|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Нерастворимые в воде и водно-солевых растворах
- Белки опорных тканей: Коллаген – это общее название большого семейства похожих протеинов, которые являются основными структурными компонентами межклеточного матрикса многих тканей. На их долю приходится > 30% всех белков (кожи, сухожилий, кровеносных сосудов, хрящей, зубов). Естественно, что в различных тканях регистрируются разные типы коллагенов. В настоящее время описано 19 типов, различающихся первичной структурой полипептидных цепей, функциями и локализацией в организме. Данный вид белков отличают следующие особенности состава и строения. 1. Полипептидная (п/п) цепь коллагена содержит около 1000 а/к остатков, чаще всего включает глицин, пролин и гидроксипролин, гидроксилизин. 2. Первичная структура коллагена – это три α -цепи с повторяющимися участками … –гли-про-Х-…, где Х – обычно аланин, нередко лизин или гидрокси пролин. 3. При формировании вторичной структуры нить коллагена укладывается в левозакрученную α -спираль, на один виток которой приходится 3 а/к остатка. Шаг спирали составляет 0, 95 нм (почти вдвое больше обычной), поэтому она сильно растянута. 4. Все три спирализованные α –цепи перекручиваются относительно друг друга, образуя правозакрученную тройную суперспираль – тропоколлаген , удерживаемую с помощью водородных связей. Пролин и гидроксипролин обеспечивают стабильность структуры, ограничивая вращение вокруг цепей. Глицин, не имеющий радикала, находится внутри тропоколлагена, что позволяет частям плотно прилегать друг к другу. В результате такого скручивания образуются сшивки между молекулами тропоколлагена, создавая коллагеновые фибриллы. Плотная структура тройной спирали и обилие поперечных связей между субъединицами делает молекулу практически нерастижимой и очень упругой.
Это основной белок эластических волокон, которые в больших количествах содержатся в межклеточном веществе кожи, стенок кровеносных сосудов, связок, лёгких. Название этих фибрилл отражает их свойство - эластичность, т.е. способность возвращаться в исходное состояние после деформации, что обусловлено особенностями состава и строения белка. Его полипептидные цепи сохраняют гибкую случайную конформацию благодаря наличию гидрофобных радикалов и пролина. А главное – в эластине возникают совершенно уникальные сшивки, присущие только этому белку за счёт формирования таких структур, как лизиннорлейцин и десмозин, образованные соответственно 2-мя или 4-мя остатками лизина разных цепей; Ретикулин отличается от коллагена наличием в своём составе серосодержащих аминокислот, которые образуя дисульфидные связи между цепочками, делают белок ещё более прочным. - Белки покровных тканей: Кератин - это протеин волос, ногтей, а также когтей, шерсти, меха. По строению его полипептидные цепи, образующие β – складчатый слой, включают до 12% серосодержащих аминокислот, с помощью которых отдельные нити прочно связываются между собой что обуславливает, например, форму волос (прямые или кудрявые), а отсюда и возможность изменения их структуры (химическая завивка). Сложные белки Это структуры, содержащие белковую часть – апопротеин и небелковый компонент – простетическую группу. В зависимости от строения последней принято различать следующие сложные белки. Фосфопротеины – это сложные белки, простетической группой которых является остаток фосфорной кислоты . Тип связи между апопротеином и небелковым компонентом – сложноэфирная, которая образуется при взаимодействии ОН-группы серина или треонина с фосфорной кислотой. К протеинам этого класса относятся казеиноген молока, фосфорилированные модификации гистонов, ферменты (РНК-полимеразы, некоторые фосфотрансферазы, фосфатазы) и другие, а также овальбумин белка и вителлин, фосвитин желтка яиц, ихтуллин икры рыб. Остатки фосфорной кислоты, включённые в белки, находятся в диссоциированом состоянии, что придаёт молекуле сильный отрицательный заряд. Поэтому фосфопротеины могут легко взаимодействовать с лигандами или отталкивать их. Так, например, они связывают ионы Са2+ и транспортируют их. Но главная особенность данной простетической группы – это возможность изменения конформации и заряда белковой молекулы, что сказывается на её свойствах и функциях. Реакция взаимодействия с фосфорной кислотой называется фосфорилирование или дефосфорилирование и используется, например, для регулирования активности ферментов и изменения функциональной активности биомембран (проницаемости и др.). Фосфопротеины необходимы для развития зародыша, плода, новорожденного, т.к. содержат оптимальный набор аминокислот, а фосфорная кислота используется для формирования скелета (связывание ионов Са2+), для образования макроэргов – АТФ. АДФ и др., которые требуются в качестве источников энергии в реакциях синтеза, особенно интенсивно протекающих в растущем организме. Нуклеопротеины – сложные белки, простетической группой которых являются нуклеотиды, и в первую очередь нуклеиновые кислоты – ДНК и РНК. В качестве апопротеина выступают белки гистоны, реже протамины. Так как эти протеины положительно заряжены, а нуклеиновые кислоты – отрицательно, то связь, образуемая между ними, ионная. Множество октамеров гистонов, комплексируясь с полинуклеотидом, образует нуклеосомы, которые затем компактно укладываются в органоидах (рис. ). В функциональном отношении они отвечают за хранение и передачу наследственной информации, и процессы биосинтеза белка. Хромопротеины («цветные белки») своей окраской обязаны простетической группе – пигменту. В зависимости от строения различают следующие подклассы: гемопротеины, флавопротеины, родопсин. Гемопротеины (красные) – сложные белки, простетической группой которых служит гем. Он представлен порфирином, состоящим из 4-х пиррольных колец, соединённых метеновыми мостиками. В центре располагается атом Fe2+ (гем b) или Mg2+ (гем а). Связь между апопротеином и небелковым компонентом координационная (донорно-акцепторная). Гемопротеины различаются по составу и структуре белка, обеспечивая разнообразие биофункций. Чаще всего встречается гем b (рис. ). Он входит в состав гемоглобина (Hb) и миоглобина, обеспечивающих связывание газов, транспорт их эритроцитами или накопление кислорода миоцитами. Мицелла Hb состоит из четырёх полипептидных цепей и служит примером обладания биологическим свойством кооперативность (см. выше) – то есть у каждой последующей субъединицы сродство к кислороду выше предыдущей. Гемоглобин присоединяет кислород в условиях его высокого парциального давления и отдает соответственно в среде с низким содержанием этого газа. Сродство гемоглобина к монооксиду углерода больше, чем к кислороду, поэтому угарный газ (СО) может вытеснять кислород из оксигемоглобина. Образующийся карбоксигемоглобин не способен служить переносчиком кислорода, в тканях развивается гипоксия, опасная для клеток. Карбоксигемоглобин имеет вишнево-красную окраску, обусловливающую цвет лица людей, отравившихся диоксидом углерода, что позволяет легко диагностировать токсическое действие газа. Зародышевый гемоглобин F обладает более высоким сродством к кислороду, чем HbА взрослых людей. Благодаря этому возможен оптимальный перенос газа от матери к плоду. Миоглобин – небольшой глобулярный белок (Mr 16500 Да); молекула его состоит из одной полипептидной цепи (153 а/к) и одного гема. Сродство к кислороду у данного гемопротеида значительно больше, чем у гемоглобина, поэтому он может принимать газ от последнего для сохранения или использования его в мышечных клетках. Гем b является также структурной единицей ферментов каталазы и пероксидазы, обезвреживающих пероксиды, и цитохромов – митохондриальных белков, участвующих в переносе электронов от окисляемых веществ в конечном итоге на кислород за счёт своей способности изменять валентность железа. Гем а встречается в растительных гемопротеинах и участвует в процессе фотосинтеза. Флавопротеины ( flavus - жёлтый). Их простетической группой являются активные формы витамина В2 (рибофлавин) – ФАД (флавинадениндинуклеотид), ФМН (флавинмононуклеотид). Классический пример данного вида белков оксидоредуктазы — ферменты, катализирующие окислительно-восстановительные реакции в клетке. В некоторые флавоноиды включены ионы металлов металлофлавопротеины и молекула гема – флавогемопротеины. Их типичными представителями, содержащими также негемовое железо, являются ксантинДГ, альдегидоксидаза, дигидрооротатДГ, ацил-КоА-ДГ. На долю последних приходится до 80 % митохондриальных белков, выполняющих важную роль в биоэнергетике клетки. Родопсин (от греч. rhodos — пурпур и opsis — зрение) – это белок, простетической группой которого служит активная форма витамина А – ретиналь. Апопротеин связан с ним иминной связью. Родопсин - основное светочувствительное вещество палочек сетчатки глаза. Его функция состоит в восприятии света в сумерках, т.е. отвечает за сумеречное зрение. Металлопротеины – сложные белки, где роль небелкового компонента выполняют ионы металлов. Связь между ними ионная или координационная (донорно-акцепторная). Типичными представителями таких белков являются железосодержащие белки. Ферритин. В его составе ионы Fe3+ депонируются в клетках селезёнки, костного мозга, но богаче всего этим белком печень. Там может накапливаться до 700 мг железа, которое используется по мере надобности для синтеза гема. Трансферрин – растворимый в воде железопротеин, содержащий ещё олигосахариды (металлогликопротеин). Этот белок вырабатывается гепатоцитами и секретируется в кровь, где служит для транспорта катионов этого металла к клеткам других органов. Молекула трансферрина удерживает два иона железа, которые захватывает в кишечнике или местах распада гема, и осуществляет их перенос к местам депонирования и утилизации. Гемосидерин – это комплекс гликопротеина и ионов железа, необходимый для кумулирования избытка железа. Церулоплазмин аналогично трансферрину обеспечивает транспорт катионов металла, только восьми ионов меди. Кроме того, многие ферменты в свой активный центр включают различные ионы металлов, выступающие в роли Ко-фактора: Zn2+ - для алкогольдегидрогеназы (алкогольДГ), Mn2+ - для аргиназы, Mg2+ - для гексокиназы, Мо2+ - для ксантинДГ и другие Гликопротеины или гликоконъюгаты. В них простетическая группа представлена углеводными компонентами и связана с белком О-гликозидными (иногда N-гликозидными) связями. Небелковый компонент некоторых гликопротеидов редко бывает представлен одним моносахаридом; как правило, это олиго- или полисахаридные разветвлённые цепочки. Причём на долю углеводов может приходиться от 1 до 85%. Исходя из вклада в общую копилку, выделяют два подкласса белков, содержащих углеводы: протеогликаны и гликопротеины. Между ними имеются существенные отличия:
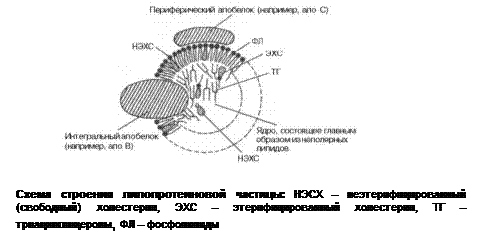
Другая группа гликоконъюгатов – протеогликаны – характеризуется Липопротеины – сложные белки, простетическая группа в которых – липиды: нейтральные жиры (ТАГ), эфиры холестерола, ФЛ. Между компонентами возникает гидрофобное взаимодействие, реже ионная или эфирная связи. Все липопротеины имеют сходное строение (рис ).
Внутри липопротеиновой частицы находится жировая капля – ядро, содержащее неполярные липиды: ТАГ и эфиры холестерола. Снаружи оно окружено однослойной мембраной, образованной ФЛ, белком и НЭХС. Белковый компонент получил название аполипопротеин(ы), некоторые из них прочно связаны со всеми фрагментами и не могут быть отделены от целой мицеллы, а другие способны переноситься от одного липопротеина к другому. Их белки выполняют несколько функций: формируют структуру частицы, взаимодействуют с рецепторами на поверхности плазмоллем, определяя, каким тканям он необходим, служат ферментами или их активаторами, действующими на липопротеин. Плотность белка выше плотности липидов, поэтому, чем меньше доля апопротеина в частице, тем больше её плотность. По соотношению входящих в частицу компонентов различают несколько классов липопротеинов: хиломикроны (ХМ), липопротеины очень низкой плотности (ЛПОНП), липопротеины промежуточной плотности (ЛППП), липопротеины низкой плотности (ЛПНП), липопротеины высокой плотности (ЛПВП). Каждый из типов ЛП образуется в разных тканях и транспортирует определённые липиды. Благодаря наличию отрицательного заряда на поверхности ЛП хорошо растворимы в крови. Мицеллы небольших размеров способны легко проникать через стенки сосудов, а некоторым это делать трудно (ХМ) из-за больших размеров, поэтому они сначала попадают в лимфатические сосуды, затем через главный грудной проток вливаются в систему полых вен, правое предсердие, малый круг кровообращения и только потом оказываются в аорте и артериях, доставляющих их к органам. Характеристика ЛП частиц представлена в таблице 1. Таблица 1. |
Последнее изменение этой страницы: 2017-05-11; Просмотров: 205; Нарушение авторского права страницы
 Эластин. В отличие от коллагена он включает около 800 а/к остатков, среди которых преобладают лизин, пролин, а также глицин, валин, аланин.
Эластин. В отличие от коллагена он включает около 800 а/к остатков, среди которых преобладают лизин, пролин, а также глицин, валин, аланин. 
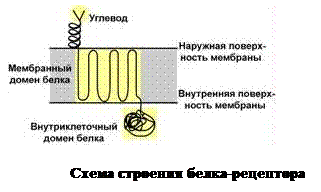 Каквидно из таблицы, для собственно гликопротеинов характерно низкое содержание углеводов 15-20 %, которые присоединены к амидному азоту аспарагина, либо к гидроксигруппе остатка серина, реже треонина, гидроксилизина. Вглициде отсутствует регулярность в чередовании моносахаридов, регистрируются манноза, галактоза, глюкоза, их аминопроизводные, N-ацетилнейраминовая кислота. Разнообразие гликановых фрагментов используется при построении мембранных рецепторов (рис. ), для обеспечения взаимодействия антиген-антитело, специфичных контактов клеток между собой, а также в защите ферментов от протеолиза. Почти все внеклеточные белки, включая глобулины крови, относятся к гликопротеинам. Некоторые гормоны (ТТГ- тиреотропный, ГТГ- гонадотропные) являются гликопротеинами. К типичным представителям данного класса белков относятся все антитела (иммуноглобулины), интерфероны, факторы комплемента, группы крови и др. Первые — синтезируются в плазмоцитах, обезвреживают антигены любой химической природы. Различают пять классов иммуноглобулинов: IgG, IgM, IgA, IgD и IgE. Интерфероны (α, β, γ ) — гликопротеины, образующиеся в клетке в ответ на внедрение вирусов и ингибирующие их размножение.
Каквидно из таблицы, для собственно гликопротеинов характерно низкое содержание углеводов 15-20 %, которые присоединены к амидному азоту аспарагина, либо к гидроксигруппе остатка серина, реже треонина, гидроксилизина. Вглициде отсутствует регулярность в чередовании моносахаридов, регистрируются манноза, галактоза, глюкоза, их аминопроизводные, N-ацетилнейраминовая кислота. Разнообразие гликановых фрагментов используется при построении мембранных рецепторов (рис. ), для обеспечения взаимодействия антиген-антитело, специфичных контактов клеток между собой, а также в защите ферментов от протеолиза. Почти все внеклеточные белки, включая глобулины крови, относятся к гликопротеинам. Некоторые гормоны (ТТГ- тиреотропный, ГТГ- гонадотропные) являются гликопротеинами. К типичным представителям данного класса белков относятся все антитела (иммуноглобулины), интерфероны, факторы комплемента, группы крови и др. Первые — синтезируются в плазмоцитах, обезвреживают антигены любой химической природы. Различают пять классов иммуноглобулинов: IgG, IgM, IgA, IgD и IgE. Интерфероны (α, β, γ ) — гликопротеины, образующиеся в клетке в ответ на внедрение вирусов и ингибирующие их размножение. наличием крупных полисахаридов (рис. ), в состав которых входят гликозаминогликаны (ГАГ): гиалуроновая кислота, хондроитин-сульфат, кератансульфаты, дерматансульфаты и гепарансульфаты. Углеводные фрагменты усиливают гидрофильные свойства белка за счёт большого количества ОН-групп и кислотных группировок. Соотношение компонентов: протеина 10-15%, а углеводов 90-95%. Цепи последних недостаточно гибкие и стремятся принять конформацию очень рыхлого случайного клубка, занимая огромный объём. Будучи гидрофильными, они притягивают много воды и даже в низких концентрациях образуют гидратированные гели. Подобная способность создаёт во внеклеточном пространстве – тургор. Протеогликаны хрящевого матрикса содержат гиалуроновую кислоту, образующую студенистый гель, выполняющий роль амортизатора в хрящах и суставных поверхностях.
наличием крупных полисахаридов (рис. ), в состав которых входят гликозаминогликаны (ГАГ): гиалуроновая кислота, хондроитин-сульфат, кератансульфаты, дерматансульфаты и гепарансульфаты. Углеводные фрагменты усиливают гидрофильные свойства белка за счёт большого количества ОН-групп и кислотных группировок. Соотношение компонентов: протеина 10-15%, а углеводов 90-95%. Цепи последних недостаточно гибкие и стремятся принять конформацию очень рыхлого случайного клубка, занимая огромный объём. Будучи гидрофильными, они притягивают много воды и даже в низких концентрациях образуют гидратированные гели. Подобная способность создаёт во внеклеточном пространстве – тургор. Протеогликаны хрящевого матрикса содержат гиалуроновую кислоту, образующую студенистый гель, выполняющий роль амортизатора в хрящах и суставных поверхностях.