|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Регуляция каталитической активности ферментов белок-белковыми взаимодействиями
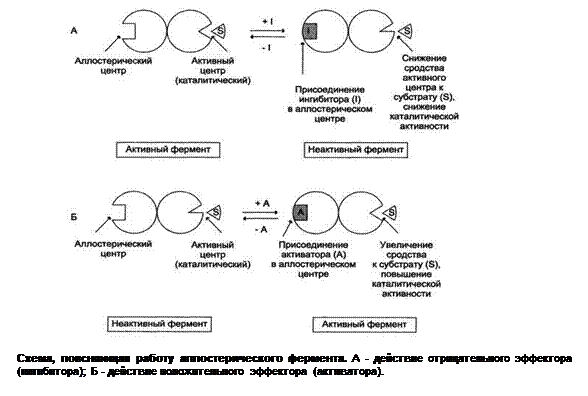
Описано два механизма активации ферментов с помощью белок-белковых взаимодействий: · в результате присоединения регуляторных белков; · вследствие ассоциации или диссоциации энзима, обладающего четвертичной структурой: Схема изложения регуляции активности Взаимодействие с субстратом, ферментом, энзим-субстратным комплексом, но выгоднее с биокатализатором, так как его концентрация невелика, значит, не требуется большого количества эффектора. Чаще с аллостерическим центром реагирует аллостерический фермент.
Регуляция путём ковалентной модификации В биологических системах часто встречается механизм регуляции активности энзимов с помощью ковалентной модификации в нихаминокислотных остатков. Быстрый и широко распространённый способ подобного явления - фосфорилирование/дефосфорилирование. Модификации подвергаются ОН-группы серина активного центра фермента. Фосфорилирование тоже требует работы биокатализаторов и осуществляется ферментами протеинкиназами, а дефосфорилирование - фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической способности. При этом результат может быть двояким: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными (рис. 5).
Изменение активности фермента, вызванное фосфорилированием, обратимо. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять направленность хода процесса в зависимости от возникающих условий. Антагонистичные по функции гормоны по-разному влияют на фосфорилирование/дефосфорилирование ферментов, вызывая противоположные эффекты в метаболизме. Например, под действием глюкагона (в период между приёмами пищи) в клетках замедляется синтез энергетического материала (жиров, гликогена), но усиливает их распад (мобилизация), спровоцированные фосфорилированием ключевых ферментов этих процессов. Под действием же инсулина (во время пищеварения), наоборот, активируется образование гликогена, за счёт происходящего дефосфорилирования тех же энзимов. 2.2.5. Частичный протеолиз – как способ активации энзима (активация зимогена) Зимоген, или профермент – это неактивная форма фермента. Когда активный центр энзима закрыт олигопептидным фрагментом белковой цепи, субстрат не может с ним взаимодействовать. Некоторые биокатализаторы, функционирующие вне клеток (в полости ЖКТ или в плазме крови), синтезируются в виде неактивных предшественников и их преобразование осуществляется с помощью гидролиза одной или нескольких определённых пептидных связей, что приводит к отщеплению участка белковой молекулы предшественника. В результате в оставшейся части апофермента происходит конформационная перестройка и открывается его активный центр. Рассмотрим механизм на примере активации протеолитического энзима трипсина. Трипсиноген, зимоген данного фермента синтезируемый в поджелудочной железе, по её протокам поступает в двенадцатиперстную кишку, где и активируется выше упомянутым способом под действием фермента кишечника энтеропептидазы, отщепляющей гексапептид с N-конца молекулы, в результате формируется активный центр в сохранившейся части мицеллы. Частичный протеолиз - пример регуляции, когда активность биокатализатора изменяется необратимо. Такие ферменты функционируют, как правило, в течение короткого времени, определяемого сроком жизни белковой молекулы. Данный способ регуляции лежит в основе активации не только протеаз, но и белков свёртывающей системы крови и фибринолиза, протеинов системы комплемента, а также пептидных гормонов. Аллостерическими эффекторами часто служат различные метаболиты. Конечные продукты метаболического пути - часто ингибиторы аллостерических ферментов, а исходные вещества - активаторы. Это так называемая гетеротропная регуляция. Такой вид аллостерической регуляции очень распространён в биологических системах.
Ингибирование ферментов Скорость ферментативной реакции, как и активность энзима, в значительной степени определяется также присутствием в среде ингибиторов, которые снижают её. Ингибиторы биокатализаторов – это вещества, вызывающие частичное (обратимое) или полное торможение процессов, катализируемых ферментами. В зависимости от прочности связи, образующейся между эффектором и энзимом, принято выделять два вида ингибирования: обратимое и необратимое. Если ингибитор связывается с ферментом ковалентной связью, вызывая стойкие изменения пространственной структуры молекулы энзима или модификацию функциональных групп, такое явление называется необратимым. Его примером может служить результат действия йодацетата, диизопропилфторфосфата (ДФФ), а также диэтил-n-нитрофенилфосфата и солей синильной кислоты, что приводит к cвязыванию и выключению функциональных групп или ионов металлов в молекуле фермента. Так, например, ДФФ – соединение из группы нервнопаралитических отравляющих веществ – прикрепляясь к остатку аминокислоты серина, находящегося в активном центре энзима ацетилхолинэстеразы, образует фермент-ингибиторный комплекс. В норме данный биокатализатор гидролизует ацетилхолин, играющий роль нейромедиатора. Почти сразу после очередного нервного импульса ацетилхолинэстераза расщепляет его на холин и уксусную кислоту и нейрон готов к передаче следующего импульса. Если этот фермент ингибирован, то ацетилхолин накапливается, нервные импульсы следуют один за другим и мышца длительное время не расслабляется. В конце концов, наступает паралич или смерть. Некоторые биокатализаторы полностью угнетаются очень малыми концентрациями ионов тяжелых металлов, например ионов ртути (Hg2+), серебра(Ag+) и мышьяка (As+) или йодуксусной кислотой. Эти электролиты необратимо соединяются с сульфгидрильными группами (–SH) и вызывают денатурацию апофермента. В свою очередь, в зависимости от типа локуса фермента, куда садится эффектор, различают конкурентное и неконкурентное ингибирование. В случае обратимого действия ингибитор образует с энзимом непрочный комплекс, способный распадаться, в результате активность биокатализатора восстанавливается. Первое явлениеможет быть вызвано веществами, имеющими структуру, сходную со строением молекулы, но несколько от него отличающуюся. Такое ингибирование основано на связывании ингибитора с активным центром биокатализатора, что нашло широкое применение в медицинской практике. Известно, например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяют сульфаниламидные препараты. Оказалось, что эти средства имеют структурное сходство с парааминобензойной кислотой, которую бактериальная клетка использует для синтеза жизненно важной фолиевой кислоты, являющейся коферментом. Благодаря близости пространственного строения сульфаниламид блокирует действие фермента путем вытеснения парааминобензойной кислоты из комплекса с ним, что ведет к торможению роста и размножения микроорганизмов. Неконкурентные ингибиторы это аллостерические регуляторы. Фермент, катализирующий превращение субстрата А в продукт В, имеет аллостерический центр для отрицательного эффектора, которым служит конечный продукт метаболического пути F. Если концентрация F увеличивается (т.е. вещество F синтезируется быстрее, чем расходуется), ингибируется активность одного из начальных ферментов. Такую регуляцию называют отрицательной обратной связью, или ретроингибированием. Отрицательная обратная связь - часто встречающийся механизм регуляции метаболизма в клетке. Такая регуляция представляется логичной, так как при накоплении конечного продукта он (конечный продукт – F1) может действовать как аллостерический центр фермента Е1, катализирующего чаще всего начальный этап данного метаболического пути:
В качестве примера можно рассмотреть регуляцию начального пути распада глюкозы. Один из конечных продуктов её распада - молекула АТФ. При накоплении данного вещества в клетке происходит ретроингибирование аллостерических ферментов фосфофруктокиназы и пируваткиназы.
|
Последнее изменение этой страницы: 2017-05-11; Просмотров: 263; Нарушение авторского права страницы