|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Химическое и биохимическое доказательство
Представь себе образ прекрасной и совершенной Вселенной… — А затем поверь только в одно: Абсолют уже создал ее в Своем Воображении все-таки немного лучше, чем это сделал ты… Ричард Бах
Рассказывая выше о строении аминокислот, я не упомянул еще об одном важном обстоятельстве. Дело в том, что многие вещества обладают оптической активностью. Атом углерода, являющийся основным «строительным материалом» органических молекул, соединяется с атомами других элементов вдоль четырех пространственных осей, направленных из центра воображаемого тетраэдра к его вершинам. В таких молекулах один или несколько атомов углерода оказываются связанными с четырьмя разными заместителями. Это приводит к тому, что возникает так называемая оптическая изомерия (рис.49). Состав двух молекул может быть одинаков {содержание углерода, кислорода, азота, серы…), строение одинаково (как и в каком порядке они соединены друг с другом), но в пространстве, если бы мы взяли в руки их модели, они не совмещаются, как бы их ни передвигали. Не совмещаются так же, как рил органических молекул не совмещаются кисти правой и левой руки. Если поставить, например, кювету13 с раствором только одного из наших изомеров на пути поляризованного света14, то свет, пройдя такое препятствие, изменит плоскость поляризации. Она будет проходить с отклонением на некоторый угол от первоначального. Знак и степень такого отклонения, замеренная в стандартных условиях, является количественным выражением степени оптической активности. Молекулы оптических изомеров различаются индексами и поэтому дальше по тексту аминокислоты будут иногда, когда это необходимо, обозначаться с индексом, например: L-npo-лин или D-аланин. При написании без индекса будет подразумеваться L-аминокислота.
Рис.49. Оптическая изомерия органических молекул Принципиальный факт биохимии живого состоит в том, что все двадцать «белковых» мономеров это именно L-аминокислоты, Вообще говоря, если сравнить эти формы одного и того же вещества в чистом виде, то обнаруживается не только биохимическое различие. Оно проявляется в небольшом, но все же важном отличии большинства физических и химических свойств (температуры плавления, растворимости, скорости реакций,,, )* Обратите внимание, что одна из аминокислот — пролин {Pro} — это не а-аминокислота, а циклическая аминокислота. Это тем более заставляет меня удивиться гармонии истинно биохимической (не химической только! ) естественной системы, в которой этот особенный элемент четко вписывается в нумерологическую последовательность и, соответственно, находит свое точное место. Выше, начиная свой рассказ об аминокислотах, я сравнивал их с древнекитайским символом «тайцзи». Я написал, что каждая из них построена, образно говоря, как «противоположение в одном», то есть представляет собой слияние воедино двух противоположных химических начал. Но, оказывается, и в структуре системы аминокислот, как и в структуре алфавита, тоже представлены два начала — есть линия аланина, или идущая от А, и есть линия глицина, или идущая от О. Отсюда видно, что общий закон природы — закон аналогии — проявляется не только в единой структуре (и, по-видимому, природе) разных систем, (аминокислот и алфавита), но и в аналогии структуры элемента и системы, эти элементы объединяющей. Элемент (аминокислота) имеет единую, но противоположную («двуначальную») природу, и система аминокислот единая, цельная, но также «двуначальная». Ну а теперь перейдем непосредственно к рассмотрению свойств аминокислот. Начнем по порядку. Сначала рассмотрим изменение количественных свойств графически, а полученные при этом выводы (если таковые появятся) попытаемся подтвердить разбором всех имеющихся до сих пор качественных классификаций аминокислот. Дело это непростое, требует соответствующего языка, поэтому тот, кто начнет путаться уже на втором абзаце, может просто взглянуть на соответствующие кривые графиков. Они скажут сами за себя без длинных доказательств. Поскольку одним из ключей к предположению о системе, существующей во множестве двадцати мономеров белка, была мысль о важной роли порядковых номеров молекул (точнее, их числовых кодов), в первую очередь возникает необходимость проверить, как в такой системе меняются молекулярные массы. Это будет, если угодно, своеобразной данью уважения И. Ньютону и Д.И. Менделееву. На горизонтальной оси встречно и последовательно расположим системные периоды аланина и глицина, пробелом отделив вторые ряды, начинающиеся с проли-на и изолейцина (рис.50).
Рис, 50. Изменение молекулярной массы в естественных периодах Аминокислот
Построив график, обнаруживаем четкое разделение на эти ряды. При этом в каждом из них обнаруживается плавный закономерный рост молекулярных масс. Разумеется, последовательно расположенные пары {глутаминовая кислота (147, 1) —глутамин (146, 2)} и {аспаргиновая кислота (133, 1) —аспаргин (132, 1)} дают на графиках участки некоторого уменьшения массы. Однако совокупность других свойств в сравнении этих пар требует оставить их в системе именно в таком порядке, обратном с точки зрения общего характера изменения массы. Если учесть заниженное значение массы молекулы пролина, объясняемое его особым качеством, то ход кривых на закономерно противопоставленных в нашей системе периодах аминокислот обнаруживает четкую качественную симметрию. Эта симметрия наглядно проявляет себя в зеркальном отражении, во-первых, последовательности чередования участков более плавных и более резких изменений, а во-вторых, в расположении точек перегиба кривых. Отметим здесь, что под качественной симметрией кривых на наших графиках будем далее понимать, кроме перечисленного, вообще, всякое геометрическое подобие кривых, разумеется, с соблюдением требования «однофазности колебаний», Следующим по очереди разберем кривые изменения радиусов и объемов радикалов аминокислот [56, с.47] (рис.51). Мы обнаружим четкую осевую симметрию подобия и четкое различие характера графиков по рядам каждого из периодов (правого и левого).
Рис.1.График изменения характеристик радикалов аминокислот
Более сложные кривые изменения констант кислотности и основности аминокислот [50, с. 11-38] (рис.52, 53), однако, если учесть характерное «выпадение» свойств лролина, то можно все-таки обнаружить подобие левой и правой частей графиков, особенно для первых рядов обоих периодов: {Ala, Thr, Leu, Met, Trp} и (His, Cys, Val, Ser, Gly}.
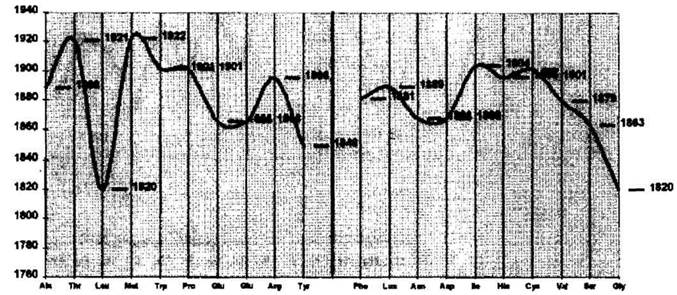
Рис.53. Кривые изменения констант основности в системе аминокислот Графики растворимости аминокислот в воде [там же] отражают уже замеченные системные закономерности (рис.54). Весьма интересным представляется ход кривых удельного оптического вращения аминокислот (рис.55) [там же]. В этом случае функциональная зависимость выражает четкую зеркальную симметрию подобия, а также различный характер графиков по двум рядам каждого периода. м „.
Рис, 54. График растворимости аминокислот в воде
Рис.55. График изменения удельного оптического вращения растворов аминокислот
Когда аминокислоты хроматографически15 отделяют друг от друга, все их физические и химические свойства оказываются интегрально выражены одним простым числом — номером в том порядке, в котором каждая из них появляется в виде «пятна» на специальной пластинке [42, с-136]. Отложим эти числа на уже знакомой системе координат {рис.56). Уже не удивительно, что и здесь налицо осевая симметрия подобия.
Рис.56. Графическое выражение порядка выхода аминокислот при хроматографии Более того, если рассмотреть известные значения изоэлектрической точки аминокислот16 [61, с. 106] и построить соответствующий график, можно обнаружить, что кривые на этом построении имеют совершенно особый вид (рис.57). Идеальную гармонию правой части графика несколько нарушает заниженное значение изоэлектрической точки у триптофана. Если скорректировать это значение (а также незначительно изменить некоторые другие) и построить соответствующий гипотетический график, то кривые будут полностью гармоничны — они точно воспроизведут известную в физике каноническую форму колебаний при наличии трения, затухающих по амплитуде с уменьшением частоты (рис, 58). Эти кривые, естественно, можно описать соответствующими математически уравнениями. Симметрия приведенных кривых, две генетические линии в системе, очень наглядно проявляют себя при построении графиков в круговых координатах (рис.59).
Рис.57, График иэоэлектрической точки аминокислот
Рис.58. Канонический характер графика изменения изолектрической точки аминокислот
Рис.59. График изоэлектрической точки аминокислот в круговой системе координат
Отметим, что если все остальные построенные mною графики в большей или в меньшей степени подтверждают обоснованность разделения каждого {правого и левого) периода на два ряда, то эти кривые, кривы изоэлектрической точки, свойства, которое одним число! синтетически выражает кислотно-основную двойственность аминокислот, наоборот, соединяют периоды, показывают их глубинное внутреннее единство. Как тут еще раз не вспомнить об эволюции молекул? Можно представить себе, что по мере движения вдоль естественной последовательности аминокислот, каждая из этих двух кривых показывает ускоренный, приближенный к экспоненциальному, рост разнообразия качеств; заключенного в амплитуде колебаний от кислотности к основности. И когда эти колебания достигают своего предельно возможного размаха — система заканчивается. Разумеется, сегодня я далек от того, чтобы сравнивал природу этих кривых — пока речь идет лишь о форме. Хотя за подобной «изоморфностью» физических колебаний и колебаний электрических свойств аминокислот наверняка стоит некий единый закон. Анализируя характер полученных кривых, я имею возможность сделать два принципиальных вывода. Они тем более важны, если учесть, что вначале наших рассуждений идея системы основывалась на порядке аминокислот по уменьшению кода шрядковых номеров, на изменении состава, строении и усложнении химического качества радикала. Нелишне будет заметить, что сама концепция естественной научной классификации (по А.А. Лю-бищеву) требует проявления основных закономерностей на всей совокупности основных свойств. Первый вывод заключается в том, что симметричный характер кривых изменения радиуса и объема радикалов аминокислот, растворимости, оптического вращения, очередности выхода «пятен» при хроматографии, % в белках (рис.60, 61) подтверждает фундаментальную общую структуру открытой системы —два начала и два конца системы, две генетические линии связи, разделение каждого периода из десяти элементов на два ряда по пять. Плавность хода кривой на графике молекулярных масс, а также гармонический закономерный характер изменения изоэлектрической точки аминокислот позволяют: однозначно подтвердить правильность первоначально принятой последовательности аминокислот в рядах. Появляются веские основания для того, чтобы решить две (уже упомянутые ранее) проблемы нашей классификации. Во-первых, пара {аргинин-лизин} располагается девятой, тогда как первоначально были предположены три варианта ее места (седьмой, девятой или десятой по счету). Во-вторых, принятое чередование пар (сначала— {глу-таминовая кислота — аспаргиновая кислота}, и затем — {глутамин-аспаргин}) оказывается обоснованным. Сведущий читатель уже заметил, что в каждом из приводимых графиков можно условно выделить свои «периоды», я же использую это понятие для закономерно выделенной десятки аминокислот.
Рис.60. График встречаемости аминокислот в белках бактерий
Рис.61. График встречаемости аминокислот в белках человека
Конечно, это разные периоды, а такое название — дань традиции, ведь исследование системы аминокислот для меня неразрывно связано с исследованием периодической системы Д, И. Менделеева. Да и сами графики, построенные мной для доказательства системы аминокислот восходят к двум удивительным зеркальным кривым (рис.62 и 63) [24, с.96 и 102], которые я обнаружил когда-то, доказывая справедливость своей спиральной симметричной системы химических элементов (см.рис.4). Интересным историческим и науковедческим подтверждением симметрии разделения аминокислот на две серии по всей совокупности их свойств (составу, строению, физико-химической и биохимической индивидуальности) является моя попытка (забавы ради! ) отложить на системной «встречной» оси годы обнаружения этих соединений [56, с.45] за сто лет (рис.64). Конечно, в данном случае качественная осевая симметрия подобия в основном проявлена в одинаковой «фазности» колебаний этой коивойт однако отдельные участки демонстрируют более полную симметрию. Действительно, аминокислоты открывали тоже в соответствии с их положением в естественной системе. Вот уж действительно, моя едва рожденная система «уже прошла испытание временем! », правда, пока не будущим, а только прошлым...
Рис.62. График изменения первого ионизационного потенциала атомов характеристической оси спиральной системы химических элементов 87
Теперь, обнаружив количественное доказательство своей теории, я обращусь к качеству, к существующим классификациям аминокислот. О самих подходах к построению систем в химии и биологии я уже рассказал читателю выше. Здесь же и дальше мы вместе попытаемся (если удастся! ) связать найденную систему именно с классификациями. Коротко объяснить различие классификации и системы нетрудно —первая разделяет объект на множества по некоторым {пусть даже и существенным) признакам, а вторая — объединяет его с учетом большинства главных. Посмотрим, как на нашей системе выглядит одна из существующих классификаций — по заряженности и полярности боковых радикалов [11]. Конечно, у химика, видевшего саму систему с указанными графическими формулами {см. рис.42), уже нет сомнений, что в системе все окажется закономерным.
Рис.64. Годы открытия аминокислот
Тем не менее, покажем это (рис.65):
Рис.65, Классификация аминокислот ло характеру радикала, нанесенная на естественную систему
Стоит напомнить, что в первую группу (I) относят аминокислоты с радикалами ионного характера {как + так и -), во вторую (II) — полярного не ионного, а в третью (III) — неполярного. Но эти термины для тех, кто еще не забыл химию, а художник может просто внимательно посмотреть на естественную систему (см. рис.42), он и без подготовки увидит как гармонично перекликаются структуры—они похожие (например, {Ala (10)—Gly (1)t Glu (70)— Asp (7)}) и в то же время — разные в каждой паре. Он легко заметит, как эволюционируют молекулы, как в каждом периоде (с началом в аланине и в глицине) они дважды усложняются, с первой по пятую (с № 10 по № 50), а затем с шестой по десятую (с № 60 по № 100), как закономерно в их структуре появляется кислород, азот, сера, а затем гетероциклические и ароматические радикалы,
* * * Сейчас я перехожу к очень важной части доказательства структуры системы аминокислот, так как, если даже все оказывается химически выверенным, какое отношение это имеет к жизни, к белку, вообще к биологии? В.И. Вернадский очень точно писал об этом когда-то: «Наше знание химического состава однородной живой материи по крайней мере на одно столетие отстало от химии минерала. Работая с биохимическими объектами, чувствуешь, что в своем распоряжении имеешь только обрывки необходимых знаний, обрывки, правда, стоящие на уровне современной химии, иногда проникающие очень глубоко в познание частностей, но совершенно не дающие общего представления q химическом составе не только живой материи, но и отдельного организма» [8, с.63]. Это абсолютно верно и теперь, полвека спустя. Мы только-только начинаем понимать основы жизни, хотя и накопили огромное количество отрывочных сведений, уже достаточных для такого понимания. Обосновывая систему алфавита, я уже приводил графики частоты встречаемости букв в текстах. Состав и структура большинства белков известна, поэтому можно также отложить число аминокислот на уже знакомой читателю системной двойной оси. Кривые {см.рис.60), построенные по данным об относительной встречаемости аминокислот в белке Escherichia Coli [30, с.35], прекрасно подтвердили мою концепцию симметрии подобия двух ветвей, двух линий системы. К сожалению, авторы книг по биохимии, не представляя себе подлинной системности своего материала, приводят единые цифры по встречаемости глутамина и глута-миновой кислоты {также аспаргина и аспаргиновой кислоты). Думаю, что отныне такого больше не будет, ведь это химически близкие, но здесь {в системе аминокислот) принципиально разные особые элементы. Это элементы системы уже не только химической, но и мономеров белка — основы жизни. Поэтому такие единые цифры пришлось разделять (сделав соответствующие подсчеты) — в одном случае беря 40 % от приводимой суммы для аспаргина, а в другом — 30 % для глутамина. Для сравнения на этом же графике я отложил уже упомянутые в первой главе данные собственных подсчетов букв алфавита. Конечно, эти графики различны, но их объединяет одно — единая зеркальная симметрия подобия, обнаруженная в структуре азбуки и в системе аминокислот. Соответствующие данные имеются по составу белков человека [56, с.48] (см. рис.61). Итак, мы с вами, дорогой читатель, вместе прошли по ступенями гипотезы» открытия и доказательства. Этот процесс теперь можно выстроить в определенную логическую цепочку: — нумерологический подхдд — аминокислоты попарно выстроились в определенном порядке; — химический подход: а) качественный — карточки с графическими формулами — через строение аминокислот, качественный элементный и функциональный характер их радикалов подтвердил правильность гипотезы; б) количественный — графики всех существенных свойств аминокислот не только блестяще подтвердили гипотезу, но и позволили утверждать, что порядок следования аминокислот был установлен правильно; — биохимический подход, выразившийся в анализе состава белков, окончательно подтвердил правильность исходных посылок. Не забудем, что в самом начале наряду с нумерологией нас вдохновляла лишь прорисовывающаяся аналогия аминокислот и азбуки, фантастическая идея химической структуры угаритских клинописных букв- Сейчас можно с полным правом утверждать, расположив уга-ритский алфавит в клетках открытой мною системы азбуки {ничего в нем не передвигая (! ), ведь он сам и есть истинная праазбука), что учителя угаритян точно располагали буквы-аминокислоты, нам же с вами это расположение пришлось доказывать. Получилось, что ничего-то я и не открыл, а только переоткрыл (или приоткрыл!? ) уже известное. Но есть и одна особенность. Я не просто подтвердил принципиальный вывод (поначалу обескураживающий), уже сделанный многими современными учеными, о том, что древние знали больше нас. Я обнаружил, что эти знания они использовали как код для создания основ нашей культуры. И алфавит — пример этому. О форме системы Наступило время, когда древние символы знания должны превратиться в научные формулы... Агни-йога
Описать сложнейшую вновь открытую систему дело не простое, она требует точных научных понятий, и поэтому уважаемому читателю придется вместе с автором немного потрудиться. Алфавит (говоря «языком» аминокислот—«аланитрит» (аланин — треонин)), представленный в графической матричной форме (то есть в виде таблицы), имеет парадоксальные свойства. Его сущность, как я уже отмечал, это не одна генетическая линия развития элементов периодической системы химических элементов, и не разрастание многообразия по графу («кусту») в системах цветковых растений. Главное в системе алфавита — две генетические линии развития элементов (ниже по тексту — просто линии (см. рис.41). Повторюсь, что при этом десять элементов одной линии образуют период алфавита, а каждая из четырех «пятерок» букв образует ряд. Чтобы не вызвать недоумения не очень внимательного читателя, специально остановлюсь подробнее на различии понятий «последовательности» и «структуры». На протяжении всего исследования я отмечаю принципиальное значение порядка, то есть строгой последовательности букв алфавита. И, строя систему, я ни в коем случае не пытаюсь как-то его изменить или «подправить». Он был, есть и будет таким (в нем-то и заключен «ключ» к системе), но как порядок перечисления, последовательного прочтения букв. Главное, что я обнаруживаю порядок прочтения букв — еще не структура алфавита- Его истинная структура заключена в симметрии двух генетических линий, изображенных на рис.41 и 42, И говоря о структуре, мы должны перечислять, прочитывать буквы уже по-другому — вдоль каждой из этих линий. Итак, я кратко повторил то главное, что отличает открытую систему алфавита и аминокислот белка от всего, что известно сегодня, и их уникальность позволяет предположить, что главное заключается не только в открытии этих систем, но и самого принципа существования в мире систем подобного типа. Попарно взятые элементы каждой линии (например, элемент 4 — метионин (Г) и элемент 4' — цистеин (С)) похожи по своей химической сущности (наличие серы) так, что нет в системе в целом более близких друг к другу элементов, но при этом метионин и цистеин противопоставлены -*> ени разные и по молекулярной массе и по сложности радикала, они принадлежат к разным линиям. То есть мы обнаруживаем здесь необычный тип зеркальной симметрии. Это и не симметрия в чистом виде (например: правая и левая рука), и не антисимметрия17 (например, одинаковые предметы черного и белого цвета), это, скорее, зеркальная симметрия подобия. Такой вид симметрии невозможно рассматривать изолированно от той системы, в которой его применяют, и он означает, что симметричны элементы хоть и разные, но наиболее близкие друг другу именно в данной системе. До сих пор мы говорили о табличной форме. А как представить обнаруженную закономерность иным способом, попытаться графически или пространственно полнее выразить ее сущность? Разумеется, прежде всего проверим испытанный вариант — спираль Архимеда на плоскости. Поскольку в алфавите мы имеем две линии развития, то нам потребуются две спирали, имеющие разные начала и к тому же не пересекающиеся, поскольку в системе нет общих аминокислот (или букв), одновременно принадлежащих обеим линиям. При этом нам необходимо: — показать общность, единство обеих линий; — противопоставить две линии (отразить осевую симметрию подобия); — показать определенное периодическое повторение структуры и свойств элементов полупериодов (вторые пять элементов линии (И—К—Л—М—Н) подобны первым пяти (О—П—Р—С—Т)). Обратим внимание, что такое подобие наблюдается не только у полупериодов каждой отдельно взятой линии, но и у полупериодов разных линий (ряд И—К—Л—М—Н второго периода подобен ряду А—Б— В—Г—Д первого). Ясно, что в этом случае нам необходимо сразу выходить в объем и рассматривать двойную спираль (! ) — именно она (и только она) удовлетворит всему перечисленному набору условий. Оба витка этой объемной спирали должны быть закручены в одну сторону, поскольку в противном случае они должны пересекаться, а это означало бы наличие общих аминокислот, принадлежащих обеим линиям. В качестве Образующей для такой спирали можно взять конус или цилиндр (и тогда она будет похожа на знаменитую структуру ДНК). Право автора назвать свое открытие, но и его право — придать ему ту или иную форму. Форма расширяющейся по конусу двойной спирали (две конические винтовые линии или две конические гелиссы) показалась мне наиболее подходящей, поскольку такое расширение показывает различие двух рядов одного периода (или, что то же самое, двух полупериодов) (рис.66 и 67). Фактически каждый из данных рисунков представляет собой проекцию этой спирали на плоскость. Такой вид системы и будем считать наиболее точной плоскостной формой. Разумеется, в объеме симметрия двух спиралей будет сложной — зеркально-поворотной (также как у ДНК), то есть плоскость симметрии будет вращаться вокруг оси симметрии вместе со спиралями. Ну а на наших рисунках (проекция на плоскости) эта ось будет осью симметрии подобия системы алфавита (буквенного и аминокислотного). И, разумеется, глядя на эту проекцию, мы не должны забывать о ее объемном происхождении — линии заходят одна за другую, но не пересекаются. Интересно, что на один полный оборот спирали ДНК приходится десять пар оснований (мономеров ДНК), а на один полный оборот спирали в открытой системе приходится десять пар аминокислот.
Рис.бб. Проекция объемной Рис. 67. Проекция объемной системы аминокислот системы алфавита
Выводы
Теперь, когда в нашем исследовании явственно про* сматривается изоморфность азбуки и системы аминокислот белка, обратим внимание на то, что такое единство генетической основы белковой организации жизни и лингвистики в науке вовсе не является неправдоподобным. Роман Якобсон еще в 1970 г. писал: «Впечатляющие открытия последних лет в области молекулярной генетики описаны самими исследователями с помощью терминологии, заимствованной из лингвистики и теории коммуникации. Название книги Дж, и М. Бидлов " Язык жизни" — не просто фигуральное выражение, и исключительно высокая степень подобия систем генетической и языковой информации полностью объясняет ведущую мысль этой книги «расшифровка ДНК-кода выявила, что мы обладаем языком, который гораздо старше иерогли-фики, языком, который также стар, как сама жизнь, языком, который является самым живым из всех живых языков» [62, с.392]. Для нашего исследования азбуки еще более важными кажутся слова из речи Жакоба, Нобелевского лауреата по биологии, которые он произнес 7 мая 1965 г. в Colledge de France «...достойно удивления то, что генетическая информация записывается не посредством идеограмм, как в китайском языке, но посредством алфавита типа французского — или даже скорее алфавита азбуки Морзе. Смысл сообщения образуется путем объединения знаков в слова и слов во фразы...» [там же]. СВ. Петухов —физик, занимающийся проблемами симметрии в биологии [41, с.39], резюмируя исследования выдающихся психологов и физиологов мира Г. Гельм-гольца, И.М. Сеченова, Э. Кассирера и А. Пуанкаре, свою гипотезу формулирует как вопрос: «Не имеет ли организм тенденции воспринимать окружающий мир в образах тех самых групп преобразований (совокупность групп преобразований определяет полную симметрию — Г.Д.), в соответствии с принципами, из которых он сам сформирован? Не родственны ли принципы структуризации, реализуемые в морфогенетических18 феноменах и психологических явлениях пространственного восприятия? » Аналогично можно сформулировать подобную гипотезу уже о речи: «Не имеет ли индивид тенденции осмысливать окружающий мир в образах тех структур, из которых он сформирован? Не родственны ли принципы структуризации и функционирования, реализованные в морфогенетических и психологических феноменах памяти, мышления и речи? » Удивительно, что все мое исследование построено на, казалось бы, абсолютно «ненаучных» принципах. К системе азбуки я подошел сначала через аналогию звука и атома, а затем через фантастическую идею единого, планетарного и вечного алфавита. К системе аминокислот (и к ее изоморфности азбуке) — через, на первый взгляд, абсурдную идею о высочайшем уровне древних знаний, отраженных в клинописи Угарита, и «пифагорейскую» веру в то, что числа... сущности природы. Причем не просто числа, не числа привычной арифметики, а числа нумерологии — «астрологические» коды, когда смыслом чисел 21 и 30 считают три... Обратим внимание, что, строя систему аминокислот (см. рис.36, 37, 42), на первом этапе я практически не пользовался своими знаниями строения молекул, я пользовался лишь, если угодно, «менделеевским» принципом—использовал порядковые сущностные номера (обратим внимание, что не массы). Немаловажно для нашего доказательства, что такое «менделеевское» (даже точнее сказать — «пифагорейское») начало совпало с «Бутлеровским19», когда я разложил карты с прорисованными графическими формулами аминокислот. Построив на «странном» фундаменте нумерологии концепцию системы химических молекул, строго научно ее доказав, я прихожу к логичному выводу, что этот фундамент и есть истинная, самая настоящая, подлинно фундаментальная (! ) наука. Размышляя об учении Пифагора, о научности нумерологии, я взял в руки книгу синолога А.И. Кобзева и... круг замкнулся. Исследователь китайской классической философии в разделе своей книги, посвященном древней нумерологии и связи нумерологии и симметрии {! ), пишет о тех же замечательных ученых, на творчество которых я вышел через систему химии и алфавит: «Лауреат Нобелевской премии по биологии Жаков подробно исследовал разительные параллели древнекитайской " Книги перемен" (" И-цзин" ), обнаруженные им в современных представлениях о триплетах в генетическом коде. Эти идеи Жакоба были подхвачены и развиты великим филологом Романом Якобсоном, чьи труды о перекрещении разных наук <...> недавно <...> стали известны и нашему читателю. В своих новаторских трудах о роли симметрии в словесном искусстве Якобсон использовал и мысли о созвучии древнекитайского символического понимания симметрии, высказанные двумя другими Нобелевскими лауреатами по физике — Ли и Яном, чьи имена связаны с открытием особого рода симметрических отношений в физике» [33, с.5]. Обратим внимание, что все эти авторы пишут о генетическом коде, как об объекте изучения со стороны не только биологии, но и лингвистики. Попробуем и мы, в свете обнаруженной системы, посмотреть — а не дает ли это новых возможностей в понимании странностей кодирования. Действительно, некоторые аминокислоты кодируются четырьмя и даже шестью вариантами триплетов, а некоторые — всего одним. По крайней мере, мы имеем возможность представить главную «шифровальную таблицу» генетического кода не в случайном, а в закономерном виде (рис.68), используя известные данные по соответствию {триплет РНК — аминокислота} [45, с. 156]. Сразу видно, что наша система все-таки остается закономерностью аминокислот, а не генетического кода, Те закономерности чередования, которые заметны, весьма сложны, а закон кодирования наверняка прост, но, тем не менее, это очень важное начало, ведь до сих пор эту таблицу соответствия рассматривали вовсе без какого бы то ни было систематического принципа. Опишем некоторые закономерности нашей системы триплетных кодонов РНК (рис.69).
Рис, 68. Представление кодировки Рис. 69. Некоторые взаимосвязи аминокислот в системе кодонов аминокислот в системе
Кодон УГГ, кодирующий триптофан, аминокислоту, стоящую в системе последней в первом полупериоде линии аланина, принадлежит тому же четырехпозиционному набору кодонов {УГА, УГГ, УГЦ и УГУ}, что и кодон УГА, кодирующий конец белка. Кодон УАЦ-УАУ, кодирующий пос леднюю аминокислоту этой же линии, принадлежит тому же набору кодонов, что и кодон УАА — УАГ, который также кодирует конец белка. Кстати, это наблюдение косвенно (из соображений симметрии), что лишний раз подтверждает правильность чередования аминокислот в системе. Для простоты обозначения пары наборов кодонов я буду их называть числом — суммой соответствующих порядковых номеров {например, кодоны для пары {His—Trp} можно назвать числом 55}, ведь все аминокислоты в системе пронумерованы. Пары кодонов 11, 22, 33, 99, 110 можно «превратить» друг в друга заменой только одного нукле-отида, например, для пары 11, это замена Г на Ц, Особая симметричная пара — это пара с числом 77. Здесь для «превращения требуется замена любого из двух последних нуклеотидов на любой из двух других последних». Ограничимся этим, поскольку остальные взаимосвязи носят более сложный характер, и их описание вряд ли будет расшифровывать не биолог, а биолог и так разберется сам. Ну, а о результатах более глубокой проработки перспектив, открывающихся с обнаружением системы аминокислот, я расскажу в своей следующей работе. |
Последнее изменение этой страницы: 2019-03-30; Просмотров: 265; Нарушение авторского права страницы