказывают противоположное влияние. Гуморальный иммунитет – это функция B-клеток и характеризуется преобразованием B-клеток в плазматические клетки, секретирующие иммуноглобулины (антитела), которые имеют специфическую активность против внедрившегося антигена.
Антитела - это гаммаглобулины, способные соединяться с гомологичными антигенами, вызывать лизис микробов, фиксировать комплемент, проникать через физиологические барьеры. Согласно международной классификации антитела называют иммуноглобулинами и обозначают символом Ig. Иммуноглобулины представляют собой молекулы белка, образованные комбинациями аминокислот (всего 12 видов). Они имеют небольшую рецепторную группу, с помощью которой прочно соединяются со специфическими антигенами. Большинство молекул иммуноглобулинов составлены из двух тяжелых (H) цепей и двух легких (L) цепей, соединенных дисульфидными связями. Легкие цепи состоят или из двух k цепей, или из двух l цепей. Тяжелые цепи могут быть одного из пяти классов IgA, IgG, IgM, IgD, и IgE - 5 видов иммуноглобулинов. Существует несколько подклассов тяжелых цепей (изотипы). Эти различные цепи иммуноглобулинов являются антигенами для животных и имеют отличающиеся антигенные детерминанты, поэтому, при введении их животным, антитела, производимые против них, могут использоваться для распознавания и определения различных типов легких цепей и классов тяжелых цепей у человека. Классы иммуноглобулинов
Каждая цепь имеет постоянный и вариабельный участок. Постоянный участок остается постоянным в последовательности аминокислот и антигенности в пределах данного класса иммуноглобулинов; вариабельный участок, напротив, характеризуется большой непостоянностью последовательности аминокислот. Именно в вариабельной части цепи происходит реакция соединения с антигеном.
Рис. 1 Строение иммуноглобулинов (IgG)
Каждая молекула IgG состоит из двух соединенных цепей, которые формируют два антигенсвязывающих участка. На вариабельном участке каждой цепи имеются гипервариабильные участки – три в легких цепях и четыре в тяжелых цепях. Разновидности последовательности аминокислот в этих гипервариабильных участках определяют специфичность антитела. При определенных условиях эти гипервариабильные области могут также выступать в роли антигенов (идиотипы). Антитело против идиотипов, т.е. производимое против гипервариабильной области антител, имеет ограниченный диапазон реактивности и соединяется только с молекулами иммуноглобулина, имеющими данную гипервариабильную область. В сущности, реактивность антител против идиотипов ограничена исключительно специфическими антителами, полученными из единственного клона. Хотя вышеописанное относится строго к IgG, другие классы иммуноглобулинов имеют такую же основную структуру, за исключением того, что IgM является пентамером (т.е. состоит из 5 основных единиц (молекул), связанных в области Fc-концов), а IgA обычно существует как димер. IgD и IgE имеют сходную структуру. IgA – димер, а IgM – пентамер. Антиген-связывающие места формируются концами тяжелой и легкой цепей; каждая молекула IgG имеет два места связывания. При энзиматическом расщеплении образуются следующие фрагменты: Fc-фрагмент содержит участки обеих постоянных частей; Fab-фрагмент содержит легкую и часть тяжелой цепи с одним антиген-связывающим участком. F(ab)’2-фрагмент состоит из двух связанных между собой Fab-фрагментов. Н - область «вилки»; HV, LV - вариабельные концы тяжелой и легкой цепей; HC, LC - постоянные концы тяжелой и легкой цепей. Постоянный участок каждой молекулы иммуноглобулина имеет рецепторы для комплемента, а также имеется на Fc-фрагменте участок, который связывается с клетками, имеющими Fc-рецепторы (что необходимо для осуществления клеточного иммунитета). Унаследованные антигенные различия между тяжелыми цепями составляют аллотипы. Молекулы иммуноглобулинов можно разбить на части различными протеолитическими ферментами. При воздействии папаина молекула разделяется в области расхождения тяжелых цепей (“вилки”) на два Fab-фрагмента и один Fc-фрагмент (кристаллизующийся). Пепсин разрывает молекулу на F(ab)’2-фрагмент и Fc-фрагмент. Fc-фрагмент представляет собой постоянный участок; отсутствие изменяемости последовательности аминокислот – главная причина возможности кристаллизации данного фрагмента. Fab и F(ab)’2-фрагменты несут один и два антиген-связывающих участка соответственно. Fc-фрагмент несет специфические антигены, включая те, которые определяют иммунологическое различие пяти главных классов антител. Участок фиксации комплемента также расположен на Fc-фрагменте. Метод ферментативного расщепления имеет историческое значение в процессе выяснения структуры иммуноглобулинов. IgA — в основном участвуют в реализации иммунных реакций на поверхности слизистых оболочек. Поэтому им принадлежит важная роль в защите организма от микробов, которые проникают в него через слизистые оболочки. IgM относятся к макроглобулинам. Они нейтрализуют микробов в крови. Данная реакция протекает с участием комплемента. Обычно для разрушения одной микробной клетки достаточно одной молекулы IgM. Подобного эффекта можно достигнуть и с помощью IgG. Но в этом случае должно участвовать не менее 2 молекул антител или даже больше. IgG являются преимущественно антитоксинами и составляют основную часть из всего количества циркулирующих в крови антител. Значение иммуноглобулинов D до настоящего времени не раскрыто. Их также называют реагинами и цитофильными антителами. Последнее их название связано с тем, что они обладают сродством к тучным клеткам организма. Адсорбируясь на этих клетках, IgE, как правило, не циркулируют в крови или же находятся в ней в весьма незначительном количестве. В таком случае при проникновении в организм антигенов, к которым уже имеются антитела, относящиеся к IgE, иммунная реакция с их участием протекает не в крови, а на поверхности тучных клеток и приводит к их разрушению. Высвобождающиеся при этом в большом количестве гистамин, серотонин, брадикинин и другие биологические вещества, которыми богаты тучные клетки, обусловливают картину реакции немедленного типа, классическим проявлением которой является анафилактический шок.
Синтез иммуноглобулинов Иммуноглобулины синтезируются плазматическими клетками, которые образуются из трансформированных, стимулированных антигеном B-лимфоцитов (B-иммунобластов). Все молекулы иммуноглобулинов, синтезированных отдельной плазматической клеткой, идентичны и имеют специфическую реактивность против единственной антигенной детерминанты. Аналогично, все плазматические клетки, полученные путем трансформации и пролиферации одного B-лимфоцита-предшественника, идентичны; то есть, они составляют клон. Молекулы иммуноглобулинов, синтезированные клетками различных клонов плазматических клеток, имеют различные последовательности аминокислот, что обусловливает различную третичную структуру молекул и придает иную специфичность антителу, то есть, они реагируют с разными антигенами. Эти различия в последовательности аминокислот происходят в так называемом V (вариабельном, переменном) участке молекулы иммуноглобулина. Регулирование производства антител: производство антител начинается после активации B-клеток антигеном. Максимальная концентрация антител в сыворотке наблюдается с 1 по 2 неделю и затем начинает снижаться. Непрерывное присутствие свободного антигена поддерживает ответ до тех пор, пока увеличение уровня антител не приведет к усиленному удалению антигена и, таким образом, прекращению стимуляции B-клеток. Существуют также более тонкие механизмы регуляции синтеза иммуноглобулинов. T-хелперы (CD4-позитивные) играют важную роль в регуляции ответа В-клеток на большое количество антигенов и их постоянное присутствие увеличивает производство антител. Этот эффект возникает благодаря высвобождению лимфокинов. T-супрессоры (CD8-позитивные) оказывают противоположное влияние, вызывая снижение иммунного ответа; сильное подавление ответа может быть одним из механизмов, лежащих в основе толерантности. Одним из дополнительных регулирующих механизмов является выработка анти-идиотипов (т.е. антител против собственных антител (аутоантител)). Предполагается, что при иммунном ответе производство специфического антитела обязательно сопровождается производством второго антитела (анти-идиотипного) со специфичностью против вариабельных (V) последовательностей (идиотипов или антиген-связывающих участков) первого антитела. Анти-идиотипное антитело способно к распознаванию идиотипов на антигенном рецепторе B-клеток (который построен из иммуноглобулина, идентичного по строению идиотипу первого антитела), таким образом, оно конкурирует с антигеном и служит для ингибирования активации B-клетки. Следует отметить, что иммуноглобулины синтезируются не только при инфекционных заболеваниях. Они продуцируются непрерывно у каждого здорового человека. В результате в организме людей имеется определенный уровень различных видов антител, практически против всех микробных антигенов, в том числе и против тех возбудителей, с которыми они никогда не встречались. Это объясняется тем, что способность организма к синтезу антител выработалась у людей в процессе эволюционного развития и является генетически обусловленной. Эти антитела (иммуноглобулины) носят название нормальных. Нормальные антитела играют большую роль в защите организма от инфекции в момент внедрения возбудителей в организм, а также в начальный период болезни (т. е. тогда, когда иммунные реакции на инфекцию еще не успели сформироваться). Обычно первые проявления инфекционного иммунитета появляются не раньше 4-го дня с момента заболевания и достигают максимальной выраженности на 14 сутки и позже. Заслуживает отдельного внимания тот факт, что продуцируемые подэпителиально расположенными лимфоцитами антитела секретируются не в кровь, а на поверхность слизистых оболочек. В то же время циркулирующие в крови антитела в норме не проникают на поверхность слизистых оболочек. Следовательно, лимфоидные клетки слизистых оболочек в значительной мере функционируют автономно. Секретируемые ими антитела образуют первую линию защиты организма от возбудителей инфекционных заболеваний. Антитела слизистых оболочек представлены в основном IgA. Только на поверхности миндалин, а также слизистой нижних отделов респираторного тракта, наряду с IgA, имеется довольно большая концентрация IgG. На модели стрептококковой инфекции установлено, что IgA блокируют рецепторы микробов, с помощью которых последние фиксируются к эпителиальным клеткам и получают условия для размножения. Биологическое значение IgG определяется их выраженным опсонизирующим эффектом. IgA не секретируются только у новорожденных детей.
МОЛЕКУЛЯРНЫЕ ОСНОВЫ МЫШЕЧНОГО СОКРАЩЕНИЯ
МЫШЕЧНОЕ СОКРАЩЕНИЕ Модель скользящих нитей Скелетные мышцы состоят из многоядерных клеток, связанных возбудимой плазматической мембраной, по которой приходит нервный импульс, инициирующий сокращение мышцы. Мышечные клетки состоят из множества сократительных волокон - миофибрилл, расположенных параллельно друг другу. Структурно-функциональными единицами миофибрилл являются саркомеры, которые располагаются вдоль мышечных волокон через каждые 2, 3 мкм. На электронно-микроскопических снимках продольного среза мышечной ткани видно, что саркомер состоит из параллельных рядов толстых и тонких нитей. Взаимное расположение толстых и тонких нитей саркомера схематически показано на рис. 1, а. Вертикальные темные линии Z соответствуют специальным структурным белкам, разделяющим миофибриллы на саркомеры. Между ними видны горизонтальные нити сократительного аппарата. От Z-линий отходят тонкие нити, которым на электронно-микроскопических снимках соответствуют светлые полосы I. В центральной части саркомера расположены толстые нити, которым соответствуют темные полосы А. В середине каждой полосы А видна более светлая полоса Н. Наличие двух темных участков полосы А определяется тем, что в этих зонах толстые нити перекрываются с тонкими нитями. Более светлая полоса (зона Н) соответствует участку саркомера, где толстые нити не перекрываются с тонкими нитями.
Рис. 1. Схематическое изображение строения саркомеров мышечного волокна: а - продольный разрез, б - поперечный разрез в области пересечения толстых и тонких нитей, в - изменение длины саркомера в результате движения толстых и тонких нитей.
Толстые нити, имеющие диаметр 15 нм, состоят главным образом из молекул миозина. Тонкие нити имеют диаметр 9 нм. Они содержат белки трех типов: актин, тропомиозин и тропониновый комплекс. Если посмотреть на поперечный срез саркомера в области, где соседствуют толстые и тонкие нити (темный участок полосы А), то можно увидеть, что каждая тонкая нить окружена тремя толстыми нитями, а каждая толстая нить окружена шестью тонкими нитями (рис. 1, б ). Толстые и тонкие нити взаимодействуют друг с другом с помощью поперечных мостиков длиной около 13 нм, которые через регулярные промежутки выходят из толстых нитей и заполняют щели между соседними толстыми и тонкими нитями. При сокращении мышцы ее длина укорачивается на одну треть. Как это происходит, стало понятно в начале 50-х годов, когда Эндрю и Хью Хаксли, Р. Нидергерк и Ж. Хэнсон на основании исследования структуры мышечных волокон методами рентгеноструктурного анализа, оптической и электронной микроскопии независимо пришли к модели скользящих нитей. В основе этой модели лежат следующие факты: 1) при сокращении мышцы длины толстых и тонких нитей саркомера не изменяются; 2) саркомер укорачивается за счет перекрывания толстых и тонких нитей, которые скользят друг относительно друга во время сокращения мышцы. Это проявляется в том, что при сокращении мышцы полосы H и I укорачиваются (рис. 1, в); 3) сила, развиваемая мышцей, создается в процессе движения соседних нитей. Скольжение толстых и тонких нитей друг относительно друга совершается за счет энергии, выделяемой при гидролизе АТР до ADP и неорганического фосфата (Pi). Открытие АТРазной активности миозина было сделано в 1939 году супругами В.А. Энгельгардтом и М.Н. Любимовой, которые показали, что препараты миозина способны расщеплять АТР на ADP и Pi. Ими было также показано, что добавление АТР к белковому препарату, состоящему из нитей миозина, влияет на его механические свойства. Вскоре после этого А. Стент-Дьорди (удостоенный впоследствии Нобелевской премии) установил, что в растворе актин и миозин образуют так называемый актомиозиновый комплекс. Примечательно, что при отсутствии актина миозин плохо гидролизует АТР. В присутствии актина АТРазная активность миозина возрастает приблизительно в 200 раз.
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 818; Нарушение авторского права страницы
 является первой существенной составляющей свободной энергии для дестабилизации глобулярных белков. В расчете на стабильный (развернутый) остаток Δ
является первой существенной составляющей свободной энергии для дестабилизации глобулярных белков. В расчете на стабильный (развернутый) остаток Δ 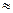 2-6
2-6  .
. (связано с разрушением водородных связей). Δ
(связано с разрушением водородных связей). Δ 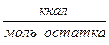 .
. при нарушении гидрофобного взаимодействия в ходе денатурации.
при нарушении гидрофобного взаимодействия в ходе денатурации. - NT Δ
- NT Δ  Δ
Δ  , где
, где 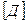 и
и  − концентрации денатурированного и нативного белка.
− концентрации денатурированного и нативного белка. O = Δ FД – Δ FN.
O = Δ FД – Δ FN.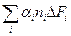 .
.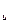 ), при которой [Д] = [N] (Δ Fu = 0), тогда
), при которой [Д] = [N] (Δ Fu = 0), тогда =−
=− 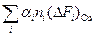 .
. , где Cp(T) − теплоёмкость при постоянном давлении.
, где Cp(T) − теплоёмкость при постоянном давлении.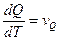 и регистрируют скорость повышения температуры образца
и регистрируют скорость повышения температуры образца  (
( 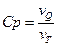 ).
).

 -
-  -
- 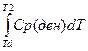 = Δ Нd -
= Δ Нd -  ,
, 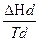 -
-  .
.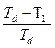 -
-  .
.