
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Основные этапы биосинтеза белка
Исследования, проведенные в различных лабораториях, показали, что синтез полипептидных цепей протекает в четыре стадии. На первой стадии синтеза белка, которая называется активацией, происходит активирование аминокислот путем связывания их с соответствующими тРНК. Этот процесс происходит в растворимой части цитоплазмы клеток за счет энергии АТФ. Вторая стадия биосинтеза белка называется инициацией полипептидной цепи. На этой стадии путем связывания иРНК и первой, или инициирующей, активированной аминокислоты со свободной 300 рибосомной 3OS-субъединицей, которая затем связывается рибосом-ной 50S-субъединицей, происходит образование инициирующего комплекса. На третьей стадии, называемой элонгацией, происходит удлинение полипептидной цепи путем последовательного присоединения новых аминокислот, которые приносятся к иРНК соответствующими тРНК. Причем каждая тРНК, переносящая определенную аминокислоту, специфически отбирается соответствующим участком иРНК — кодо-ном. После образования очередной пептидной связи молекула иРНК и образовавшаяся полипептидная цепь перемещается, или трансло-цируется на рибосоме. При этом следующий кодон иРНК занимает на рибосоме рабочее положение, к нему может присоединяться новая тРНК с принесенной аминокислотой. На стадии элонгации используется энергия, поставляемая в форме ГТФ. На четвертой стадии, которая называется терминацией полипептидной цепи, синтез белка заканчивается. Сигналами терминации служат определенные участки иРНК. Когда при транслокации они достигают рибосомы, синтез полипептидной цепи останавливается и она отделяется от рибосомы. Рассмотрим теперь более подробно весь процесс биосинтеза белка. Активация аминокислот. На первой стадии белкового синтеза 20 аминокислот, входящих в состав белков, активируются. Они соединяются с определенными тРНК под действием ферментов, называемых аминоацил-тРНК-синтетазами. При этом каждой аминокислоте соответствует свой специфический фермент. Активация аминокислот происходит в два этапа. Сначала аминокислота взаимодействует с АТФ, образуя аминоациладениловую кислоту: АТФ + Аминокислота На втором этапе происходит перенос аминоацильной группы с ами-ноациладениловой кислоты на тРНК. При этом образуется амино-ацил-тРНК и адениловая кислота: Аминоациладениловая кислота + тРНК Аминоацильная группа связывается с тРНК на том конце ее молекулы, который имеет последовательность нуклеотидов ЦЦА. Напомним, что на противоположном конце молекулы тРНК, которая имеет форму клеверного листка, находится особый участок, состоящий из трех нуклеотидов, или триплет, называемый антикодоном тРНК. Благодаря антикодону, имеющему определенную последовательность нуклеотидов, каждая тРНК узнает свое место на иРНК, а транспортируемая аминокислота занимает правильное место в растущей полипептидной цепи. Инициация полипептидной цепи. Установлено, что полипептидная цепь строится, начиная с NН2-концевой аминокислоты, т.е. первая аминокислота вступает в образование пептидной связи своей карбоксильной группой. Кроме того, синтез большинства, а возможно, и всех белков начинается с аминокислоты метионина. Однако инициирующий остаток метионина включается в начало белкового синтеза не в виде метионил-тРНК, а как N-формилметионил-тРНК. Иначе говоря, в молекуле метионина аминогруппа блокирована формильной группой. Поэтому данная кислота и не участвует в образовании пептидной связи своей аминогруппой. Остаток N-формилметионила не остается в составе синтезированной полипептидной цепи, он отщепляется после окончания белкового синтеза. Участие метионина в виде формилметионил-тРНК в начале белкового синтеза исключает, очевидно, возможность синтеза рибосомой пептидной цепи с середины иРНК. Очень важным обстоятельством в инициации полипептидной цепи являются постоянная диссоциация 70 S-рибосомы на 50 S- и 30 S-субъ-единицы и постоянная реассоциация этих субъединиц с образованием 70 S-рибосомы. Диссоциация и реассоциация 70 S-рибосомы имеет важное биологическое значение, состоящие в том, что рибосома начинает синтез полипептидной цепи именно с начала молекулы иРНК, а не в любом ее месте. Процесс инициации осуществляется после диссоциации 70 S-рибосомы на 50S-и 30 S-субъединицы. Он начинается с образования комплекса между 30 S-субъединицей рибосомы, иРНК и формилметионил-тРНК. В этом процессе участвует ГТФ и три вещества белковой природы, называемых факторами инициации. К образовавшемуся инициирующему комплексу далее присоединяется 50 S-субъединица, в результате чего образуется полная функциональная 70 S-рибосома, связанная с началом полинуклеотидной цепи и РНК. Формилметионил-тРНК, поступившая вначале в аминоацильный участок рибосомы, перемещается затем в пептидильный участок. На этом процесс инициации заканчивается. Элонгация полипептидной цепи. В момент окончания процесса инициации формилметионил-тРНК расположена в пептидильном участке рибосомы, аминоацильный участок свободен и готов к принятию следующей аминоацил-тРНК. Стадия элонгации состоит из трех этапов. На первом этапе в аминоацильный участок 70S-рибосомы поступает новая аминоацил-тРНК, которая связывается со следующим по порядку кодоном иРНК. Этот этап протекает с использованием энергии, поставляемой в виде ГТФ. На втором этапе на аминоацильном участке 70 S-рибосомы происходит образование пептидной связи между новой аминоацил-тРНК и формил-метионил-тРНК. Последняя перемещается на этот участок с пепти-дильного. При этом освободившаяся от формилметионильного остатка тРНК остается все еще связанной с пептидильным участком. Образование пептидной связи происходит без участия АТФ и ГТФ, но под действием фермента пептидилтрансферазы, которая является частью 50 S-субъединицы рибосомы. На третьем, заключительном этапе элонгации образовавшийся дипептид, связанный с тРНК, которая принесла новую аминокислоту, перемещается с аминоацильного участка на пептидильный, вытесняя из него освободившуюся ранее от формилметионильного остатка тРНК. Процесс перемещения дипептида (или полипептида) с аминоацильного участка на пептидильный называется транслокацией. Он сопровождается затратой энергии в форме ГТФ. В процессе транс- локации происходит освобождение аминоацильного участка рибосомы, к которому присоединяется следующая аминоацил-тРНК, и весь процесс повторяется. Механизм элонгации поли пептидной цепи остается одинаковым на протяжении всего процесса биосинтеза белка. Терминация полипептидной цепи. Считывание информации с иРНК, т.е. трансляция, и рост полипептидной цепи продолжается до последнего С-концевого остатка ее молекулы. Сигналом для окончания синтеза служит поступление на аминоацильный участок рибосомы одного из так называемых «бессмысленных» кодонов (триплетов УАА, УАГ и УГА), которые не транслируют ни одной аминокислоты. Один из таких триплетов (или два подряд) расположены на иРНК сразу за кодоном последней аминокислоты. Процесс терминации изучен в меньшей степени, чем другие стадии белкового синтеза. Однако известно, что сначала полипептидная цепь перемещается с аминоацильного участка, где образовалась последняя пептидная связь, на пептидильный. Затем происходит разрыв связи между концевой аминокислотой и ее тРНК и полипептидная цепь покидает рибосому. После этого распадается комплекс рибосома — иРНК, а сама рибосома диссоциирует на две субъединицы, которые снова используются в биосинтезе. Синтезированная полипептидная цепь, попадая в цитоплазму, приобретает свойственную ей пространственную конфигурацию.
Весь процесс биосинтеза белка можно изобразить схематически (рис. 47). Как видим, биосинтез белка в клетке осуществляется с участием нуклеиновых кислот, которые справедливо называют «живыми молекулами». Ведущее место в белковом синтезе принадлежит молекуле ДНК, в полинуклеотидных цепях которой в виде уникальной и специфической последовательности нуклеотидов заключена генетическая информация о последовательности аминокислот для миллионов синтезируемых белков. Молекулы ДНК хранят наследственную информацию и путем репликации передают ее из поколения в поколение. Доказательством ведущей роли ДНК в формировании специфических особенностей белка и его синтезе является тот факт, что при разрушении ДНК ядер нарушается или совсем останавливается белковый синтез. Это наблюдается при действии радиоактивного излучения, добавлении радиоактивного фосфора, а также при расщеплении ДНК под влиянием ферментов. Свою наследственную функцию ДНК выполняет не непосредственно, а при помощи РНК. Так, информация о структуре белка передается молекулой ДНК на иРНК, которая является посредником между ДНК и белками. Остальные РНК выполняют как бы подсобные функции: тРНК доставляет требуемые для белкового синтеза аминокислоты, другие РНК входят в состав рибосом и помогают списывать информацию с иРНК. Таким образом, нуклеиновые кислоты имеют чрезвычайно важное значение в белковом синтезе. Рассмотрим теперь, как происходит расшифровка генетического кода, т.е. каким образом нуклеиновые кислоты, состоящие из четырех нуклеотидов, кодируют и передают информацию на аминокислотный язык белков. Генетический код В настоящее время твердо установлено, что информация о специфическом строении белковой молекулы заложена в последовательности нуклеотидов молекулы ДНК, которая называется кодом. Само собой разумеется, что передача информации не может осуществляться по принципу «один нуклеотид — одна аминокислота», поскольку таким путем можно закодировать только 4 аминокислоты. Двух нуклеотидов также не хватает, поскольку четыре нуклеотида могут дать только 16 различных сочетаний по два нуклеотида (42 = 16). Количество же различных сочетаний из трех нуклеотидов составляет уже 64 (43= 64) и, следовательно, 20 аминокислот могут быть закодированы с помощью триплетного кода. Таким образом, код для 20 аминокислот, входящих в состав молекул природных белков, должен быть как минимум триплетным. Первое экспериментальное подтверждение триплетности генетического кода было получено в 1961 г. Ф. Криком, а позднее было окончательно доказано М. Ниренбергом, П. Ледером и X. Кораной. Доказательство триплетности кода поставило новый вопрос: поскольку для двадцати аминокислот вполне достаточно двадцати триплетов, какова роль лишних (сорока четырех) кодонов? Как показали исследования, отдельные аминокислоты могут кодироваться не одним, а несколькими кодонами. Кодоны 20 аминокислот представлены в табл. 8. Таблица 8. Генетический код
Из таблицы следует, что только триптофан и метионин кодируются каждый одним триплетом, а на остальные аминокислоты приходится по 2, 3, 4 и даже по 6 кодонов. Из 64 триплетов 61 кодируют аминокислоты, а 3 оставшихся не кодируют никаких аминокислот — их назвали «бессмысленными кодонами». Роль этих кодонов состоит в подаче сигнала об окончании биосинтеза белка, т.е. стадии терминации, поэтому они называются еще кодонами терминации. Существование нескольких кодонов для одной и той же аминокислоты называют вырожденностью генетического кода. Это свойство генетического кода имеет важный биологический смысл. Если бы каждой аминокислоте соответствовал только один триплет, то при замене одного нуклеотида в кодоне он превращался бы в бессмысленный, что приводило бы к остановке белкового синтеза. Однако вследствие того, что код вырожден, т.е. почти каждой аминокислоте соответствует несколько кодонов, то замена одного нуклеотида в кодоне приводит лишь к замене одной аминокислоты на другую и синтез белка не останавливается. При этом, если замена аминокислоты произошла не в активном центре белка, его биологические свойства не нарушаются. Из таблицы генетического кода видно, что код вырожден неравномерно. Например, для серина и лейцина он вырожден шестикратно (существует 6 кодонов для серина и 6 — для лейцина), а для таких аминокислот, как тирозин, гистидин, глутаминовая кислота и др., код вырожден лишь двукратно. Нетрудно заметить, что во многих случаях вырожденность кода касается только третьего нуклеотида в кодоне, причем это положение занято либо пиримидиновым, либо пуриновым основанием. Другими словами, кодирование одной аминокислоты определяется главным образом двумя первыми нуклеотида-ми, а третий нуклеотид не имеет существенного значения. Поэтому можно считать, что генетический код не триплетный, а 21/2. Важной особенностью генетического кода является отсутствие «знаков препинания» между кодонами. В связи с этим считывание информации с иРНК должно начинаться со строго определенного места, иначе вся последовательность нуклеотидов будет прочитана неправильно, что приведет к биосинтезу «бессмысленного» белка с неправильной структурой. Необходимо также отметить, что генетический код универсален — триплеты, кодирующие одни и те же аминокислоты, одинаковы у всех живых организмов, даже у таких очень отдаленных видов, как человек, растения и бактерии. ОБМЕН НУКЛЕИНОВЫХ КИСЛОТ Нуклеиновые кислоты являются составной частью нуклеопротеидов и входят в состав пищевых продуктов. Особенно богаты нуклеиновыми кислотами продукты животного происхождения. Под действием пищеварительных соков пищевого канала нуклеопротеиды расщепляются на простые белки и нуклеиновые кислоты. Белки расщепляются до свободных аминокислот уже известным нам путем, а нуклеиновые кислоты под влиянием ферментов, называемых нуклеазами, расщепляются на отдельные мононуклеотиды. Образовавшиеся мононуклеотиды распадаются далее на нуклеозиды и фосфорную кислоту под действием нуклеотидаз. Нуклеозиды и отдельные мононуклеотиды являются теми формами нуклеиновых кислот, которые всасываются стенкой кишок и включаются вклеточный обмен. В результате полного расщепления мононуклеотидов образуются азотистые основания (пуринового и пиримидинового ряда), пентозы (рибоза или дезоксирибоза) и фосфорная кислота. Эти продукты могут превращаться всоответствующие конечные продукты или же использоваться организмом для биосинтеза различных соединений, в том числе нуклеотидов, нуклеиновых кислот и нуклеопротеидов. Фосфорная кислота используется для синтеза АТФ, фосфатидов, костной ткани. Пентозы через ряд промежуточных реакций превращаются в СО2 и Н2О или вступают в другие реакции. Что же касается азотистых оснований, то они подвергаются превращениям, в результате которых из пуриновых оснований образуются мочевая кислота и аллантоин, из пиримидиновых — мочевина, щавелевая кислота и углекислый газ. Превращение пуриновых оснований. Превращение аденина и гуанина сопровождается дезаминированием и последующим их окислением в оксипурин. Аденин под действием аденазы дезаминируется до гипоксантина, который превращается затем в ксантин, гуанин дезаминируется гуаназой до ксантина. Ксантин окисляется далее ксантиноксидазой до мочевой кислоты:
Мочевая кислота является конечным продуктом обмена пуриновых оснований у человека и человекообразных обезьян и выделяется из организма вместе с мочой. У других животных (лошадей, собак, кошек, кроликов) мочевая кислота окисляется дальше до аллантоина, который является у них конечным продуктом обмена пуриновых оснований:
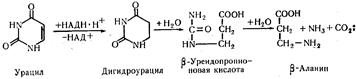
Превращение пиримидиновых оснований. Превращения цитозина, урацила и тимина по сравнению с пуриновыми основаниями имеют некоторые особенности. Пиримидиновые основания в тканях и печени сначала восстанав-ливаются, а затем расщепляются на более простые вещества. Цитозин сначала дезаминируется и превращается в урацил:
Урацил превращается в β -аланин, при этом освобождаются аммиак и углекислый газ:
Тимин образует β -аминоизомасляную кислоту, аммиак и углекислый газ:
Образовавшиеся β -аминокислоты (β -аланин и β -аминоизомасляная кислота) превращаются затем в мочевину, выделившиеся при распаде пиримидиновых оснований аммиак и углекислый газ также используются для биосинтеза мочевины. Таким образом, основными азотсодержащими конечными продуктами обмена нуклеиновых кислот и нуклеопротеидов в организме человека являются мочевая кислота и мочевина. Биосинтез нуклеотидов Поскольку мононуклеотиды состоят из азотистых оснований (аденина, гуанина, урацила, цитозина и тимина), рибозы или дезоксирибозы и фосфорной кислоты, для синтеза отдельных мононуклеотидов необходимо наличие в клетке этих составных компонентов. Пентозы и фосфорная кислота, поступая с пищевыми продуктами, находятся в организме в достаточном количестве. Основным путем обеспечения организма азотистыми основаниями является их синтез в клетках тканей человека и высших животных. Предшественники пуриновых и пиримидиновых оснований. Методом меченых атомов установлено, что для синтеза азотистых соединений пуринового ряда необходимы следующие вещества-предшественники; аспарагиновая кислота, глутамин, муравьиная кислота, глицин и СО2. В нижеприведенной формуле пурина показано, за счет каких перечисленных соединений образуются отдельные части пуринового кольца: Для синтеза пиримидиновых оснований используются аспарагиновая кислота, углекислый газ и аммиак. На первом этапе из аммиака и углекислого газа с участием АТФ образуется карбамоилфосфорная кислота, которая образует одну из частей пиримидинового кольца. Остальная часть этого кольца образуется за счет аспарагиновой кислоты. Синтез пиримидиновых оснований осуществляется через промежуточное соединение — оротовую кислоту, которая образует одно из пиримидиновых азотистых оснований — урацил. Ниже показан цикл пиримидина и превращение оротовой кислоты в урацил:
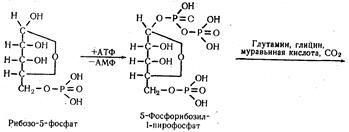
Урацил является азотистым основанием мононуклеотида — уридиловой кислоты, которая путем аминирования превращается в цитидиловую кислоту. Последняя в результате метилирования превращается в тимидиловую кислоту. Биосинтез мононуклеотидов. Пуриновые мононуклеотиды синтезируются не путем простого соединения их основных компонентов между собой в порядке азотистое основание — пентоза — фосфорная кислота. Их синтез начинается с пирофосфатного фосфорилирования рибозо-5-фосфата за счет энергии АТФ. Затем к полуацетальному гидроксилу пентозы в определенной последовательности присоединяются вещества, участвующие в построении кольца пуринового основания:
В результате этих последовательных реакций образуется инозиновая кислота, которая с участием аспарагиновой кислоты и АТФ превращается в адениловую. При взаимодействии инозиновой кислоты с глутамином образуется гуаниловая кислота. Таким образом, при синтезе пуриновых мононуклеотидов кольцо азотистого основания формируется на фосфорилированной пентозе. Синтез пиримидиновых мононуклеотидов, в отличие от пуриновых, происходит после того как сформируется кольцо пиримидинового основания, а именно, когда из карбамоилфосфата и аспарагиновой кислоты образуется оротовая кислота. К кольцу оротовой кислоты присоединяется рибозо-5-фосфат, в результате чего образуется оротидил-5-фосфат. Последний, декарбоксилируясь, превращается в уридиловую кислоту (пиримидиновый мононуклеотид). Уридиловая кислота в форме УТФ, образовавшегося за счет энергии АТФ, служит предшественником других пиримидиновых нуклеотидов: УМФ + АТФ Уридинтрифосфат путем аминирования с участием энергии АТФ образует цитидинтрифосфат: УТФ + NH3 Путем метилирования уридиловой кислоты образуется тимидиловая кислота. |
Последнее изменение этой страницы: 2017-03-14; Просмотров: 1242; Нарушение авторского права страницы
 Аминоациладениловая кислота + H4P2O7.
Аминоациладениловая кислота + H4P2O7. Рис. 47. Схема биосинтеза белка.
Рис. 47. Схема биосинтеза белка.






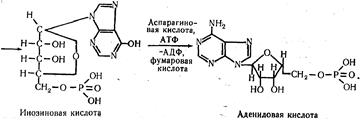
 ЦТФ.
ЦТФ.