|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Функциональных продуктов питания
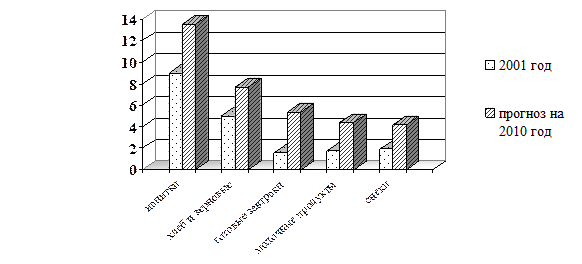
Мировой рынок функциональных продуктов интенсивно развивается, ежегодно он увеличивается на (15-20) % [64]. Мировым лидером в развитии функционального питания до настоящего времени остается Япония. Для европейских стран концепция функционального питания достаточно нова и отражает последние направления развития пищевой индустрии [124, 127]. Диаграммы на рис. 4 иллюстрируют тенденции, сложившиеся и сохраняющиеся в настоящее время [49]. Во всех группах функциональных продуктов наблюдается интенсивный рост объема производства [48, 126].
Рис. 4. Тенденции изменения рынка функциональных продуктов к 2010 году
Поразительная динамика развития рынка функциональных продуктов питания и напитков обусловлена двумя взаимосвязанными факторами - усилиями производителей, старающихся выпускать продукты с повышенной ценностью, и растущим спросом со стороны потребителей, которые, по результатам социологических опросов, весьма благожелательно относятся к товарам, несущим очевидную пользу для здоровья. Эти же опросы выявляют отчетливое предпочтение в пользу функциональных продуктов питания по сравнению с диетическими [83, 127]. Если говорить о классификации функциональных продуктов питания, то определившимися современными тенденциями на сегодняшний день следует считать: – продукция, позиционируемая как «Лучший выбор для Вашего здоровья», - пищевые продукты с акцентом на содержание только натуральных ингредиентов, не содержащие консервантов, с пониженным содержанием сахара и холестерина; – продукция, позиционируемая как возможность получить внешний «косметический эффект» – продукты с концепцией «Красота изнутри», «Результат на лицо», или продукты, способные сократить признаки старения; – концепции «Здоровый кишечник», «Здоровое сердце», «Улучшение мозговой деятельности», «Укрепление иммунитета» базируются на использовании микроорганизмов – пробиотиков, привлекают все больше внимания, становясь основой для значительного числа новых разработок во всех развитых странах; – продукты, специально фокусирующиеся на целевой группе «дети и подростки» - функциональные продукты, позволяющие вырастить здоровое поколение и максимально реализовать потенциал ребенка (например, продукты - синбиотики). В России наибольшую практическую реализацию нашли приемы коррекции микробной экологии человека, базирующиеся на использовании специально подобранных пробиотических микроорганизмов, как в составе поликомпонентных заквасок для кисломолочных напитков, так и в виде монокультур [44, 97, 123]. Причем в производстве таких продуктов большое внимание уделяется качеству и видовому составу микроорганизмов заквасочных культур, так как они вызывают основные биохимические изменения в молоке [4, 37, 98].
1.3. Микроорганизмы, используемые Микроорганизмы, входящие в состав кисломолочных продуктов, обуславливают лечебно- профилактические и диетические свойства, обладают широким спектром позитивного целенаправленного воздействия на организм человека, составляют основу микробиоценозов, характеризующихся определенным составом и занимающих тот или иной биотоп в организме человека [17, 99]. Большой интерес вызывает изучение, разработка и производство пробиотических продуктов, ферментированных молочнокислыми и пропионовокислыми бактериями, представителями нормальной микрофлоры кишечника [25]. Эти микроорганизмы обладают высокой метаболической активностью, участвуют в синтезе ряда витаминов, гидролизе желчных солей и холестерина, оказывают антагонистическое действие по отношению к условно-патогенной и патогенной микрофлоре (за счет изменения рН среды, продуцирования бактериоцинов, лишения нутриентов и мест адгезии конкурирующих микроорганизмов), благотворно влияют на пищеварение, устраняют дисбиотические нарушения [8, 75]. Чистая культура молочнокислой болгарской палочки Lactobacillus delbrueckii subsp. bulgariqus (рис. 5) в гидролизованном молоке представляет собой длинную палочку, а в молоке в момент свертывания – короткие палочки, расположенные по одиночке в более коротких или более длинных цепочках [74].
Рис. 5. Культура Lactobacillus delbrueckii subsp. bulgariсus [168]
В молоке при высокой температуре (45±1)0C с повышением кислотности клетки болгарской палочки удлиняются [99]. При наличии антибиотиков и других ингибирующих веществ в молоке или при отсутствии ростовых веществ, таких как витамины, аминокислоты и летучие жирные кислоты, клетки Lactobacillus delbrueckii subsp. bulgariсus становятся тоньше, удлиняются и в них появляются валютиновые зёрна [68]. Клетки болгарской палочки представляют собой короткие палочки с закруглёнными концами [14]. Длина клеток варьирует от 0, 8 до 1, 0 мкм. Они грамположительные, среди старых встречаются клетки, не окрашивающиеся по Граму. Клетки Lactobacillus delbrueckii subsp. bulgaricus окрашиваются краской метиленовым голубым, Леффлера и особенно хорошо контрастной краской Кантранджиева. Валютиновые зёрна окрашиваются метахроматично от синего до сине-красного Болгарская палочка свертывает молоко при 450С от 6 до 10 часов, придавая специфический кисломолочный вкус и запах, более выраженные при совместном культивировании со Streptococcus salivarius subsp. thermophilus. Сгусток молока, как правило, неслизистый, но при понижении температуры сквашивания до 300С, некоторые штаммы могут вызвать ослизнение сгустков [15]. Некоторые штаммы болгарской палочки образуют полисахариды, состоящие из 10% - 19% арабинозы, 4% -27% маннозы, 9%-15% глюкозы, 34%-36% галактозы, которые обуславливают достаточно эластичную консистенцию и плотную структуру продукта. При культивировании в молоке в течение 10 часов при 210С болгарская палочка образует от 2мг % до 20мг % этаналя; не восстанавливает этаналь в спирт [19]. Болгарская палочка обладает также протеолитической активностью, в результате деятельности которой в готовом продукте накапливаются различные растворимые азотистые вещества, участвующие наряду с другими соединениями в формировании запаха, вкуса и аромата; она образует продукты протеолиза, в первую очередь свободные аминокислоты (САК), до 15мг% [13, 73]. Многие амины молока, образующиеся при декарбоксилировании аминокислот, имеют неприятный запах и вкус, так как болгарская палочка продуцирует гистамин, тирамин, триптамин. Поэтому при селекции всех молочнокислых бактерий необходимо учитывать их декарбоксилазную активность и подбирать заквасочные культуры, которые не образуют горькие пептиды [73]. При идентификации болгарской палочки наибольшие разногласия возникают в определении способности микроорганизмов сбраживать углеводы. Это объясняются тем, что не существует штамма с точно определенной характеристикой его способности к сбраживанию углеводов. Кроме того, разные авторы работали с различными питательными средами и с неодинаковыми по чистоте углеводами, а также с различными видами и количеством штаммов. Но ни один автор не указывает на способность ее к сбраживанию трегалозы и салицина [15, 99]. Клетки Streptococcus salivarius subsp. thermophilus (рис. 6) имеют шарообразную или овальную форму и величину от 0, 7 до1, 0 мкм.
Рис. 6. Культура Streptococcus salivarius subsp. thermophilus [168]
Встречаются парами и в виде длинных S-образных цепочек. Неподвижны, грамположительны. В момент свертывания молока обнаруживаются клетки, интенсивно окрашивающиеся в синий цвет краской Леффлера или контрастной краской Кантранджиева [13, 99]. Термофильный стрептококк хорошо растет в цельном или обезжиренном коровьем молоке, особенно в овечьем молоке или в смеси коровьего и овечьего (1: 1). В молоке с антибиотиками или другими ингибирующими веществами клетки термофильного стрептококка удлиняются и приобретают форму короткой толстой палочки [15]. На твердых питательных средах образуются S-колонии. Поверхностные колонии круглые, с гладким светлым краем и слабозернистой структурой, величиной от 1, 0 до 1, 2 мкм. В глубине колонии – линзовидные. В жидкой питательной среде с глюкозой при оптимальной температуре развития термофильный стрептококк растет интенсивно с помутнением бульона [21, 73]. Оптимальная температура развития термофильного стрептококка от 400С до 450С. При 100С – не растет. При температуре свыше 540С развитие замедляется. Терморезистентен: выдерживает нагревание при 630С в течении 30 минут [74]. При оптимальных условиях развития термофильный стрептококк сбраживает лактозу, глюкозу, фруктозу, сахарозу. Не сбраживает мальтозу, инулин, глицерин, манит и салицин [15]. Данная культура обладает низкой липолитической активностью. Протеолитическая активность термофильного стрептококка ниже, чем у болгарской палочки (от 2% до 4%) и продуктами протеолиза являются гистидин, аргинин, тирозин, фенилаланин и лейцин [99]. Термофильный стрептококк свертывает молоко при 450С от 6 до 8 часов (при внесении 1%). Образует от 0, 9% до 1, 1% L(+) молочной кислоты. Чаще всего придает сгустку молока слабозернистую структуру и приятный кисломолочный вкус, более типично выраженный при его совместном культивировании с болгарской палочкой. Streptococcus salivarius subsp. thermophilus не разжижает желатин [14, 19]. Пропионовокислые бактерии объединены в род Propionibacterium (рис. 7), который входит в состав семейства Propionibacteriaceae. Другой род этого семейства – Eubacterium [28, 147].
Рис. 7. Культура рода Propionibacterium [169]
Они были открыты в 1906 году Орла-Енсеном и Фройденрайхом и выделены впервые из эмментальского сыра [133]. После исследований Дугласа и Гантера в род Propionibacterium стали включать виды анаэробных коринебактерий. Первым русским ученым, начавшим исследование пропионовокислых бактерий, стал А.Ф. Войткевич [28]. Пропионовокислые бактерии, применяемые в молочной промышленности, выделяются из молочных и других пищевых продуктов. Различают два основных типа данных микроорганизмов: – Propionibacterium freudenreichii ssp. freudenreichii – Propionibacterium freudenreichii ssp. shermanii [29]. Так более 60% штаммов, относящихся к Р. shermanii и Р. freudenreichii, выделены из эмментальского сыра [1]. Чистые культуры выделенных бактерий составляют три морфологические группы: Bacterium acidi propionici a, Bacterium acidi propionici b, Bacillus acidi propionici [43, 133]. Классические пропионовокислые бактерии характеризуются как аэротолерантные (микроаэрофильные) бактерии [112]. Они грамположительные; плохо окрашиваются метиленовой синью (в отличие от молочнокислых); каталазоположительные; факультативно анаэробные; неподвижные; неспорообразующие формы; диаметр от 0, 5 до 0, 8 μ m, длина от 0, 5 до 1, 5 μ m [135, 28]. Морфологически пропионовокислые бактерии довольно трудно дифференцировать. Колонии – на средах влажные, округлые, блестящие, маслянистые, некоторые штаммы образуют тяжелый тянущийся осадок. Цвет колоний – кремовый, желтый, оранжевый, красный, коричневый. Форма может быть округлая, расщепленная или с ответвлениями. В микроскопическом препарате можно наблюдать одиночные и попарно расположенные микроорганизмы или короткие V-образные или Y-образные цепочки. Молодые клетки в аэробных условиях имеют вид коротких палочек, в анаэробных условиях и при повышении температуры – становятся более длинными [13, 28]. От других видов пропионовокислые бактерии отличаются более высокой термоустойчивостью. Это связано с особенностью химизма состава клетки – цитоплазматическая мембрана многослойна и богата фосфолипидами. Липиды пропионовокислых бактерий не только входят в структурные элементы клеток, но и играют роль защитных компонентов против действия ряда антибиотиков. Состав клеточных фосфолипидов служит таксономическим признаком рода Propionibacterium. Основной фосфолипид представлен мономанозидом глицерил фосфорилмиоинозита. Общее количество фосфолипидов у пропионовокислых бактерий составляет до 10% от общего количества липидов [29, 112]. Пропионовокислые бактерии растут в пределах температуры от 150С до 400С. Оптимальная температура развития классических пропионовокислых бактерий (30±1)0С [110]. Оптимальная величина рН роста – от 6, 5 до 7, 0; максимальная - 8, 0; минимальная – от 5, 0 [28, 99]. При выделении пропионовокислых бактерий учитывают хороший рост их в анаэробных условиях при нейтральном значении рН. Анаэробные условия создают путем внесения сульфита Nа, цисцеина, тиогликолята Nа к агаровой среде [29, 112]. Пропионовокислые бактерии хорошо утилизируют лактаты. Лактат в качестве источника углерода обеспечивает более высокую скорость роста пропионовокислых бактерий. Пропионовокислые бактерии лучше всего используют лактат в присутствии безклеточного экстракта штаммов Streptococcus salivarius subsp. thermophilus [28]. При культивировании на средах пропионовокислые бактерии могут синтезировать аминокислоты за счет ассимиляции азота. Серосодержащие аминокислоты стимулируют рост пропионовокислых бактерий, проявляющих устойчивость к высоким концентрациям Н2S в среде. Из серосодержащих аминокислот пептидов молока бактерии образуют диметилсульфид, обладающий антимутагенными свойствами [27]. В результате брожения, зависимого от кофермента В12, род пропионовокислых бактерий образует пропионовую кислоту. Кроме пропионовой кислоты при брожении образуются: уксусная кислота, углекислый газ, диацетил и янтарная кислота [26, 28, 147]. При ферментации молока пропионовокислые бактерии проявляют слабую кислотообразующую активность: сквашивают молоко от 5 до 7 дней. В настоящее время теоретические и практические разработки в области изучения и производства молочных продуктов, в состав которых входят пропионовокислые бактерии, активно осуществляются в Восточно-Сибирском государственном технологическом университете, под руководством доктора технических наук, профессора Ирины Сергеевны Хамагаевой. Следует отметить некоторые особенности культивирования и размножения пропионовокислых микроорганизмов типа P.freudenreichii. Это строго анаэробные микроорганизмы. Все традиционные пропионовокислые бактерии вырабатывают каталазу. Для их развития требуются пантотенат, биотин и аминокислоты, присутствующие в молоке [28]. Пептидазактивные микроорганизмы представлены некоторыми штаммами лактобацилл. Во время созревания их число увеличивается вследствие декомпозиции протеинов до аминокислот, оба кофактора являются ключевыми в ферментации пропионовокислых микроорганизмов. Некоторые из пропионовокислых бактерий нуждаются дополнительно в тиамине (витамин В1, ) и р-аминобензокислоте [25]. Свободные аминокислоты не существенны для роста пропионовокислых микроорганизмов, но обладают стимулирующим эффектом. Это объясняет причину, почему L.helveticus и другие бактерии, обладающие сильным протеолитическим действием, стимулируют рост пропионовокислых бактерий [29]. Интерес заслуживает изучение влияния уровня рН на ферментацию пропионовокислых бактерий, которые растут сразу после помещения сыра в камеру охлаждения. Начальный уровень рН сыра оказывает действие на ферментацию пропионовокислых бактерий: чем выше уровень рН, тем быстрее развивается пропионовокислая микрофлора. Рекомендуемая кислотность (5, 35±0, 05) по рН после солильного бассейна перед камерой созревания [28, 43, 121]. Пропионовокислые бактерии не растут при уровне рН ниже, чем 5, 0. Сбраживание цитратов зависит от количества хлорида натрия и нитратов. Даже небольшое количество микроорганизмов сильно влияет на ферментацию. Рост уровня рН иногда зависит от остаточного уровня галактозы, в результате сбраживания которой увеличивается уровень пропионовокислого брожения [28, 77]. Оптимальная температура для роста от 300С до 320С, хотя строго ограниченный рост и метаболизм наблюдаются и между 30С и 100С. В зависимости от скорости формирования глазков температура в период созревания в теплой камере составляет от 200С до 24°С. Пропионовокислые бактерии ингибируются хлоридом натрия в зависимости от уровня рН. Быстрый рост пропионовокислых бактерий совершенно ингибируется при уровне рН 5, 2 и 7, 0 и содержании 3% и 6 % NaCl. На практике хлорид натрия содержится в сыре в количестве менее (1, 7 ± 0, 2) % [1, 43, 133]. Нитрат также ингибирует рост пропионовокислой микрофлоры. Нитрат применяется только в случае невозможности бактофугирования молока и дозах, не превышающих 7 г KN03 на 100 л молока [121]. Все традиционные пропионовокислые микроорганизмы в процессе метаболизма в состоянии расщеплять глицерин, фруктозу, глюкозу и галактозу до молочной кислоты. Сахароза может быть расщеплена только P.jensenii, P.theonii и P.acidi propionici, лактоза - P.freudenreichii ssp. shermanii, хотя все штаммы способны использовать лактат как субстанцию для начала метаболизма. Конечными продуктами их метаболизма являются пропионовая и уксусная кислота, СО2, и небольшое количество сукцината [77, 112]. Теоретически уравнение реакции для пропионовокислой ферментации лактата выглядит следующим образом:
3 молекулы лактата = 2 молекулы пропионата + 1 молекула ацетата + 1 молекула С02 + Н20. На практике соотношение между пропионатом и ацетатом изменяется в зависимости от воздействия многих факторов. Увеличение аспарагина ведет к росту количества сукцината, СО2 и количества ацетата [28, 112]. Независимо от способности формировать аспартат сукцината аминокислоты аспарагин, аланин, серии и глицин могут тоже расщепляться. Это так называемый аминокислотный метаболизм. Таким образом, пропионаты и ацетаты формируются в соотношении 1: 1. Метаболизм аминокислот независимо от протеолиза связан и оказывает действие на ферментацию пропионовокислых бактерий и, исходя из этого, заметно влияет на образование аромата [28, 77]. Штамм Propionibacterium freudenreichii связан с формированием типичного сладковатого вкусоследствия и образования свободного пролина, который получается в результате протеолиза казеина, при воздействии фермента пролинаминопептидазы [25, 154]. |
Последнее изменение этой страницы: 2017-04-13; Просмотров: 726; Нарушение авторского права страницы