
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Конечные продукты распада аминокислот
В результате различных превращений аминокислот в тканях организма образуются аммиак, углекислый газ и вода. После дезаминирования α -аминокислот образуются безазотистые соединения. Мы уже знаем, что в основном это α -кетокислоты, которые подвергаются дегидрированию и декарбоксилированию, и в конечном итоге превращаются в Н2О и СО2. Некоторые кетокислоты, подвергаясь окислительному декарбоксилированию, образуют кислоты с различной химической структурой, в зависимости от которой в одних случаях они окисляются по пути β -окисления, в других — по пути превращения углеводов. Однако и в этих случаях в конце концов образуются СО2 и Н2О. Углекислый газ частично выводится из организма, оставшаяся часть используется для синтетических процессов, например, для синтеза высших жирных кислот, пуриновых оснований и др. Обмен аммиака. Основным источником аммиака в организме являются аминокислоты. Аммиак — сильное токсическое вещество. Даже незначительное увеличение его количества в крови вызывает отравление организма, в особенности нервной системы. Ежесуточно в организме человека дезаминируется около 100 — 120 г аминокислот, что соответствует примерно 18—23, 6 г аммиака. Однако несмотря на непрерывное образование аммиака в тканях и поступление в кровь, концентрация его в крови очень невелика. Даже в капиллярах почек, где количество аммиака наибольшее, его концентрация в норме не превышает 1 мг %. В ходе эволюции у человека выработались специальные механизмы для его обезвреживания. К ним относятся образование глутамина, аспарагина, амидов белков, восстановительное аминирование, нейтрализация кислот, синтез мочевины. Одним из важнейших механизмов обезвреживания аммиака является образование глутамина и аспарагина. Синтез этих амидов имеет для организма большое значение, поскольку протекает в местах непосредственного образования аммиака: в печени, мозгу, почках, сетчатке глаза, мышцах. Глутаминовая и аспарагиновая кислоты, всегда находящиеся в тканях в свободном состоянии, связывают аммиак путем реакции амидирования. В результате из глутаминовой кислоты образуется глутамин, из аспарагиновой — аспарагин. Эти реакции протекают при помощи ферментов глутамин- и аспарагинсинтетаз:
Обезвреженный таким путем аммиак может быть использован организмом в качестве источника азота, например, для синтеза пуриновых и пиримидиновых оснований, гликозамингликанов и т.д. В тканях человека существует еще один ферментативный механизм обезвреживания аммиака — связывание его путем амидирования карбоксильных групп белков тканей. Амидирование белков обусловлено наличием в их составе свободных карбоксильных групп глутаминовой и аспарагиновой кислот. Часть аммиака обезвреживается путем реакций восстановительного аминирования — процесса, обратного дезаминированию, что обеспечивает связывание аммиака кетокислотами с образованием соответствующих аминокислот. Основная роль в этом принадлежит α -кетоглутаровой кислоте. Небольшое количество аммиака используется для нейтрализации неорганических и органических кислот, в результате чего образуются аммонийные соли. Этот процесс обезвреживания аммиака имеет место только в почках. Основной же путь обезвреживания аммиака связан с образованием мочевины. Синтез мочевины является основным путем детоксикации аммиака у млекопитающих и главной формой выделения белкового азота из организма. Мочевина, представляющая собой нейтральное соединение, растворимое в воде, выводится из организма с мочой. На долю азота мочевины приходится от 80 до 85 % общего количества азотистых веществ мочи. Впервые схему синтеза мочевины предложил М.В. Ненцкий. Он доказал, что синтез этого соединения происходит за счет двух молекул аммиака и одной молекулы углекислого газа. С помощью специальных опытов И. В. Залесский и С. С. Салазкин установили, что основным органом, где происходят обезвреживание аммиака и синтез мочевины, является печень. Образование мочевины протекает с участием ряда ферментов и веществ по определенному циклу, существование которого было доказано В. А. Кребсом и К. Хенселайтом, в связи с чем этот цикл получил название цикла мочевины, или цикла Кребса — Хенселайта. В цикле мочевины выделяют два основных этапа: первый сопровождается синтезом аргинина, второй гидролизом аргинина с образованием мочевины и орнитина. В настоящее время установлено, что источником одной из аминогрупп молекулы мочевины является аммиак, образующийся в процессе окислительного дезаминирования глутаминовой кислоты в митохондриях: Глутаминовая кислота + НАД+ + Н2О → α -Кетоглутаровая кислота + НАД•Н + Н+ + NH3. Этот аммиак вместе с СО2 образует соединение карбамоилфосфат. Реакция протекает под действием фермента карбамоилфосфатсинтетазы с использованием энергии двух молекул АТФ:
Карбамоилфосфат далее реагирует с орнитином, в результате чего образуются цитруллин и фосфорная кислота. Реакция катализируется ферментом орнитинтранскарбамилазой:
На следующей стадии в цикл мочевины вовлекается вторая аминогруппа, которая является аминогруппой аспарагиновой кислоты, образовавшейся из глутаминовой под действием фермента аспартат: глутамат — аминотрансферазы: Глутаминовая кислота + Щавелево-уксусная кислота → α -Кетоглутаровая кислота + Аспарагиновая кислота. Таким образом, источником аммиака для синтеза мочевины в конечном счете является глутаминовая кислота, в которой собран весь фонд аминогрупп в результате реакций переаминирования между α -амино-кислотами, образовавшимися при гидролитическом расщеплении белков в пищевом канале, и α -кетоглутаровой кислоты. Этим еще раз подчеркивается важная биологическая роль глутаминовой кислоты в азотистом обмене. Следующие реакции сопровождаются превращением цитруллина варгинин. Сначала цитруллин взаимодействует с аспарагиновой кислотой с участием аргининосукцинат-синтетазы с образованием аргининоянтарной кислоты. В этой реакции используется энергия одной молекулы АТФ:
Затем аргининоянтарная кислота подвергается ферментативному расщеплению с образованием аргинина и фумаровой кислоты:
Реакция катализируется аргининосукцинатлиазой. На этом заканчивается первый этап синтеза мочевины. Фумаровая кислота возвращается в цикл трикарбоновых кислот, где превращается в щавелево-уксусную. Последняя, вступая в реакцию с глутаминовой кислотой, снова образует аспарагиновую, которая на известной нам уже стадии цикла мочевины передает свою аминогруппу цитруллину. На втором этапе образовавшийся аргинин под действием аргиназы гидролитически расщепляется на орнитин и мочевину:
Орнитин может снова вступать в реакцию с карбамоилфосфатом, и весь цикл начнется сначала. Таким образом, в организме человека и высших животных существуют довольно сильные механизмы обезвреживания аммиака и использование его для биосинтеза. Аммиак является не только конечным продуктом белкового обмена, но и строительным материалом для образования многих соединений. Качество белкового питания Белки, как уже известно, являются основным пластическим материалом нашего организма, который другими веществами заменить невозможно. Белки отличаются друг от друга содержанием отдельных аминокислот. Поэтому в зависимости от аминокислотного состава для покрытия потребностей организма в белке требуется разное количество различных белков. Другими словами, различные белки имеют неодинаковую биологическую ценность. Чем ближе аминокислотный состав того или иного белка к составу белков данного организма, тем выше его биологическая ценность. Наиболее ценными являются белки, в которых в достаточном количестве содержатся все необходимые организму незаменимые аминокислоты (т.е. такие, которые в организме совсем не синтезируются или синтезируются в очень малом количестве). Таким образом, биологическая ценность белка определяется качественным иколичественным составом аминокислот, входящих в его молекулу. Для человека ценными являются белки молока, мяса и куриного яйца, из растительных белков — белок картофеля, который по своей биологической ценности превышает не только белок хлеба, но в отношении некоторых аминокислот даже белок коровьего молока. В смешанной диете, состоящей из 110—120 г белков, 40—60 г жиров и 600—700 г углеводов, должно содержаться не менее 50% белков животного происхождения. Такая диета обеспечивает потребность организма во всех незаменимых аминокислотах. Биосинтез белка Одним из основных путей превращения аминокислот, образовавшихся в процессе расщепления пищевых белков в пищевом канале, является их использование для биосинтеза специфически видовых белков. Этот процесс с различной скоростью протекает во всех органах и тканях. Ежесуточно в организме человека образуется около 1, 3 г белка на 1 кг массы тела. Если учесть, что средняя масса человека составляет 70 кг, то за сутки в его организме синтезируется около 90 г белков. Общее же количество белков в организме составляет 12—14 кг. Таким образом, все они обновляются приблизительно за 135—155 сут. Синтез некоторых белков происходит в организме с очень большой скоростью. Например, за 1 с в крови человека разрушается и вновь синтезируется около 3 млн. эритроцитов, каждый из которых содержит около 300 молекул гемоглобина. В общем же синтез молекулы белка в живой клетке заканчивается за 2—3 с. Строение рибосомы. Биосинтез белка — сложный многоступенчатый процесс, который осуществляется на рибосомах — клеточных микроструктурах, расположенных в цитоплазме. По своей химической природе рибосомы являются нуклеопротеидами, состоящими из РНК (50—60 %) и белков (35—50 %). Рибосомы всех клеток обладают одинаковой структурой и отличаются лишь размерами и содержанием РНК. Каждая рибосома состоит из двух субъединиц (частей) разного размера, на которые она может диссоциировать, а затем снова ассоциировать. Диссоциация и ассоциация рибосом на две субъединицы, как будет показано ниже, является характерной особенностью этих микроструктур и непосредственно связана с механизмом биосинтеза белка. В рибосоме имеется два участка — аминоацильный, или А-учас-ток, и пептидильный, или П-участок. Аминоацильный участок служит для присоединения аминокислот, поступающих из цитоплазмы, которые необходимы для биосинтеза белка и образования пептидной связи между ними, пептидильный — для удержания растущей полипептидной цепи (рис. 44).
Наиболее изученными являются рибосомы бактериальных клеток. Вся рибосома бактериальной клетки обозначается как 70S образование, диссоциирующее на две субъединицы — 50S и 30S (рис. 45) (S - константа седиментации отдельных субъединиц и рибосомы в в целом). Субъединицы 50S и 30S имеют разный набор белков. Известно, что некоторые из них выполняют каталитические функции.
В клетке рибосомы находятся обычно в виде скоплений от 3 до 100 единиц, образуя полирибосому, или полисому. В полисоме отдельные рибосомы соединены между собой своеобразной нитью, видимой под электронным микроскопом. Этой нитью является информационная РНК (рис. 46). Каждая рибосома способна самостоятельно синтезировать одну полипептидную цепь, группа рибосом, т.е. полисома, — несколько таких цепей.
Матричный путь биосинтеза белка. Как известно, отличие одного индивидуального белка от другого определяется прежде всего природой и последовательностью чередования аминокислот, входящих в его состав, т.е. первичной структурой. Передача этих сведений и составляет одну из основных черт биосинтеза белка. Центром управления белкового синтеза является ядро клетки, в котором находится носитель всей информации — молекула ДНК. Информация закодирована в ДНК определенной последовательностью азотистых оснований, входящих в ее молекулу. Передача наследственных признаков, информации о первичной структуре белка осуществляется через информационную РНК (иРНК), которая синтезируется на определенном участке (гене) одной из цепей ДНК. В основе передачи информации лежит принцип комплементарности (см. гл. 15). Напомним, что синтез иРНК заключается в том, что молекула ДНК, имеющая форму двойной спирали, в отдельные моменты раскручивается на определенных участках (этот участок носит название гена). На каждом раскрученном участке двух нитей ДНК строится молекула иРНК по принципу комплементарности. Происходит своего рода «списывание» порядка чередования азотистых оснований раскрученного участка одной из нитей ДНК на молекулу РНК. Этот процесс «списывания» информации называется транскрипцией. В результате транскрипции порядок чередования азотистых оснований в молекуле иРНК комплементарно повторяет последовательность азотистых оснований раскрученного участка молекулы ДНК. Таким образом молекула иРНК получает информацию о первичной структуре белка. Попадая в цитоплазму, молекула иРНК «обрастает» рибосомами и служит матрицей, на которой строится белок в соответствии с заключенной в ней информацией о порядке аминокислот. Процесс передачи информации от иРНК, состоящей в строго определенной последовательности чередования нуклеотидов в ее молекуле на язык аминокислот молекул белка носит название трансляции. В связи с тем что для синтеза белка с определенной первичной структурой используется информация, заключенная в молекуле иРНК, которая служит матрицей, такой путь белкового синтеза называют матричным. Сама ДНК непосредственного участия в биосинтезе белка не принимает, она лишь снабжает иРНК всей необходимой информацией. Кратко матричный путь биосинтеза белка можно изобразить так; ДНК Процессы, связанные с репликацией ДНК, транскрипцией и трансляцией, начинаются с момента зарождения организма и с разной интенсивностью протекают на протяжении всей жизни. |
Последнее изменение этой страницы: 2017-03-14; Просмотров: 1231; Нарушение авторского права страницы

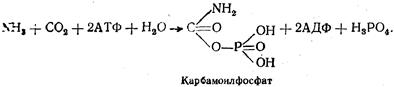
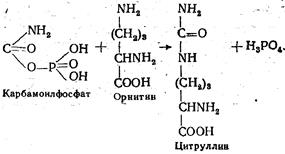
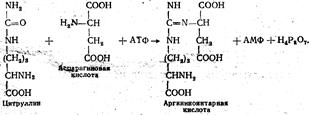

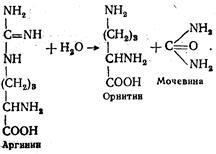
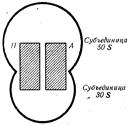 Рис. 44. Строение рибосомы.
Рис. 44. Строение рибосомы.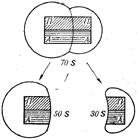 Рис. 45. Субъединицы 70 S-рибосомы.
Рис. 45. Субъединицы 70 S-рибосомы.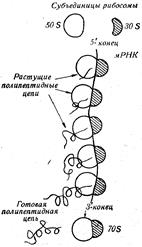 Рис. 46. Механизм функционирования полирибосомы.
Рис. 46. Механизм функционирования полирибосомы. РНК
РНК  Белок.
Белок.