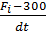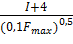|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ГИПЕРТРОФИЯ СКЕЛЕТНЫХ МЫШЦ ЧЕЛОВЕКА И МЕТОДЫ ЕЕ ОЦЕНКИСтр 1 из 31Следующая ⇒
СОДЕРЖАНИЕ 1 ГИПЕРТРОФИЯ СКЕЛЕТНЫХ МЫШЦ ЧЕЛОВЕКА И МЕТОДЫ ЕЕ ОЦЕНКИ 2 1.1 Классификации скелетных мышц. 2 1.2 Особенности функционирования веретенообразных и перистых мышц. 4 1.3 Морфологические показатели, характеризующие степень гипертрофии скелетных мышц, и методы их оценки. 5 1.3.1. Площадь поперечного сечения и объем скелетных мышц. 5 1.3.2. Методы оценки морфологических показателей скелетных мышц. 6 1.3.3. Обхватные размеры тела человека и способы их измерения. 7 1.4. Факторы, влияющие на гипертрофию скелетных мышц. 9 1.4.1. Локализация гипертрофической реакции мышцы.. 9 1.4.2. Расположение мышц. 10 1.4.3. Влияние пола на гипертрофию скелетных мышц. 11 1.4.4. Влияние возраста но гипертрофию скелетных мышц. 12 1.5. Влияние силовой тренировки на морфологические характеристики скелетных мышц. 12 КОНТРОЛЬНЫЕ ВОПРОСЫ... 12 2 ФАКТОРЫ, ВЛИЯЮЩИЕ НА ГИПЕРТРОФИЮ СКЕЛЕТНОЙ МЫШЦЫ... 14 2.1. Скелетная мышца как орган. 14 2.2. Соединение мышечных и сухожильных волокон. 19 2.3. Соединение мышечного волокна и двигательного нерва. 20 2.4. Управление активностью мышцы со стороны цнс. 20 2.5. Биохимия процессов сокращения на уровне мышцы.. 21 2.6. Параметры, определяющие объем скелетных мышц. 21 2.7. Методы оценки параметров, определяющих объем скелетных мышц. 23 2.8. Влияние силовой тренировки на параметры, определяющие объем скелетных мышц. 25 КОНТРОЛЬНЫЕ ВОПРОСЫ... 27 3 ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ГИПЕРТРОФИЮ СКЕЛЕТНОЙ МЫШЦЫ С УЧЕТОМ ТИПОВ МЫШЕЧНЫХ ВОЛОКОН.. 29 3.1. Типы мышечных волокон. 29 3.2. ТИПЫ ДВИГАТЕЛЬНЫХ ЕДИНИЦ.. 30 3.3. Регуляция силы и скорости сокращения мышцы центральной нервной системой. 31 3.4. Параметры, определяющие объем мышцы с учетом типов мышечных волокон. 33 3.5. Факторы, влияющие на площадь поперечного сечения мышечных волокон различных типов. 33 3.6. Влияние силовой тренировки на площадь поперечного сечения мышечных волокон различных типов. 35 3.7. Факторы, определяющие композицию мышечных волокон в скелетных мышцах. 36 3.8. Методы оценки композиции мышечных волокон в скелетных мышцах. 41 3.8.1. Инвазивные методы оценки композиции мышечных волокон в скелетных мышцах 41 3.8.2. Неинвазивные методы оценки композиции мышечных волокон в скелетных мышцах. 42 КОНТРОЛЬНЫЕ ВОПРОСЫ... 48 4 ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ГИПЕРТРОФИЮ СКЕЛЕТНОЙ МЫШЦЫ НА УРОВНЕ МЫШЕЧНОГО ВОЛОКНА.. 49 4.1. Состав мышечного волокна. 49 4.2. Строение мышечного волокна. 54 4.3. Сокращение и расслабление мышечного волокна. 56 4.4. Биохимические процессы, происходящие в мышечном волокне при сокращении и расслаблении мышцы.. 58 4.5. Состав, строение и морфофункционадьная характеристика мышечных волокон различных типов. 61 4.6. Параметры, определяющие объем мышечного волокна. 64 4.7. Влияние тренировки на параметры, определяющие гипертрофию мышечного волокна. 64 4.8. Гистогенез мышечных волокон. 66 4.9. Регенерация мышечных волокон. 67 КОНТРОЛЬНЫЕ ВОПРОСЫ... 70 5 ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ГИПЕРТРОФИЮ СКЕЛЕТНЫХ МЫШЦ НА УРОВНЕ МИОФИБРИЛЛЫ... 71 5.1. Состав и структура миофибриллы.. 71 5.2. Состав и структура саркомера. 71 5.2.1. Состав и структура толстого филамента. 74 5.2.2. Состав и структура тонкого филамента. 74 5.2.3. Состав и структура Z-диска. 75 5.2.4. Состав и структура М-диска. 76 5.3. Модель сокращения мышцы на уровне саркомера. 76 5.4. Параметры, определяющие объем миофибриллы.. 78 5.5. Влияние силовой тренировки на параметры миофибрилл. 79 КОНТРОЛЬНЫЕ ВОПРОСЫ... 80 6 ГИПЕРТРОФИЯ СКЕЛЕТНЫХ МЫШЦ КАК ПРОЯВЛЕНИЕ ДОЛГОВРЕМЕННОЙ АДАПТАЦИИ ЧЕЛОВЕКА К ФИЗИЧЕСКИМ НАГРУЗКАМ.. 81 6.1. Понятие адаптации организма человека к физическим нагрузкам.. 81 6.2. Виды адаптации и тренировочного эффекта. 81 6.3. Условия адаптации. 83 6.4. Виды гипертрофии мышечных волокон. 87 6.5. Гипотезы миофибриллярной гипертрофии мышечных волокон. 88 6.6. Механическое повреждение мышечных волокон как стимул повышенного синтеза белка в мышцах. 91 КОНТРОЛЬНЫЕ ВОПРОСЫ... 95 7 МЕТАБОЛИЗМ БЕЛКОВ В ОРГАНИЗМЕ ЧЕЛОВЕКА.. 97 7.1. Строение и функции нуклеиновых кислот. 97 7.2. Строение молекулы белка. 99 7.3. Переваривание и всасывание белков. 100 7.4. Катаболизм белков в мышечных волокнах. 101 7.5. Синтез белков в мышечных волокнах. 103 7.6. Миофибриллогенез. 105 7.7. Формирование новых мышечных волокон и их гиперплазия. 109 7.8 Концепции, объясняющие повышенный синтез бедка в скелетных мышцах при выполнении силовых тренировок. 111 КОНТРОЛЬНЫЕ ВОПРОСЫ... 115 8 ВЛИЯНИЕ РАЗЛИЧНЫХ ПАРАМЕТРОВ ТРЕНИРОВКИ НА ГИПЕРТРОФИЮ СКЕЛЕТНЫХ МЫШЦ.. 116 8.1. Факторы, сопутствующие миофибриллярной гипертрофии скелетных мышц. 116 8.1.1. Увеличение силы скелетных мышц. 116 8.1.2. Мышечные боли, возникающие ори выполнении силовых упражнений. 118 8.2. Влияние силовой тренировки с отягощениями различной массы на гипертрофию скелетных мышц. 121 8.2.1. Характеристика силовой тренировки с отягощением различной массы.. 121 8.2.2. Гипотеза, объясняющая тренировочные эффекты воздействия на скелетные мышцы отягощений различной массы.. 122 8.3. Влияние силовой тренировки, выполняемой в различных режимах мышечного сокращения, на гипертрофию скелетных мышц. 123 8.3.1. Тренировочные эффекты воздействия на скелетные мышцы работы в различных режимах. 123 8.3.2. Гипотезы, объясняющие тренировочные эффекты воздействияно скелетные мышцы роботы в различных режимах. 125 8.4. Влияние тренировки методом «до отказа» на гипертрофию скелетных мышц. 128 8.4.1. Характеристика метода «до отказа». 128 8.4.2. Тренировочные эффекты воздействия на скелетные мышцы тренировки методом «до отказа». 130 8.4.3. Гипотезы, объясняющие тренировочные эффекты воздействия но скелетные мышцы роботы методом «до отказа». 131 8.5. Влияние прекращения тренировки и последующего ее возобновления на гипертрофию скелетных мышц. 135 8.6. Влияние порядка упражнений, используемых в тренировке, на силу и гипертрофию скелетных мышц. 136 8.7. Влияние различных программ тренировки на силу и гипертрофию скелетных мышц. 137 КОНТРОЛЬНЫЕ ВОПРОСЫ... 138 ГЛОССАРИЙ... 140
Классификация по функции: · совместного действия — синергисты. Пример: мышц-синергистов: длинная и короткая головки двуглавой мышцы плеча; · противоположного действия — антагонисты. Пример мышц-антагонистов — двуглавая и трехглавая мышцы плеча. Совместная тяга мышц-синергистов определяет направление движения звена. В то же время мышцы-антагонисты, растягиваясь, тормозят движение. Мышцы сильные: · прикрепляются к большим поверхностям; · сокращаются медленно; · имеют короткие мышечные волокна. Пример: четырехглавая мышца бедра. Мышцы ловкие: · прикрепляются на небольших поверхностях; · сокращаются быстро; · имеют длинные мышечные волокна. Пример: лучевой сгибатель запястья. Таблица l.l Площадь SМ поперечного сечения мышц нижних конечностей у людей, не занимающихся физической культурой и спортом
F. М. Ivey et al. (2000) установили, что объем четырехглавой мышцы бедра у молодых мужчин равен 2297+170 см3, у молодых женщин — 1435+82 см3, у мужчин преклонного возраста — 1753 +44 см3, у женщин преклонного возраста — 1125 +53 см3. Таблица 1.2 Обхваты верхних и нижних конечностей у спортсменов высших разрядов в различных видах спорта, правая сторона тела (Ф. А. Завилейский, 1968)
Обхват плеча (в спокойном состоянии) измеряется в горизонтальной плоскости в месте наибольшего развития двуглавой мышцы плеча при свободно опущенной руке. Обхват плеча (в напряженном состоянии) измеряется также, но при сокращенных мышцах передней поверхности плеча. Разница в показателях между обхватом плеча в спокойном состоянии и обхватом плеча в напряженном состоянии характеризует экскурсию мышц плеча. Обхват предплечья измеряется в горизонтальной плоскости в месте наибольшего развития мышц предплечья при свободно опущенной руке. Обхват бедра измеряется аналогичным способом. Лента накладывается под ягодичной складкой и замыкается на наружной поверхности бедра. Обхват голени измеряется так же. Лента накладывается горизонтально в месте наибольшего развития трехглавой мышцы голени. Обхват груди в спокойном состоянии измеряется миллиметровой лентой, которая накладывается так, что сзади она проходит под нижним углом лопаток, сбоку — между туловищем и руками, а спереди закрывает нижние сегменты околососковых кружков (у женщин — верхний край грудных желез). Обхват груди при вдохе измеряется также, но во время максимального вдоха. При этом испытуемый не должен поднимать плеч. Обхват груди при выдохе измеряется так же, но во время максимального выдоха. Разница в показателях между обхватом груди при максимальном вдохе и при максимальном выдохе дает значение так называемой экскурсии грудной клетки. Обхват шеи измеряется путем накладывания ленты вокруг шеи горизонтально под кадыком. Обхват талии измеряется горизонтально: у мужчин — на уровне пупка, а у женщин — на уровне минимальной ширины брюшной полости. Обхват таза измеряется путем накладывания ленты в области тазобедренных суставов (Дж. Вейдер, 1992). Расположение мышц Известно, что у ранее не тренированных людей при стандартной тренировке наблюдается большая гипертрофия мышц верхних конечностей по сравнению с нижними. Так, S. Welle et al. (1996) установили, что у молодых мужчин после трех месяцев силовой тренировки площадь поперечного сечения мышц-сгибателей предплечья увеличилась на 22+4 %, а мышц-разгибателей голени — на 4±1 %. В то же время установлено, что мышцы нижних конечностей характеризуются большими значениями площади поперечного сечения по сравнению с мышцами верхних конечностей. Это связано с противодействием мышц нижних конечностей силе гравитации. При этом в большей степени ей противодействуют мышцы-разгибатели нижней конечности (антигравитационные мышцы). В связи с этим данные мышцы имеют большие массу и объем по сравнению со своими антагонистами. Соответственно, большие значения имеют их площадь поперечного сечения и обхват. Необходимость постоянного противодействия силе гравитации привела к тому, что у мышц нижней конечности отношение массы разгибателей к массе их антагонистов составляет 2: 1, а у мышц верхней конечности это соотношение равно 1: 1 (П.Ф. Лесгафт, 1905). Асимметрия физического развития приводит к тому, что обхватные размеры одной части тела, например правой, могут быть больше, чем другой. Существуют определенные соотношения между массой и ростом тела и его обхватными размерами (табл. 1.3). Таблица 1.3 Идеальные пропорции тела человека (Дж. Вейдер, 1992)
КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Охарактеризуйте существующие классификации скелетных мышц человека. 2. Дайте характеристику особенностей функционирования веретенообразных и перистых мышц человека. 3. Перечислите морфологические показатели, по степени изменения которых судят о гипертрофии скелетных мышц человека. 4. Перечислите методы оценки морфологических показателей, характеризующих гипертрофию скелетных мышц. 5. Приведите значения площади поперечного сечения и объема скелетных мышц человека. 6. Дайте характеристику значениям обхватных размеров тела человека и способам их измерения. 7. Дайте характеристику факторам, влияющим на гипертрофию скелетных мышц человека. 8. Опишите влияние гипертрофической силовой тренировки на морфологические характеристики скелетных мышц человека.
Скелетная мышца как орган Скелетные мышцы, несмотря на свою внешнюю простоту, представляют собой удивительные по своему составу и функциям сложнейшие, многоуровневые системы. В организме человека скелетные мышцы одновременно выполняют ряд разнообразных функций. Важнейшими из них являются: двигательная, рецепторная и преобразователя энергии. Рассматривая мышцу как орган, можно выделить восемь макрокомпонентов, из которых она состоит: 1. мышечные волокна; 2. соединительно-тканные оболочки; 3. сухожилия; 4. нервы; 5. рецепторы; 6. кровеносные сосуды; 7. лимфатические сосуды; 8. тканевая жидкость. 1. Мышечные волокна, объединенные в пучки, формируют брюшко мышцы, которое плавно переходит в сухожилие. По мере приближения к сухожилию мышечные волокна значительно сужаются, что придает брюшку мышцы его типичную веретенообразную форму. Мышечные волокна представляют собой компонент мышцы, в котором происходит преобразование химической энергии в механическую (механическое сокращение). Если рассечь мышцу поперек мышечных волокон (рис. 2.1), то можно увидеть, что снаружи мышца окружена плотной соединительной тканью — эпимизием. Эпимизий состоит из пучков коллагеновых волокон. Разрезав эпимизий, можно увидеть пучки мышечных волокон, как бы «завернутых» в оболочку соединительной ткани. Эта соединительно-тканная оболочка называется перимизием. Перимизий также достаточно плотный и относительно толстый. Поперечное сечение пучков мышечных волокон представляет собой фигуру сложной формы. Разрезав перимизий, можно увидеть отдельные мышечные волокна, окруженные рыхлой соединительной тканью. Эта оболочка называется эндомизием. Следует отметить, что перимизий не только окутывает пучки мышечных волокон, но и соединяет их с эпимизием.
Рис. 2.1. Соединительно-тканные структуры мышцы (В. С. Гурфинкель, Ю. С. Левик, 1985): 1 — перимизий; 2 — эндомизий; 3 — эпимизий 2. Соединительно-тканные оболочки играют роль футляров, в которые заключены мышечные волокна, пучки MB и мышцы, а также роль среды (перемизий), в которой есть каналы для кровеносных и лимфатических сосудов, а также нервов. Кроме того, соединительно-тканные оболочки противостоят активному и пассивному растяжению мышцы и передают усилие от мышцы сухожилию (А. Вайн, 1990). 3. Сухожилия состоят из плотной волокнистой соединительной ткани, богатой коллагеновыми волокнами. Они формируются как продолжение внутримышечных соединительно-тканных элементов и вплетаются в надкостницу. Сухожилие мало растяжимо, обладает значительной прочностью и выдерживает огромные нагрузки. Основными частями сухожилий являются толстые, плотно уложенные коллагеновые пучки разной длины и толщины. Для них характерна продольная полосатость, и во многих местах они сливаются друг с другом. Предел прочности сухожилий 40—60 МПа. Основная функция сухожилия — передача усилия от мышцы костному звену. 4. Нерв, подходящий к мышце, содержит три вида волокон: двигательные, вегетативные и чувствительные. По двигательным волокнам к мышце поступают импульсы из центральной нервной системы (ЦНС), побуждающие ее к сокращению. После входа в мышцу двигательные волокна ветвятся, и к каждому мышечному волокну подходит одна веточка, которая его иннервирует. Вегетативные волокна проводят к мышце импульсы из соответствующих вегетативных центров, влияющих на адаптационно-трофические функции (обмен веществ, состояние стенок сосудов, рост и развитие мышцы). По чувствительным волокнам импульсы поступают от мышцы в мозг. Одни из них проводят импульсы после температурных и болевых раздражений, другие сигнализируют о состоянии мышцы: степени напряжения, длине и скорости сокращения. 5. Рецепторы — специальные образования, информирующие нервную систему о длине, скорости сокращения мышцы, ее напряжении, а также о боли. Рецепторы опорно-двигательного аппарата (ОДА) расположены между мышечными волокнами, в сухожилиях, а также в суставной капсуле. Помимо мышечных волокон, осуществляющих сокращение мышцы, в ней имеются сложные по своей организации рецепторные органы — мышечные веретена. Мышечные веретена своими концами прикрепляются к мышечным волокнам или к перимизию и мышечному волокну. Такое параллельное расположение мышечных веретен относительно мышечных волокон приводит к тому, что их длина изменяется в соответствии с длиной мышцы. Внутри мышечного веретена расположен рецепторный аппарат. Когда мышца уменьшает свою длину, снижается натяжение мышечного веретена, активность рецепторов понижается. Когда мышца расслабляется (увеличивает свою длину), ее веретено растягивается и активность рецепторов повышается. Установлено, что от рецепторов мышечных веретен посредством афферентных (чувствительных) нервов в ЦНС поступает информация не только о текущей длине, но и о скорости сокращения мышцы (Р. В. С. Matthews, 1963; A. Prochazka, 1997). В месте перехода мышечных волокон в сухожилие располагаются сухожильные рецепторы (сухожильные органы Гольджи), которые информируют ЦНС о степени напряжения мышцы. Когда мышечные волокна сокращаются, коллагеновые волокна сухожилия натягиваются (рис. 2.2) и сжимают нервные веточки, которые начинают импульси ровать. Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам и сухожилию они возбуждаются при укорочении возбужденной мышцы. Следует отметить, что сухожильные рецепторы возбуждаются в 1, 5-8 раз более эффективно при укорочении мышцы, нежели при ее растяжении. Сухожильные рецепторы выполняют защитную функцию, снижая вероятность травмы. При их возбуждении тормозные импульсы поступают к мышцам-агонистам, а возбуждающие — к мышцам-антагонистам. Считается, что уменьшение влияния рецепторов Гольджи приводит к более мощному сокращению мышц. Этот механизм объясняет, по крайней мере частично, прирост мышечной силы вследствие тренировок силовой направленности.
Рис. 2.2. Строение сухожильного органа Гольджи (А. Дж. Мак-Комас, 2001) 6. Кровеносные сосуды (артерии и вены), также как и лимфатические, входят в мышцу и выходят из нее вместе с нервами. Через кровеносные сосуды мышца получает питательные вещества, кислород, гормоны и отдает продукты обмена веществ (углекислый газ, воду, соли и т. д.). Артерии, проникая в мышцу через эпимизий, ветвятся в перимизии. Ветвление продолжается и в эндомизии, где располагаются капилляры кровеносных сосудов. Они идут вдоль мышечных волокон. При этом на одно мышечное волокно приходится от одного до пяти капилляров. Процесс диффузии кислорода и субстратов осуществляется через стенки мышечных волокон. 7. Лимфатические сосуды представляют собой часть лимфатической системы. Ее функциями являются дренаж тканей, фильтрация, поддержание количества и состава тканевой жидкости, удаление из нее чужеродных веществ, образовавшихся в организме, а также участие в иммунных реакциях (М. Г. Ткачук, И. А. Степаник, 2010). По строению своей стенки лимфатические сосуды напоминают вены, так как все лимфатические сосуды снабжены богато развитой системой клапанов. На месте каждого клапана сосуд немного расширяется, что придает лимфатическим сосудам весьма характерный вид. Стенки лимфатических сосудов еще больше приспособлены к «проталкиванию» находящейся в них жидкости, чем стенки вен. В связи с этим в лимфатических сосудах больше клапанов, а в стенках сильнее развита мускулатура. Лимфатические сосуды со своими клапанами являются как бы насосом. Лимфатические сосуды, укрупняясь, образуют лимфатические стволы, которые в итоге сливаются в два лимфатических протока, открывающихся в венозное русло. Параллельно артериальным сосудам в мышце идут венозные и лимфатические сосуды. Мышечные волокна окружены лимфатическими капиллярами, которые выполняют в основном дренажную функцию — отток тканевой жидкости, содержащей продукты обмена веществ и инородные вещества, вначале в лимфатические сосуды, между которыми расположены лимфатические узлы, после этого — в лимфатические протоки, а затем — в крупные вены шеи. Любые частицы, попавшие в лимфу, задерживаются в лимфатических узлах, где сталкиваются с лимфоцитами. Лимфоциты, относящиеся к белым клеткам крови (лейкоцитам), циркулируют в лимфе и крови и составляют преобладающий тип клеток лимфоидных органов. В их функцию входит формирование иммунного ответа на внедрившиеся в организм бактерии и вирусы. Благодаря тому, что диаметр лимфатических капилляров в несколько раз больше, чем кровеносных, стенки очень тонкие и сильно проницаемые, через лимфатическую систему удаляются продукты обмена веществ, которые не могут попасть в кровеносные сосуды: большие молекулы и частицы, в том числе бактерии, которые не могут проникнуть в кровеносные капилляры. В лимфатических капиллярах и сосудах скелетных мышц поток лимфы обеспечивается сокращениями окружающих их скелетных мышц. Объемная скорость потока лимфы при мышечной работе может возрастать по сравнению с покоем в 10—15 раз. 8. Тканевая жидкость — жидкость, омывающая мышечные волокна и другие компоненты мышцы. Она соприкасается со всеми тканевыми элементами и является, наряду с кровью и лимфой, внутренней средой организма. Из тканевой жидкости мышечные волокна поглощают необходимые питательные вещества и выводят в нее продукты обмена. Оттекая от органов в лимфатические сосуды, тканевая жидкость превращается в лимфу. Рис. 2.4. Биопсия мышцы (Дж. X. Уилмор, Д. Л. Костилл, 1997) Перед проведением микроскопического исследования мышечные волокна замораживаются, а затем производится их поперечный разрез. После этого посредством оптического микроскопа определяются площадь поперечного сечения мышечных волокон (Н. Hoppeler, 1986) и площадь SНС, занимаемая несократительными компонентами мышцы. Следует заметить, что этот метод имеет существенный недостаток: мышечная ткань при уколе сильно сокращается и нормальное расположение мышечных волокон нарушается. На основе значений площади поперечного сечения мышцы, средней площади поперечного сечения мышечного волокна и площади, занимаемой несократительными компонентами мышцы, определяется количество мышечных волокон:
n МВ = S М - S НС / SMB
где nМВ — количество мышечных волокон; SМ — площадь поперечного сечения мышцы; SНС — площадь поперечного сечения несократительной части мышцы; SMB — площадь поперечного сечения мышечного волокна. Следует отметить, что данный метод оценки количества мышечных волокон имеет большую погрешность, так как значение площади поперечного сечения мышечных волокон значительно варьирует. Оно зависит от места, где взята биопсийная проба, утла сечения, количества разрезов мышечного волокна (Н. Hoppeler, 1986). Способ оценки длины мышечных волокон. Так как мышечные волокна представляют собой достаточно большие объекты (от 2 до 12 см), измерение их длины не представляет трудностей и может осуществляться любыми инструментами для измерения длиннотных размеров, имеющими миллиметровую шкалу. КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Перечислите основные компоненты мышцы. 2. Какова основная функция мышечных волокон? 3. Как называются оболочки, окружающие мышечные волокна, пучки мы шечных волокон и всю мышцу в целом? 4. Каковы функции соединительно-тканных оболочек мышцы? 5. Какова функция сухожилий? 6. Дайте характеристику рецепторному аппарату мышцы. 7. Какую функцию выполняют мышечные веретена? 8. Какую функцию выполняют рецепторы Гольджи? 9. Какую функцию выполняют кровеносные сосуды? 10. Каковы функции лимфатических сосудов и тканевой жидкости? 11. Как называется соединение мышечного волокна и нерва? 12. Как называются вещества, выделяемые в синаптическую щель при пере даче нервного импульса от мотонейрона мышце? 13. Что такое двигательная единица? 14. Дайте характеристику реакции гидролиза АТФ. 15. Что такое ресинтез АТФ? 16. Перечислите параметры, определяющие объем скелетной мышцы. 17. Дайте определение гипертрофии и гиперплазии мышечных волокон. 18. Опишите методы оценки параметров, определяющих объем мышцы. 19. Дайте характеристику влияния силовой тренировки на параметры, определяющие объем скелетной мышцы. Типы мышечных волокон В настоящее время общепринято считать, что у человека скелетные мышцы состоят из волокон различных типов. Медленные мышечные волокна (красные) обозначаются как волокна типа I. Быстрые мышечные волокна (белые) обозначаются как волокна типа II. Среди быстрых волокон различают волокна типа IIA (устойчивые к утомлению) и типа IIB (быстроутомляемые) (табл. 3.1). Основная функция мышечных волокон типа I — выполнение длительной работы низкой интенсивности. Они активны также при поддержании позы. Поэтому антигравитационные мышцы в основном состоят из медленных волокон типа I. Мышечные волокна типов IIА и IIB способны сокращаться с большой силой и скоростью. В среднем у лиц, не занимающихся физической культурой и спортом, площадь поперечного сечения волокон типа I меньше, чем волокон типа II. В мышцах волокна объединены в пучки, которые содержат мышечные волокна различных типов. Волокна в пучке расположены в виде мозаики (рис. 3.1). Однако, как показали исследования (J. Lexcell, К. Henrikssen- Larrsen, М. Sjostrom, 1983), MB типа I расположены более глубоко, a MB типа II — на периферии мышцы. Таблица 3.1 Характеристики мышечных волокон различных типов
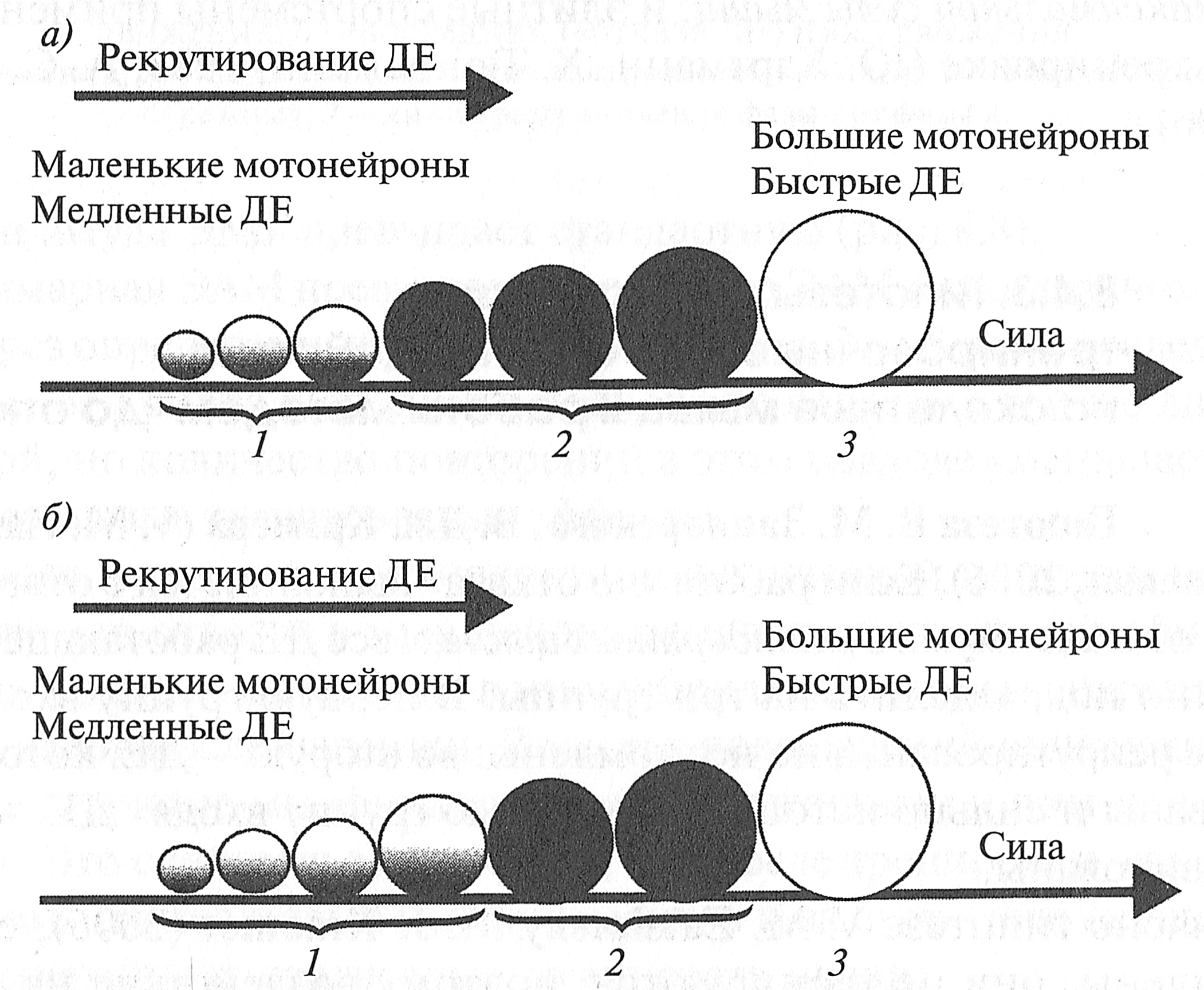
Рис. 3.1. Типы мышечных волокон в скелетных мышцах человека (Дж. X. Уилмор, Д. Л. Костилл, 1997). Темный цвет — тип I, серый цвет — тип IIA, белый цвет — тип IIВ ТИПЫ ДВИГАТЕЛЬНЫХ ЕДИНИЦ Напомним, что под словосочетанием двигательная единица (ДЕ) понимается функциональная система, состоящая из мотонейрона и иннервируемых им мышечных волокон. Существуют различные классификации ДЕ. Исходя из значимости для организма Р. Берк с соавт. (R. Е. Burke et al., 1973) предложил разделять ДЕ по сочетанию двух признаков — скорости сокращения и устойчивости к утомлению. По этой классификации ДЕ делятся на три типа: S (slow) — медленные, устойчивые к утомлению; FR (fast resistant) — быстрые, устойчивые к утомлению, FF (fast fatigable) — быстрые, быстроутомляемые. Этим ДЕ соответствуют различные типы мышечных волокон: Тип ДЕ S FR FF Тип мышечного волокна Тип I Тип IIА Тип IIB Строение и функции мотонейрона соответствуют морфологическим характеристикам мышечных волокон, которые он иннервирует. Так, мотонейрон ДЕ типа S имеет небольшое клеточное тело и иннервирует от 10 до 180 мышечных волокон, а мотонейрон ДЕ типа FF имеет большое клеточное тело и иннервирует от 300 до 800 мышечных волокон (Дж. X. Уилмор, Д. JL Костилл, 1997). Двигательные единицы типа S имеют низкий порог активации, поэтому при развитии силы мышцы они включаются в работу первыми. Двигательные единицы типа FF обладают высоким входным сопротивлением, поэтому при развитии усилия в мышце они активируются последними. Благодаря тому что мышечные волокна, принадлежащие различным ДЕ, рассредоточены по всей мышце, а не находятся в одном пучке, развитие силы мышцы характеризуется плавностью. Однако из-за того, что между соседними мышечными волокнами существуют соединительно-тканные связи, при сокращении одних мышечных волокон (например, входящих в состав ДЕ типа S) и расслабленном состоянии других (например, входящих в состав ДЕ типа FF) должны возникать силы трения, обусловливающие высокую вязкость мышцы. Г. В. Васюков (1967) показал, что при небольших напряжениях мышцы (30% от максимума) вязкость мышцы максимальна. При дальнейшем напряжении мышцы, когда одновременно возбуждено много мышечных волокон, вязкость мышцы скачкообразно уменьшается. Таблица 3.2 Площадь поперечного сечения различных типов мышечных волокон в зависимости от направленности тренировочного процесса
в то время как для верхних конечностей — 4090 мкм2 (двуглавая мышца плеча) и 4660 мкм2 (дельтовидная мышца). Такое превышение связано с тем, что мышцы нижних конечностей выполняют антигравитационную функцию. Пол. Для понимания механизмов, лежащих в основе адаптации мышц к силовой тренировке, существенным является тот факт, что у мужчин, не занимающихся физической культурой и спортом, площадь поперечного сечения мышечных волокон типа II больше, чем мышечных волокон типа I. Для мужчин этой группы отношение площади мышечного волокна типа II к площади мышечного волокна типа I составляет 1, 1—1, 4. У нетренирующихся женщин площадь поперечного сечения мышцы, приходящаяся на волокна различных типов, приблизительно равна, при этом отношение площади поперечного сечения мышечного волокна типа II к аналогичному показателю волокна типа I составляет 0, 9-1, 2. По данным В. Drinkwater (1988), площадь поперечного сечения волокон типа II на 40 % больше у тренированных мужчин по сравнению с нетренированными, в то время как тренированные женщины имеют только 15%-ное превосходство над нетренированными. Направленность тренировочного процесса. Установлено (табл. 3.2), что направленность тренировочного процесса значительно влияет на поперечное сечение мышечных волокон различных типов. Так, тренировка на выносливость (аэробная тренировка) приводит к преимущественному увеличению площади поперечного сечения мышечных волокон типа I. Силовая тренировка приводит к преимущественному увеличению площади поперечного сечения мышечных волокон типа II (табл. 3.2). Таблица 3.3 Композиция M B скелетных мышц человека
Установлено, что конституция человека является существенным фактором, определяющим композицию MB в скелетных мышцах человека, то есть состав MB в скелетных мышцах человека задан генетически. Генетическая предрасположенность во многом определяет отбор спортсменов для занятий тем или иным видом спорта. Установлено, что в мышцах спортсменов, занимающихся скоростно-силовыми видами спорта, преобладают мышечные волокна типа II (то есть мышечных волокон типа I меньше, чем мышечных волокон типа II) (рис. 3.2). В. В. Язвиковым и В.Г. Петрухиным (1991) показано, что у конькобежцев-спринтеров, имеющих очень высокую квалификацию (ЗМС), содержание в четырехглавой мышце бедра мышечных волокон типа I составляет всего 17, 0+7, 0 %, а у выдающихся спринтеров-легкоатлетов — не более 25 % (Дж. X. Уилмор, Д. JI. Костилл, 1997).
Рис. 3.2. Композиция MB латеральной широкой мышцы бедра у двух элитных спортсменов: а — пловца (кроль на 50 м); б — велосипедиста (шоссейные гонки), ( R. Billeter and Н. Hoppeler, 2003) Это означает, что в видах спорта, связанных с однократным выполнением работы максимальной мощности, преуспевают индивиды, в мышцах которых преобладают мышечные волокна типа II. В противоположность скоростно-силовым видам спорта у спортсменов, тренирующих выносливость, преобладают волокна типа I. Так, у спортсменов, тренирующихся в беге на длинные дистанции, содержание MB типа I составляет 73 % и выше (рис. 3.3). Таблица 3.4 Содержание MB различных типов в латеральной широкой мышце бедра у людей, не занимающихся спортом (J. A. Simoneau, С. Bourchard, 1989)
Таким образом, отбор существенно влияет на композицию мышечных волокон в мышцах спортсменов. У представителей скоростно-силовых видов спорта отмечено незначительное содержание мышечных волокон типа I в скелетных мышцах. У представителей видов спорта с направленностью тренировочного процесса на развитие выносливости мышечных волокон типа I значительно больше, чем типа II. Промежуточную группу представляют спортсмены, тренировка которых направлена на развитие силовой выносливости. Состав мышечных волокон типов I и II у этих спортсменов приблизительно одинаков. Таким образом, с одной стороны, композиция мышечных волокон может служить надежным генетическим маркером (Е. Б. Сологуб, В. А. Тай- мазов, 2002), а с другой — индивиды, у которых композиция мышечных волокон не соответствует требованиям работы в данном виде спорта, отсеиваются на этапах квалификационной лестницы (В. В. Язвиков, В. Г. Петрухин, 1991).
Рис. 3.3. Количество мышечных волокон типа I, адекватное для работы различной продолжительности (Е. Б. Сологуб, В. А. Таймазов, 2002) Пол. Исследованиями J. A. Simoneau, С. Bourchard (1989) доказано, что в латеральной широкой мышце бедра у женщин достоверно больше МВ типа I, в то время как у мужчин — МВ типа IIB (табл. 3.4). Возраст. Исследованиями В. Д. Сонькина и Р. В. Тамбовцевой (2011) доказано, что в ходе индивидуального развития человека MB проходят рад последовательных изменений. В возрасте одного года в мышцах детей отмечается большое количество MB типа I. Затем, по мере взросления детей, композиция MB в скелетных мышцах изменяется. Окончательное формирование состава MB в скелетных мышцах человека происходит к 17—20 годам. Затем, в течение долгого времени, процентное соотношение MB типа I и типа II в скелетных мышцах человека остается неизменным. Однако по мере старения (старше 70 лет) в скелетных мышцах пожилых людей процент MB типа II уменьшается.
Направленность тренировочного процесса. До настоящего времени вопрос о превращении одного типа мышечных волокон в другой под влиянием специфической тренировки окончательно не решен. Однако установлено, что процентное содержание в мышце волокон типа I генетически детерминировано и не меняется в процессе спортивной тренировки. Под воздействием тренировки на выносливость мышечные волокна типа IIB приобретают свойства мышечных волокон типа IIA (В. В. Язвиков, 1988; В. В. Язвиков, В. Г. Петрухин, 1991). Авторы объясняют такой тип превращения тем, что под воздействием тренировки на выносливость в мышечных волокнах типа IIB индуцируется синтез медленного миозина, что приводит к изменению активности АТФ-азы миозина, на основании которой определяется тип мышечных волокон. Скоростно-силовая тренировка не оказывает влияние на соотношение в мышце волокон типов IIАи IIB (В. В. Язвиков, 1988). Однако существуют противоречивые мнения относительно влияния силовой тренировки на преобразование МВ типа II. Согласно первой точке зрения, под воздействием силовой тренировки мышечные волокна типа IIA приобретают свойства мышечных волокон типа IIВ (В. Н. Платонов, 2005), то есть происходит процентное увеличение МВ типа IIB. Согласно второй точке зрения, которую поддерживает большое число ученых, после программы силовой тренировки происходит уменьшение процента мышечных волокон типа IIB и увеличение процента мышечных волокон типа IIA (R. S. Staron et al., 1991; G. R. Adams et al., 1993; J. L. Andersen, P. Aagaard, 2000; T. Hortobagyi, et al., 2000). В серии исследований R. S. Staron et al. (1990, 1991, 1994) изучалась адаптация скелетных мышц к силовым тренировкам у мужчин и женщин. После 20 недель тяжелых силовых тренировок у нетренирующихся женщин R. S. Staron et al. (1990) отметили значительное уменьшение количества волокон типа типа IIВ (16, 2 % до тренировок и 2, 7 % после тренировок) и параллельное увеличение количества волокон типа ILA (32, 5 % до тренировки и 39, 3 % после тренировки). Аналогичный эффект был установлен после шести недель тренировок (R. S. Staron et al., 1991). У женщин посредством биопсии (R. S. Staron et al., 1994) было выявлено значительное уменьшение доли МВ типа IIB уже после второй недели тренировок (то есть после четырех тренировок), а у мужчин — после четвертой недели тренировок. J. L. Andersen и P. Aagaard (2000) установили, что после трех месяцев силовой тренировки у нетренированных мужчин процент МВ типа IIВ в четырехглавой мышце бедра снизился с 9, 3+2, 1% до 2, 0±0, 8 % (Р< 0, 01). Соответственно увеличился процент МВ типа IIАс42, 4±3, 9 % до 49, 6±4, 0 % (Р< 0, 05). Последующее прекращение тренировок на три месяца вызвало значительное достоверное возрастание доли МВ типа IIВ (17, 2+3, 2 %, Р < 0, 01) и уменьшение доли мышечных волокон типа IIА. Рис. 3.4. Определение на кривых одиночного сокращения площадей, соответствующих мышечным волокнам типа IIB (АБВ), мышечным волокнам типа НА (АГД) и мышечным волокнам типа I (ЛЕЖ) (Н. В. Зимкин, М. С. Цветков, 1988) - - - предполагаемая скрытая часть восходящей кривой мышечных волокон типа IIA и мышечных волокон типа I Оценка композиции MB на основе тензиомиографии (ТМГ). В конце XX века в Словении (V. Valencic, N. Knez, 1997) для оценки композиции мышечных волокон была разработана методика тензиомиографии, идея которой полностью повторяла разработки исследователей из России. В 2004 году Войко Валенчич (V. Valencic, 2000) получил патент на изобретение метода и устройства селективной и неинвазивной оценки контрактильных свойств мышц. В этом устройстве посредством датчика измеряется радиальное увеличение мышечного брюшка. Важнейшим параметром, измеряемым по кривой ТМГ, является время сокращения мышцы. Следует отметить, что в отличие от российских исследователей ученым из Словении удалось разработать компактное и удобное устройство для оценки композиции мышц, которое могло бы быть применено в тренировочном процессе и реабилитации спортсменов различной квалификации. В настоящее время это устройство используется во многих известных футбольных клубах Европы, таких как FC Barcelona, FC Liverpool, FC Parma, FC Manchester city, ФК Динамо (Киев). Неинвазивные методы оценки композиции MB в нескольких скелетных мышцах. В настоящее время предложено несколько простых неинва- зивных методов оценки композиции мышц на основе измерения различных биомеханических характеристик. P. V. Komi, P. Tesch (1979) изучали максимальные произвольные разгибания ноги в колене на изокинетическом тренажере. Было установлено, что здоровые люди, имеющие большой процент быстрых мышечных волокон, демонстрировали более высокий момент силы при разгибании ноги, но быстрее утомлялись. После 100 сокращений мышцы у людей, имеющих высокий процент быстрых MB, достоверно снижалась интегрированная ЭМГ. Авторы предполагают, что в быстрых MB быстрее наступает отказ от работы. А. В. Шишкиной (2008) на основе идей, высказанных P. V. Komi, P. Tesch (1979), предложен метод косвенной оценки состава мышечных волокон. Однако, вместо работы на изокинетическом тренажере, анализировались изменения высоты выпрыгивания при прыжках с места. С этой целью исследуемые должны были выполнить 35 прыжков в удобном для них темпе с установкой: «прыгать вверх из положения полуприседа как можно выше в каждом прыжке». Посредством видеосъемки регистрировалась высота выпрыгивания. Затем рассчитывался показатель содержания медленных волокон в четырехглавой мышце бедра по следующей формуле: К= где Выбор показателя Суть неинвазивного метода косвенного определения композиции MB в мышцах-разгибателях ноги, предложенного В. Н. Селуяновым, Ю. В. Верхошанским и С. К. Сарсания (1985), заключается в следующем. Исследуемый располагается на силоизмерительной установке. Его туловище находится в вертикальном положении, угол между туловищем и бедром составляет 35°, между голенью и бедром — 110°. Стопа устанавливается на динамометрической площадке. После этого исследуемый выполняет два теста: 1. Тест МПС. Исследуемый с максимальной силой разгибает ногу в суставах (выполняются три попытки, выбирается лучшая). 2. Тест МБС. Исследуемый максимально быстро развивает изометрическую силу (выполняются пять—девять попыток, из них выбирается попытка, в которой достигается максимальный градиент / нарастания силы). Градиент нарастания силы вычисляется по формуле I= где Fi — максимальное значение силы в тесте, H, dt— время проявления силы начиная от 300 Н до максимума мс. После этого рассчитывается коэффициент К, который, по мнению авторов, может характеризовать отношение в мышцах-разгибателях ноги быстрых волокон к медленным. Коэффициент K вычисляется по формуле K= Дальнейшее исследование с участием спортсменов различной квалификации показало, что имеются достоверные различия между значениями К у спортсменов различных специализаций (табл. 3.5). Следует, однако, заметить, что методы, используемые в исследованиях В. Н. Селуянова, Ю. В. Верхошанского, С. К. Сарсания (1985), В. Н. Селуянова, М. П. Шестакова (2000), А. Ё. Шишкиной (2008), обладают существенным недостатком — они дают косвенную оценку композиции мышечных волокон в среднем для нескольких мышц (или мышечных групп) конечности, например для мышц-разгибателей ноги. Однако, как указывалось выше, композиция МВ в различных скелетных мышцах даже у одного и того же исследуемого различна. Поэтому эти методы дают очень приближенную оценку состава скелетных мышц человека. Следует упомянуть также об упрощенных методах оценки композиции МВ, которые, однако, обладают селективностью. Так, Т. Y. Pipes (1994) и N. Nilsson (2004) предлагают следующую последовательность действий при определении композиции МВ в скелетных мышцах. Вначале необходимо определить максимальное значение нагрузки в изолирующем упражнении (например, при выполнении сгибания руки в локтевом суставе со штангой, которое спортсмен может выполнить только один раз). Затем установить значение нагрузки, равное 80 % от максимума, и выполнить столько повторений, сколько спортсмен может, то есть работать «до отказа». После этого оценить результаты. По Т. V. Pipes (1996): мышца спортсмена, который выполнил меньше семи повторений, содержит более 50 % быстрых МВ. Если спортсмен выполнил от 7 до 12 повторений, в его мышце находится одинаковое количество быстрых и медленных волокон. Если спортсмен смог выполнить более 12 повторений, его мышца содержит более 50 % медленных МВ. Таблица 3.5 Скоростно-силовые характеристики мышц-разгибателей ноги у спортсменов разной специализации (В. Н. Селуянов, М. П. Шестаков, 2000)
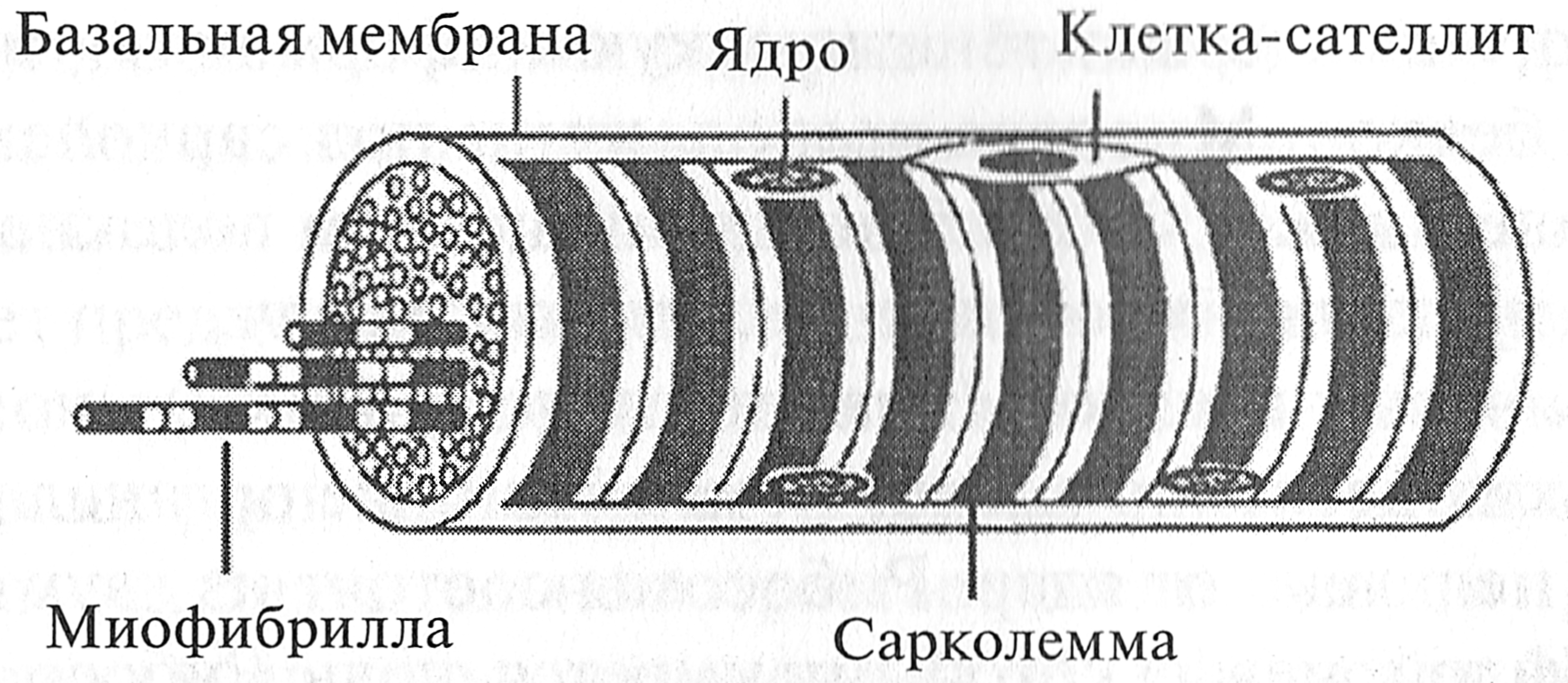
По N. Nilsson (2004): если спортсмен выполнил не более 4—7 повторений, у него в мышце преобладают быстро сокращающиеся волокна (тип IIB). Если спортсмен выполнил от 10 до 12 повторений, в его мышце преобладают промежуточные MB типа IIA. Если спортсмен сумел выполнить от 15 до 20 повторений, в его мышце преобладают медленно сокращающиеся волокна типа I. Следует заметить, что эти рекомендации основаны на эмпирическом подходе тренеров и не имеют экспериментального подтверждения. Таким образом, из всего вышесказанного можно сделать вывод, что определить в мышцах человека композицию мышечных волокон можно различными методами. Более точными считаются инвазивные методы, основанные на биопсии. Точность оценки композиции мышечных волокон посредством косвенных методов значительно ниже, однако они проще в использовании. КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Дайте характеристику различным типам мышечных волокон. 2. Какие типы двигательных единиц вы знаете? 3. Каково соответствие между типами ДЕ и типами МВ? 4. Посредством каких механизмов ЦНС регулирует уровень силы, развиваемой мышцей? 5. Укажите факторы, влияющие на площадь поперечного сечения различных типов мышечных волокон. 6. Как влияет гипертрофическая силовая тренировка на площадь поперечного сечения МВ различных типов? 7. Что такое композиция МВ? 8. Перечислите факторы, влияющие на композицию МВ. 9. Опишите влияние направленности тренировки на композицию МВ в скелетных мышцах человека. 10. Опишите прямые методы оценки композиции МВ. 11. Дайте характеристику неинвазивным (косвенным) методам оценки композиции МВ. 12. Опишите, как влияет силовая тренировка на характеристики мышечных волокон различных типов 1. Состав мышечного волокна Мышечное волокно имеет вид удлиненного цилиндра с закругленными концами. Очень часто его называют клеткой, но это не совсем правильно, так как клетка содержит только одно ядро и способна делиться. Мышечное волокно содержит до нескольких тысяч ядер и утратило способность к делению. В гистологии образования, в состав которого входит большое количество ядер, называют симпластами. В случае мышечного волокна это образование называется миосимпластом. Мышечное волокно, подобно термосу, имеет две оболочки. Внешняя оболочка мышечного волокна называется базальной мембраной. От нее отходят коллагеновые волокна, связывающие базальную мембрану с эндомизием. Внутренняя оболочка мышечного волокна называется сарколеммой (плазмалеммой). Между этими двумя оболочками в углублениях сарколеммы располагаются клетки-сателлиты (миосателлитоциты) (рис. 4.1).
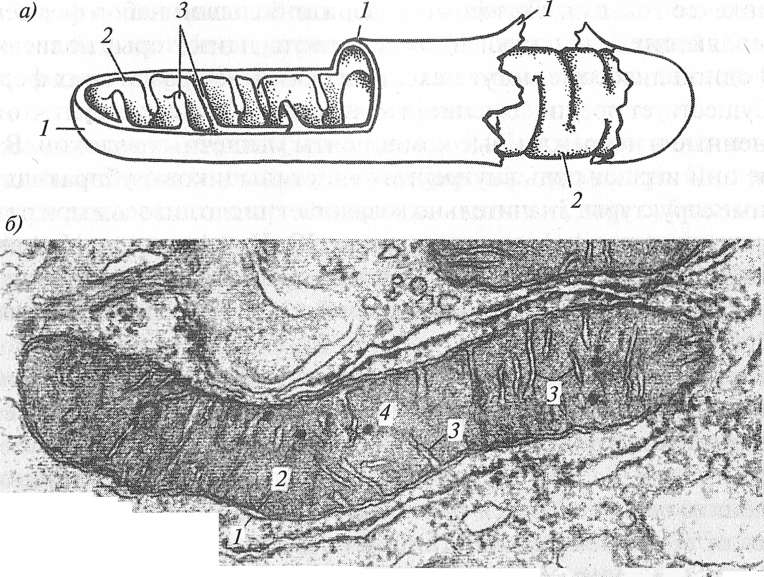
Рис. 4.1. Мышечное волокно (В. JI. Быков, 1998) Клетки-сателлиты играют важную роль в гипертрофии мышечного волокна. В обычных условиях деятельности человека они не проявляют высокой активности, однако при выполнении силовых упражнений активность клеток-сателлитов увеличивается, они начинают делиться, что приводит к повышенному синтезу белка. Клетки-сателлиты также участвуют в регенерации мышечных волокон. Объем внутри мышечного волокна заполнен гелеобразным коллоидным раствором — саркоплазмой. В ней протекают активные биохимические процессы расщепления и синтеза разнообразных органических веществ, обеспечивающих энергетическое снабжение сократительного аппарата. Саркоплазма обладает сравнительно высокой вязкостью, которая еще больше возрастает при возбуждении мышечного волокна. Вследствие этого она оказывает сопротивление укорочению миофибрилл, то есть создает внутреннее трение и в большей или меньшей мере замедляет сокращение или расслабление мышцы (Е. К. Жуков, 1969). Внутри саркоплазмы содержатся: · органеллы специального назначения; · органеллы общего назначения; · включения. Органеллами мышечного волокна специального назначения являются миофибриллы — тонкие нити, расположенные параллельно друг другу и идущие вдоль мышечного волокна. Миофибриллы занимают от 75 до 85 % объема мышечного волокна (Н. Hoppeler, 1986). В организме человека и животных миофибриллы являются главными преобразователями химической энергии в механическую. Вследствие этого преобразования происходит сокращение миофибрилл, мышечного волокна и всей мышцы. Миофибриллы состоят из саркомеров, разделенных Z-дисками. Органеллы общего назначения являются обязательными элементами любой клетки. К ним относятся: · миоядра (ядра); · рибосомы; · эндоплазматическая сеть; · комплекс Гольджи; · лизосомы; · митохондрии; · цитоскелет. Миоядра (ядра) — образования овальной формы длиной 10—20 мкм. В миоядрах находятся молекулы дезоксирибонуклеиновой кислоты (ДНК), которые содержат генетическую информацию, необходимую для синтеза белков. Миоядра расположены под сарколеммой. Количество миоядер, как и миофибрилл в мышечном волокне, достаточно большое: от нескольких сотен до нескольких тысяч. Рибосомы — очень мелкие (диаметром до 0, 02 мкм) многочисленные (до нескольких десятков тысяч) немембранные органеллы. Они располагаются недалеко от ядер. Рибосомы состоят из двух субчастиц — большой и малой, между которыми имеется щель. Рибосомы формируются в ядре, а затем через ядерные поры выходят в саркоплазму. Основная функция рибосом — синтез белков из аминокислот. Эндоплазматическая сеть (ретикулум) — мембранная органелла, представляющая собой разветвленную сеть трубочек и полостей. В мышечном волокне она представлена в виде шероховатой эндоплазматической сети и гладкой эндоплазматической сети (саркоплазматического ретикулума). Шероховатая эндоплазматическая сеть — мембранная органелла, окружающая миоядра. На ее поверхности располагаются рибосомы. На рибосомах синтезируются разнообразные белки, необходимые для нормального функционирования мышечного волокна: миозин, актин, тропонин, тропомиозин, десмин, виментин и многие другие. Эти белки представляют собой полипептидные цепочки (цепочки аминокислот). В полостях шероховатой эндоплазматической сети эти цепочки аминокислот обрезаются и сворачиваются. В результате возникает объемная трехмерная структура белка. Саркоплазматический ретикулум (гладкая эндоплазматическая сеть) — мембранная органелла, представляющая собой систему трубочек и мешочков (цистерн), окружающих миофибриллы. Основная функция саркоплазматического ретикулума — депонирование и выделение ионов кальция (Са2+), необходимых для процесса сокращения мышечного волокна. От поверхности мышечного волокна к расширенным участкам саркоплазматического ретикулума направляются выпячивания сарколеммы — поперечные трубочки, называемые Т-системой. Комплекс Голъджи — мембранная органелла, имеющая вид плоских цистерн, на периферии которых содержатся многочисленные мелкие пузырьки. Комплекс Гольджи расположен рядом с эндоплазматической сетью. В комплексе Гольджи происходит дальнейшее формирование структур белков и некоторых других веществ. После этого они сортируются и упаковываются в мембранные пузырьки, которые затем транспортируются в другие места мышечного волокна, где они необходимы. Некоторые вещества переносятся к сарколемме и выводятся за пределы мышечного волокна. Лизосомы — мембранные органеллы, представляющие собой микроскопические пузырьки, размером 0, 2—0, 4 мкм. Они формируются в комплексе Гольджи. Лизосомы содержат бол ьшой набор ферментов, расщепляющих белки, нуклеиновые кислоты и некоторые полисахариды. В одной лизосоме могут находиться до 30—50 различных ферментов. Существует предположение, что лизосомы отбирают и уничтожают измененные и поврежденные компоненты мышечных волокон. В этом случае они играют роль внутренних «чистильщиков», убирающих дефектные структуры. Значительно возрастает число лизосом при различных повреждениях мышечных волокон (Ю. И. Афанасьев, Н. А. Юри- I га, 1989). Лизосомы принимают участие и в регенеративных процессах, обеспечивающих гипертрофию мышечных волокон (Н. И. Волков с со- авт., 2000). Митохондрии — мембранные органеллы мышечного волокна размером до 2—3 мкм. В митохондриях протекает окисление углеводов, жиров и аминокислот до углекислого газа и воды с использованием кислорода воздуха. За счет энергии, выделяющейся в митохондриях при окислении, осуществляется синтез АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки или органеллами тканевого (клеточного) дыхания. Обычно митохондрии скапливаются вблизи тех участков саркоплазмы, где возникает потребность в АТФ. Митохондрии состоят из двух мембран (внешней и внутренней) и внутреннего содержимого — матрикса. Внутренняя мембрана образует выпячивания внутрь митохондрии — кристы (рис. 4.2). Именно на кристах происходит процесс окисления углеводов, жиров и аминокислот. Митохондрии в клетках могут увеличиваться в размерах и числе, то есть осуществляются их гипертрофия и гиперплазия. В последнем случае происходит деление перетяжкой или фрагментация исходных крупных митохондрий на более мелкие, которые, в свою очередь, могут расти и снова делиться. По расположению различают субсарколеммные и межфибриллярные митохондрии. Субсарколеммные митохондрии расположены непосредственно под сарколеммой мышечного волокна. Напомним, что в мышечном волокне под сарколеммой расположены многочисленные ядра и рибосомы, в которых происходит синтез белка. В связи с этим субсарколеммные митохондрии снабжают рибосомы необходимой энергией для синтеза белка. Они составляют 10-15 % от общего количества митохондрий (Н. Hoppeler, 1986). Межфибриллярные митохондрии расположены между миофибриллами. АТФ, выделяемая межфибиллярными митохондриями, необходима для процесса мышечного сокращения. Считается, что окислительная емкость клетки тесно связана с количеством в ней митохондрий. Объемная плотность митохондрий всех видов в мышечном волокне составляет от 3, 5 до 5, 7 % (Н. Hoppeler, 1986).
Рис. 4.2. Ультрамикроскопическое строение митохондрий (Ю. И. Афанасьев, Н. А. Юрина, 1989): а — схема; б — электронная микрофотография: 1 — наружная митохондриальная мембрана; 2 — внутренняя митохондриальная мембрана; 3 — кристы; 4 — митохондриальный матрикс Цитоскелет мышечного волокна представляет собой систему немембранных органелл, необходимых для создания определенной внутриклеточной структуры. Благодаря цитоскелету миофибриллы имеют постоянную локализацию и фиксацию, а мышечное волокно характеризуется поперечной исчерченностью. В саркоплазме мышечного волокна включения представлены в виде молекул АТФ, АДФ и АМФ, гранул гликогена, креатинфосфата (КрФ), липидов, белков и (или) их компонентов (миоглобина, ферментов и аминокислот), а также экстрактивных веществ. Строение мышечного волокна Исследования, проведенные в последние годы, свидетельствуют о том, что мышечное волокно имеет сложное строение. Расположение органелл общего и специального назначения строго упорядо- ченно. Выше указывалось, что между базальной мембранной и сарколеммой располагаются клетки-сателлиты. Миоядра разбросаны по всему волокну и в неповрежденном волокне находятся на его периферии под сарколеммой. Расстояние между миоядрами составляет приблизительно 5 мкм. Ядра окружает шероховатая эндоплазматическая сеть, на поверхности которой расположены рибосомы. Рядом с эндоплазматической сетью находится комплекс Гольджи, в котором происходит дальнейшее формирование структур белков и некоторых других веществ.
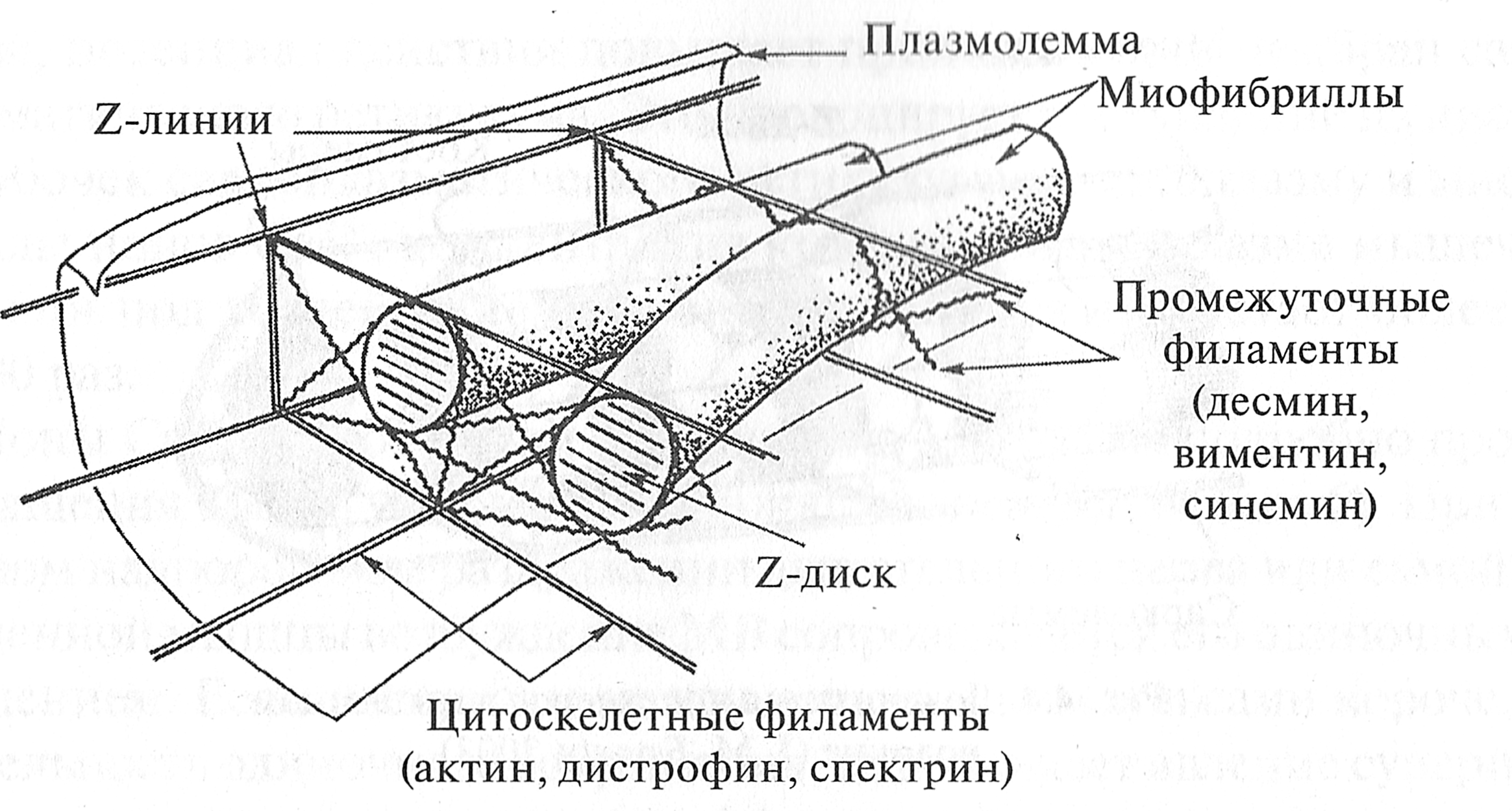
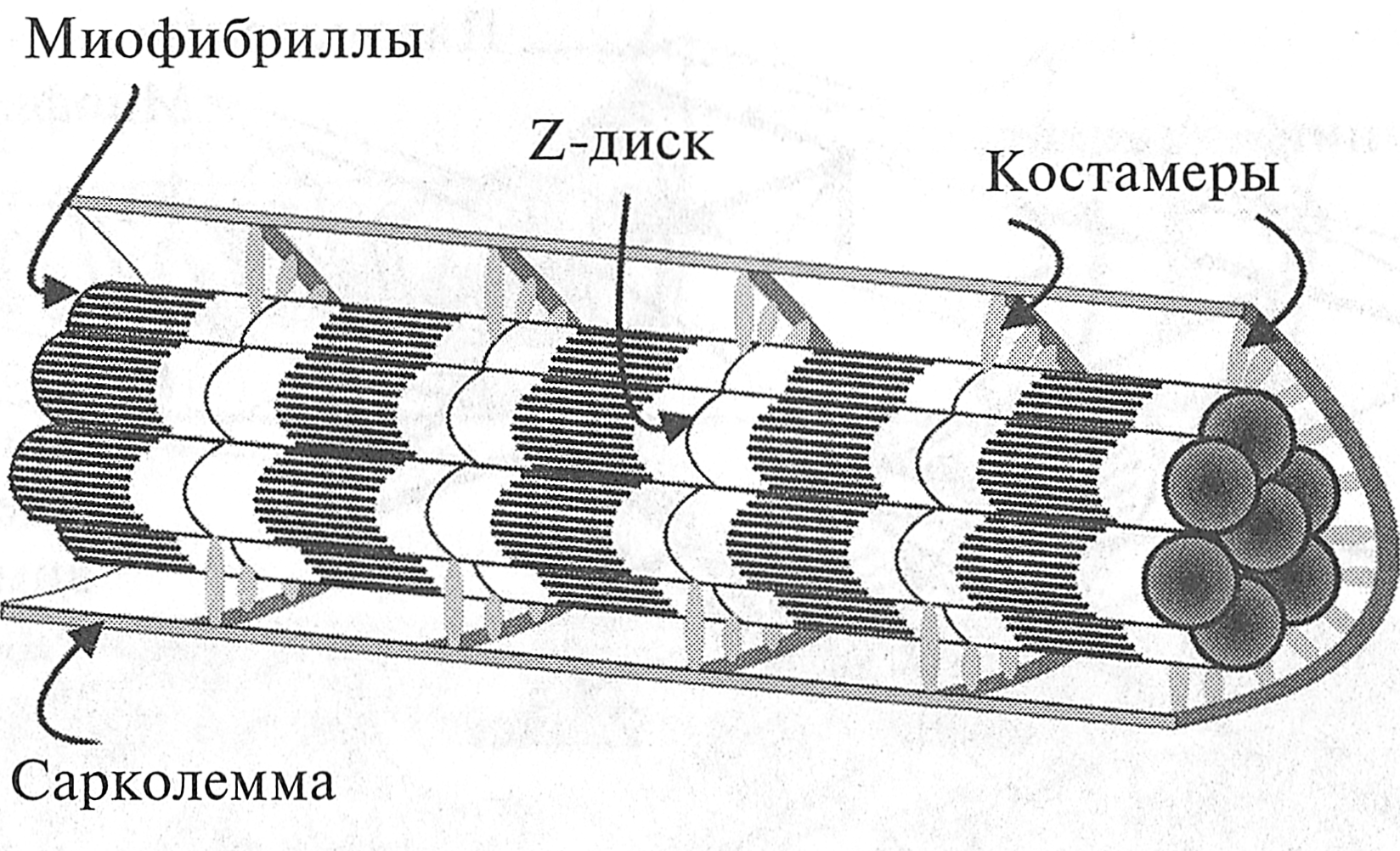
Рис. 4.3. Привязка Z-дисков к элементам цитоскелета (Е. Lazarides, Y. G. Kapetenaki, 1986) Расположение миофибрилл в волокне строго упорядочение. Существует специальная структура — цитоскелет мышечного волокна, обеспечивающая стабилизацию миофибрилл в мышечном волокне. Цитоскелет образован поперечными и продольными нитями, имеющими толщину около 10 нм (рис. 4.3). Основу поперечных нитей составляют структурные белки десмин, виментин и синемин. Поперечные нити соединяют соседние миофибриллы в области Z-дисков. Это обстоятельство определяет тот факт, что Z-диски всех миофибрилл мышечного волокна лежат друг против друга. Поэтому мышечное волокно обладает выраженной поперечной исчерченностью. Продольные нити, связывающие соседние Z-диски миофибриллы, состоят из структурных белков актина, дистрофина и спектрина. Благодаря своим упругим свойствам продольные нити предотвращают чрезмерное растяжение саркомера. Периферические миофибриллы имеют также связи с сарколеммой. Эти связи обеспечиваются за счет мембранного скелета мышечного волокна. Этот скелет расположен с внутренней стороны сарколеммы. Он представляет собой своеобразные ребра — костамеры, которые наподобие обручей обхватывают изнутри мышечное волокно (рис. 4.4). Костамеры состоят из различных структурных белков, одним из основных является белок винкулин. Костамеры связывают периферические миофибриллы с сарколеммой. Они прикрепляются к миофибриллам в области Z-дисков. Поэтому повреждение Z-дисков периферических миофибрилл может приводить к нарушениям в сарколемме мышечного волокна.
Рис. 4.4. Локализация костамеров в мышечном волокне (J. М. Ervasty, 2003)
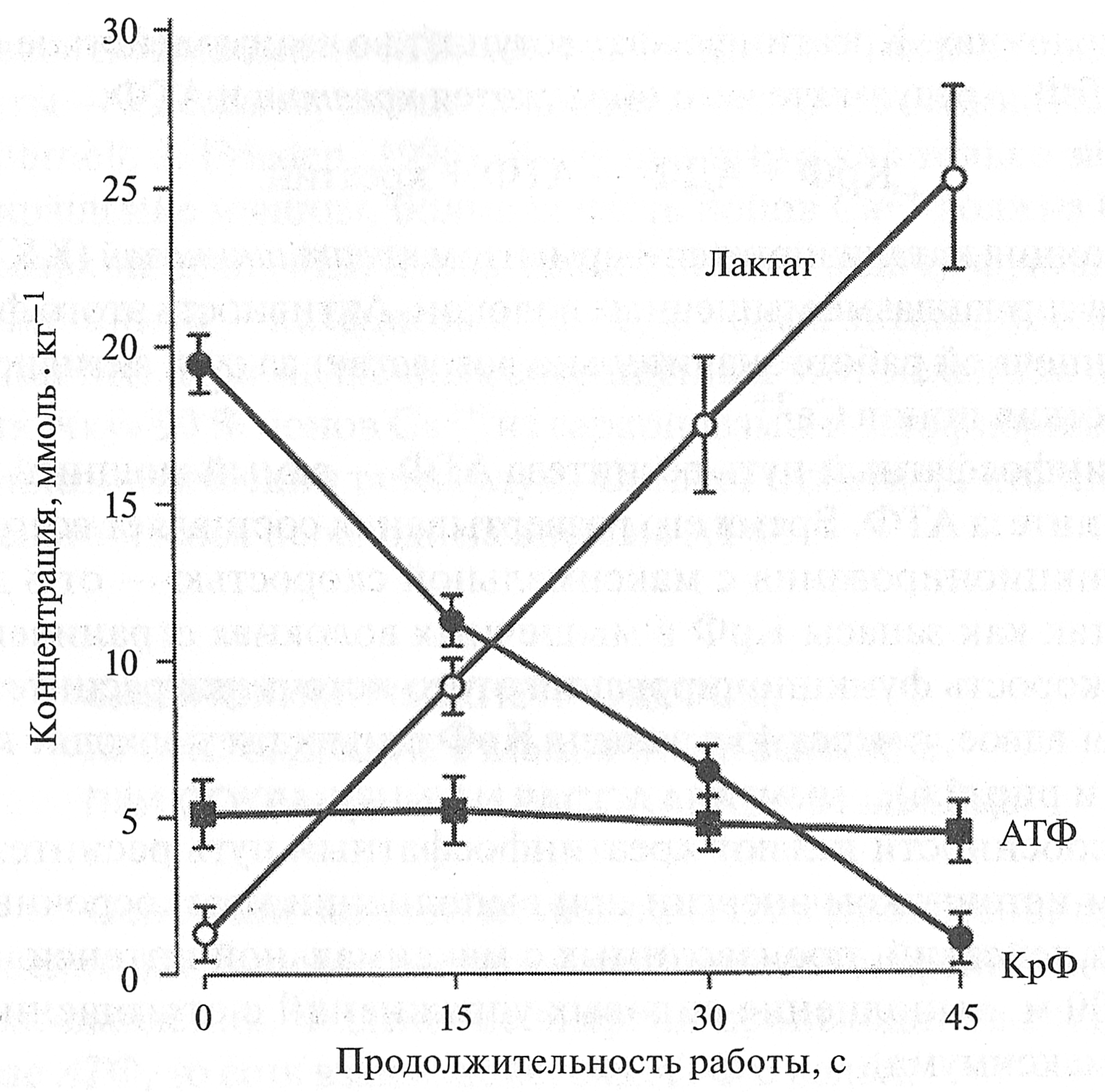
В отличие от других органелл мышечного волокна, миофибриллы не имеют оболочки, однако их наподобие муфты окружает саркоплазматический ретикулум. В пространстве между миофибриллами располагаются митохондрии. Расположение митохондрий вблизи миофибрилл диктуется необходимостью обеспечения процессов мышечного сокращения энергией. Выше указывалось, что в митохондриях происходят процессы тканевого дыхания, результатом которых является производство АТФ, необходимого для сокращения миофибрилл. Большая часть саркоплазмы находится в тончайших щелевидных пространствах между миофибриллами, вследствие чего общая поверхность соприкосновения саркоплазмы с сократительными элементами клетки очень велика. Это обстоятельство благоприятно для быстрого обмена веществ между саркоплазмой и миофибриллами (Е. К. Жуков, 1969). Таблица 4.1 Концентрация энергетических субстратов и рН в скелетных мышцах при выполнении физической работы (Н. И. Волков с соавт., 2000)
Эти особенности делают креатинфосфатный путь ресинтеза АТФ основным источником энергии при выполнении краткосрочных двигательных действий, производимых с максимальной интенсивностью (бег на 100 м, выполнение силовых упражнений с отягощением 70— 100 % от максимума).
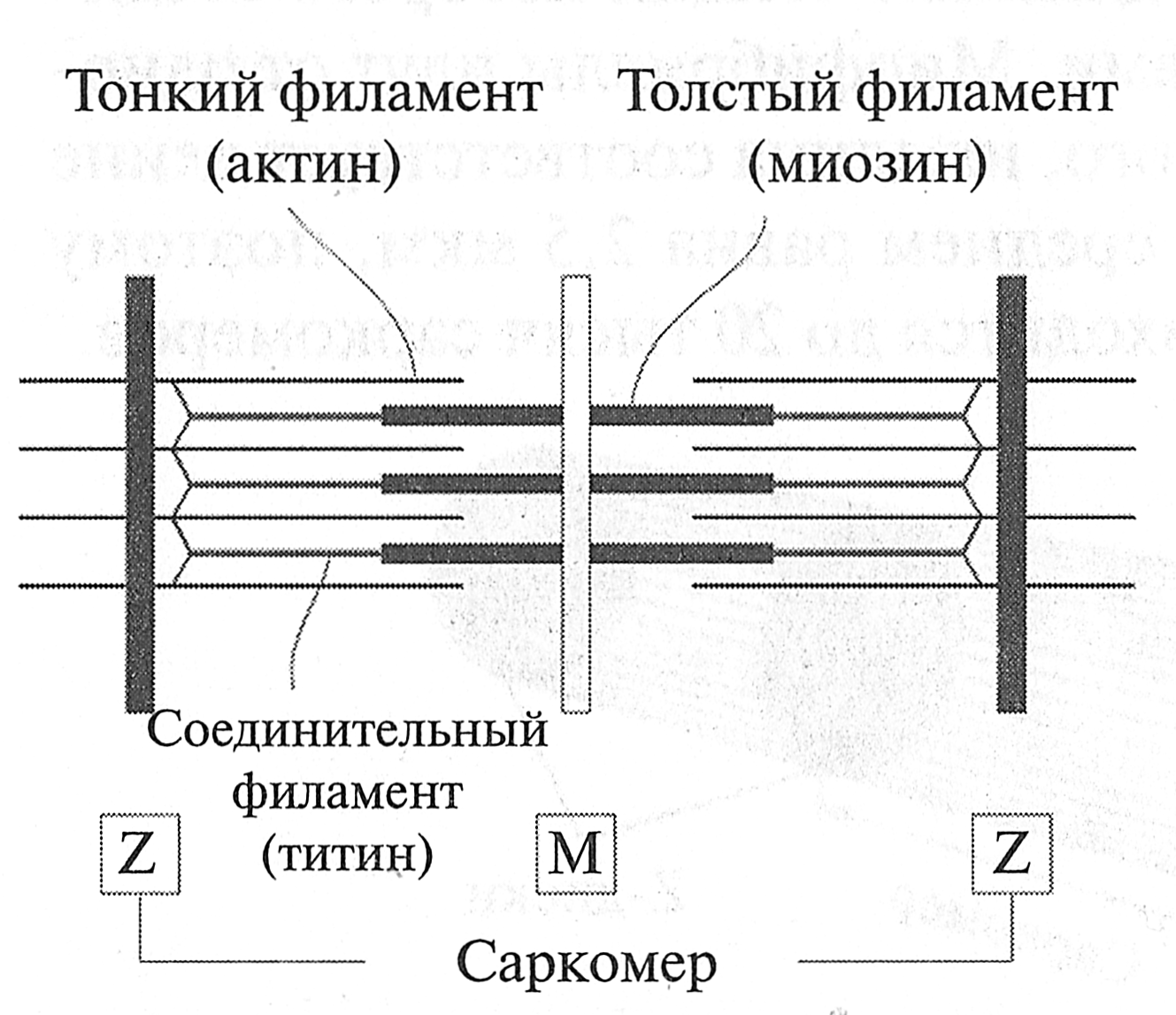
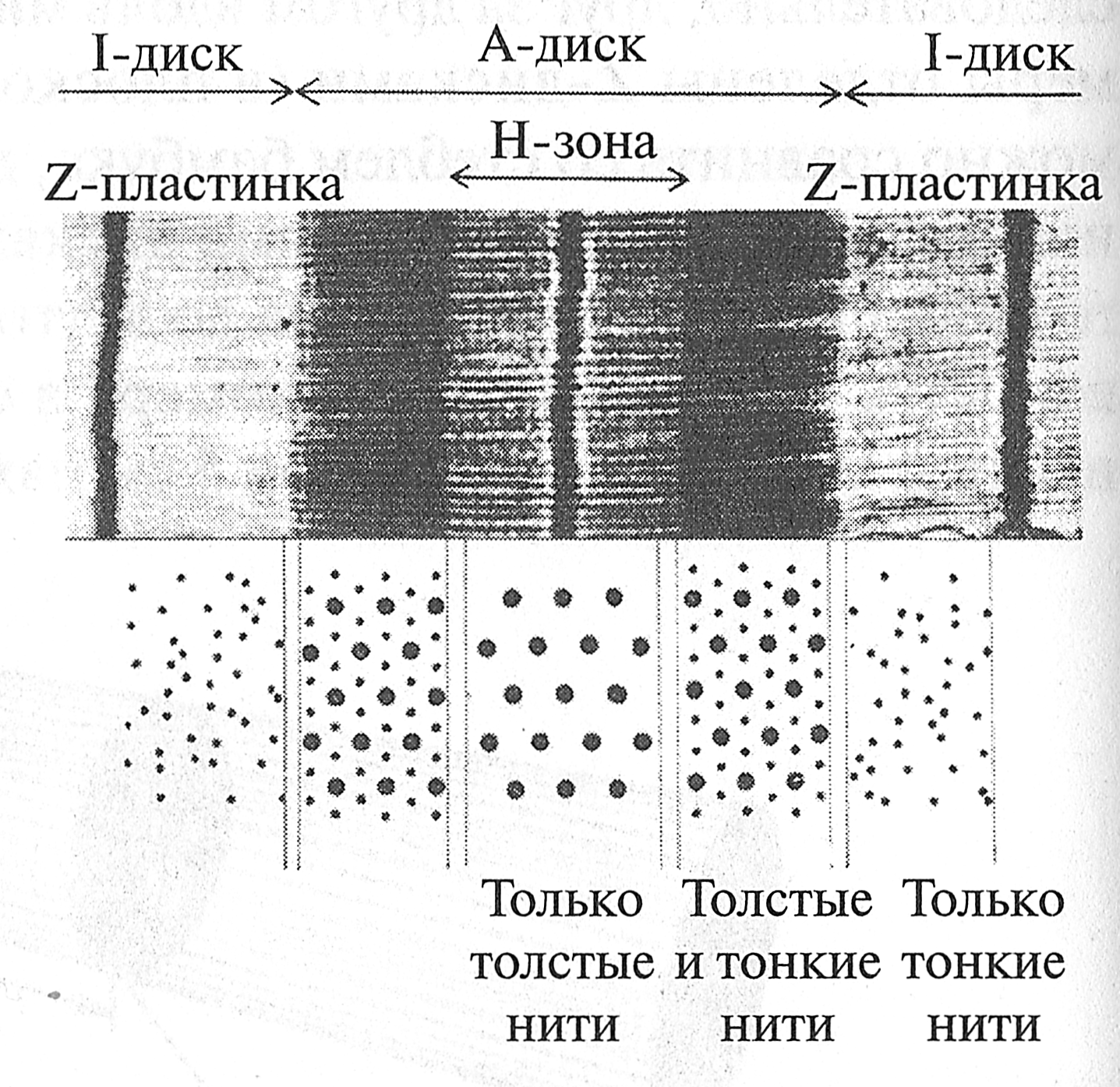
Гистогенез мышечных волокон Проблема гистогенеза мышечных волокон интересна тем, что при повреждении мышечной ткани и последующей ее регенерации происходят те же процессы, что и при гистогенезе. Установлено, что гипертрофия мышечных волокон также связана с их микротравмами и последующей их регенерацией. Источником развития элементов скелетной поперечно-полосатой мышечной ткани являются клетки миотомов. Одни из них дифференцируются на месте. Другие же мигрируют из миотомов в мезенхиму. Они уже детерминированы в направлении развития элементов мышечной ткани, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки будущих мышц. При этом возникают две линии дифференцировки. Из одних клеток (митотических миобластов) возникают клетки- сателлиты, которые, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни. Эта их способность обеспечивает регенерацию мышечной ткани. Из других клеток (постмитотических миобластов) образуются цепочки, которые сливаются друг с другом и клетками-сателлитами и образуют мышечную трубочку (миотубу). Ядра в мышечных трубочках расположены в центре. В мышечных трубочках происходит сборка миофибрилл, которые вначале располагаются под сарколеммой, а затем заполняют большую часть мышечной трубочки, оттесняя ядра к ее периферии. Миоядра к этому времени теряют способность к делению, и за ними остается функция хранения генетической информации, необходимой для синтеза белка. По мере дифференцировки изменяется тип вырабатываемого миозина, увеличивается содержание митохондрий, формируются элементы саркоплазматического ретикулума. Постепенно возникают различия в структурных, цитохимических и функциональных характеристиках, которые лежат в основе разделения мышечных волокон на типы. Окончательная дифференцировка мышечной трубочки происходит после возникновения ее иннервации. Миотубы вступают во взаимодействие с растущими аксонами мотонейронов, что способствует последующему развитию и дифференцировке волокон. Таким образом, мышечная трубочка превращается в мышечное волокно. КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Опишите состав мышечного волокна. 2. Опишите строение мышечного волокна. 3. Дайте характеристику органеллам специального назначения. 4. Дайте характеристику органеллам общего назначения. 5. Что представляют собой включения? 6. Каково строение (цитоскелет) мышечного волокна? 7. Опишите процессы, приводящие к сокращению и расслаблению мышечного волокна. 8. Дайте характеристику креатинфосфатному пути ресинтеза АТФ. 9. Дайте характеристику гликолитическому пути ресинтеза АТФ. 10. Дайте характеристику аэробному пути ресинтеза АТФ. 11. Опишите мышечные волокна различных типов на основе их состава и строения. 12. Опишите параметры, определяющие объем мышечного волокна. 13. Дайте характеристику саркоплазматической гипертрофии скелетной мышцы. 14. Охарактеризуйте процесс миофибриллярной гипертрофии скелетной мышцы. 15. Дайте характеристику гистогенезу мышечных волокон. 16. Опишите процесс регенерации мышечных волокон. Рис. 5.2. Строение сота. Толстый филамент, подобно грифелю карандаша, расположен в середине, на ребрах карандаша находятся тонкие филаменты. Структура Z-диска и прикрепление толстого филамента к Z-диску при помощи белка титина не показаны Если осуществить продольный разрез саркомера (рис. 5.3 и 5.4), то можно увидеть, что тонкие филаменты располагаются параллельно толстым и одним концом прикрепляются к Z-линии (Z-диску). Толстые филаменты размещены в центре саркомера. Одним концом они соединяются друг с другом и образуют М-зону (М-диск); другим концом толстые филаменты при помощи белка тишина прикрепляются к Z-линии (Z-диску).
Рис. 5.3. Строение саркомера (G.H. Pollak, 1990) Поперечный разрез саркомера в области I-дисков, А-дисков и Н-зоны (рис. 5.4) покажет совершенно другую картину. Если провести разрез в области светлых I-дисков, будут видны только тонкие филаменты. В области максимального затемнения (А-диски) поперечный разрез саркомера покажет сложную структуру, где один толстый фила- мент окружен шестью тонкими филаментами, а каждый тонкий — тремя толстыми. В области Н-зоны при поперечном разрезе видны только толстые филаменты.
Рис. 5.4. Вверху: микрофотография продольного разреза саркомера; внизу. поперечный разрез саркомера в его различных участках (Н. Е. Huxley, 1972) В начале изучения структуры саркомера в составе тонкого филамента был обнаружен белок актин, а в составе толстого — миозин. Поэтому иногда тонкий филамент называют актиновым филаментом, а толстый — миозиновым. Однако последующие исследования показали, что структура тонкого и толстого филаментов более сложная, чем предполагалось ранее. Рис. 5.6. Тонкий филамент, состоящий из молекул актина, тропомиозина и тропонина (Дж. X. Уилмор, Д. Л. Костилл, 1997) Состав и структура Z-диск а Выше было показано, что саркомеры в миофибрилле соединены друг с другом посредством Z-дисков (по первой букве немецкого слова zwischenscheibe — промежуточный диск). В состав Z-диска входят следующие белки: алъфа-актинин (осуществляет прикрепление тонких филаментов к Z-диску), десмин, виментин, синемин, дистрофии, спектрин. При этом белок алъфа-актинин-2 располагается преимущественно в MB типа I, а белок алъфа-актинин-3 — в MB типа IIB. Дефицит белка альфа-актинина-3 в быстрых волокнах может стать причиной низкого уровня развития скоростно-силовых качеств человека (И. И. Ахметов, 2009). Согласно модели, предложенной D. Е. Kelly (1967), Z-диск при продольном разрезе саркомера похож на зигзагообразную линию, к которой прикрепляются тонкие филаменты (рис. 5.7, а). При поперечном разрезе Z-диск похож на тканную корзину или квадратную решетку (рис. 5.7, б). Таблица 5.1 Толщина Z-дисков в мышечных волокнах различных типов у нетренированных мужчин
Для понимания механизма гипертрофии существенно, что у мышечных волокон различных типов толщина Z-дисков неодинакова. Самые толстые Z-диски у медленных волокон типа I. Самые тонкие — у мышечных волокон типа IIB (табл. 5.1). Из этого следует, что мышечные волокна типа IIВ могут быть легче повреждены в области Z-дисков по сравнению с мышечными волокнами типа I. Состав и структура М-диска Уже в первых исследованиях, в которых использовали оптический микроскоп, в центре А-диска наблюдали темную линию, которую назвали М-линией или М-диском (от немецкого слова mittelscheibe — центральный диск). Затем, применив электронный микроскоп, установили, что в действительности имеются несколько М-линий, поэтому более уместно говорить не о М-линии, а о М-зоне. Именно к этой зоне прикрепляются толстые филаменты саркомера. В состав М-диска входят белки: М-белок, миомезин, креатинкиназа. Толщина М-диска различна в мышечных волокнах различных типов. В волокнах типа I М-диск содержит пять сильных М-линий, в волокнах типа IIА — три сильные и две слабые линии, в волокнах типа IIB — три сильные и, возможно, две очень слабые линии (М. Sjostrom et al., 1982). Следует отметить, что в начале изучения быстрых и медленных типов мышечных волокон их классификация осуществлялась на основе измерения толщины Z- и М-дисков. 5.3. Модель сокращения мышцы на уровне саркомера Как сокращаются саркомер, миофибрилла, мышечное волокно и мышца в целом? Схему сокращения саркомера объясняет так называемая «теория скользящих нитей» или «гребковая гипотеза». Установлено, что во время сокращения (укорочения) саркомера длина тонкого и толстого филаментов не меняется. При этом неизменной особенностью сокращения является центральное положение толстого филамента в саркомере, посередине между Z-дисками. При поступлении по аксону мотонейрона нервного импульса нервные окончания выделяют нейромедиатор — ацетилхолин, который «привязывается» к рецепторам сарколеммы. При достаточном его количестве электрический заряд передается по всей длине волокна. Этот процесс называется развитием потенциала действия. Кроме деполяризации мембраны мышечного волокна, электрический импульс проходит через сеть трубочек волокна (Т-трубочки и саркоплазматический ретикулум) во внутреннюю часть MB. Поступление электрического импульса приводит к выделению значительного количества ионов Са2+ в саркоплазму. Следует заметить, что концентрация ионов Са2+ в сар- коплазматическом ретикулуме выше, чем в саркоплазме. После этого ионы Са2+ связываются с тропонином, который начинает процесс сокращения, «поднимая» молекулы тропомиозина с активных участков актиновых филаментов. Миозин в покое неактивен, так как на его головке находится отрицательно заряженный комплекс MgATФ, не позволяющий белку проявлять АТФ-азные свойства. После поступления ионов Са2+ происходит нейтрализация заряда на головке, что приводит миозин в возбужденное состояние. После этого миозиновые головки начинают прикрепляться к активным участкам актинового филамента. Когда миозиновая головка толстого филамента прикрепляется к тонкому филаменту, между толстым и тонким филаментами образуется поперечный мостик. При взаимодействии с актином каждая миозиновая молекула ежесекундно гидролизует до 10 молекул АТФ. За счет энергии, высвобождающейся при расщеплении АТФ, миозиновая головка поворачивается, что приводит к скольжению толстого и тонкого филаментов относительно друг друга (рис. 5.8). В конце гребка (поворота) к ми- озиновой головке присоединяется новая молекула АТФ, что приводит к отделению головки от актина и присоединению к новому активному участку тонкого филамента до тех пор, пока миозиновые головки не достигнут Z-диска. Так как при сокращении саркомера расстояние между Z-дисками уменьшается, происходит сокращение его длины. Одновременное сокращение всех саркомеров приводит к уменьшению длины миофибриллы и мышечного волокна. Ввиду того, что саркомер представляет собой не плоскую, а объемную структуру, при его сокращении происходит не только уменьшение его длины, но и увеличение его поперечного сечения (когда тоньше нити втягиваются в толстые), поперечного сечения мышечных волокон и всей мышцы.
Рис. 5.8. Схема, иллюстрирующая сокращение саркомера (сота) (JI. Страйер, 1985)
Прекращение нервного импульса приводит к расщеплению ацетил- холина и разрыву поперечных мостиков между актином и миозином. Благодаря действию «кальциевого насоса» ионы Са2+ возвращаются в саркоплазматический ретикулум, актин и миозин инактивируются, длина саркомера возвращается к исходному значению. Мышца расслабляется. Мышечное сокращение может продолжаться до тех пор, пока не истощатся запасы ионов кальция. Затем они снова перекачиваются в саркоплазматический ретикулум посредством активной системы «кальциевого насоса». Следует отметить, что для осуществления этого процесса необходима энергия АТФ. Каким образом доставляется энергия к филаментам? Кроме участка для прикрепления к тонкому филаменту, миозиновая головка содержит участок, в котором локализуется АТФ. Освобождающая вследствие реакции гидролиза (расщепления АТФ) энергия используется для прикрепления миозиновой головки к тонкому филаменту, а после осуществления поворота головки — для отделения миозиновой головки от тонкого филамента. КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Опишите состав и структуру миофибриллы. 2. Дайте характеристику состава и структуры саркомера. 3. Что такое сот? 4. Опишите состав и структуру толстого филамента. 5. Опишите состав и структуру тонкого филамента. 6. Опишите состав и структуру Z-диска. 7. Опишите состав и структуру М-диска. 8. Что представляет собой «теория скользящих нитей»? 9. Дайте характеристику параметрам, определяющим объем миофибриллы. 10. Опишите влияние тренировки на параметры миофибрилл.
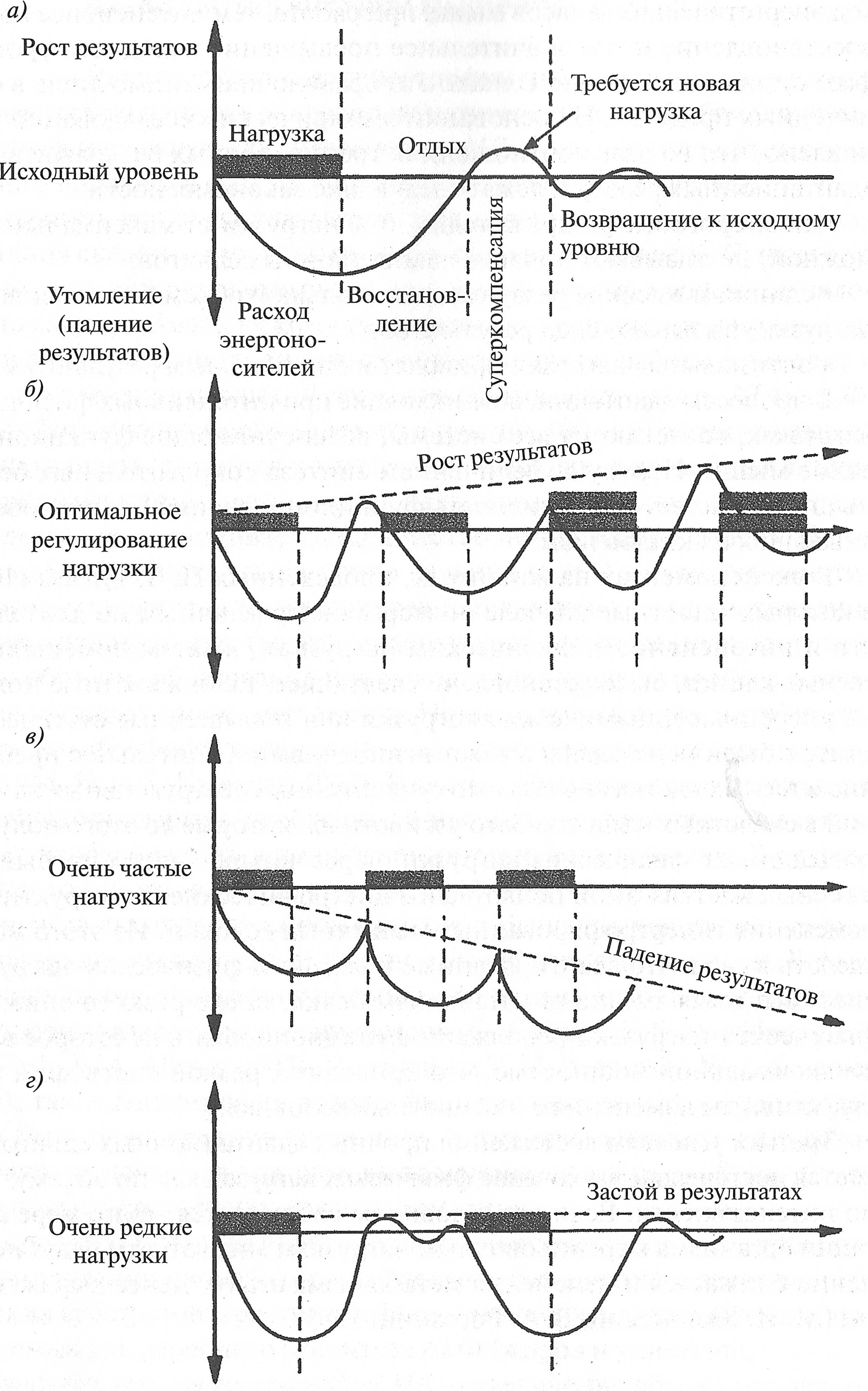
Условия адаптации Первым условием является многократное (повторное) применение физических нагрузок (П. К. Анохин, 1968; В. В. Васильева, 1969). Однократная физическая нагрузка не вызывает стойких адаптационных перестроек в организме. Если же физические нагрузки повторяются, в организме создается необходимый метаболический фон, который обеспечивает постепенность формирования морфологических, биохимических и функциональных изменений. При повторяющихся тренировочных нагрузках благодаря активации генетического аппарата мышечных волокон в мышцах увеличивается содержание структурных белков, вследствие чего мышцы становятся более резистентными к задаваемой нагрузке (М. И. Калинский, В. А. Рогозкин, 1989). Вторым условием, определяющим процесс адаптации организма к физическим нагрузкам, является их регулярное применение. Необходимость регулярно выполнять физические упражнения связана с изменениями метаболизма, которые происходят в организме в процессе физических нагрузок. В зависимости от интенсивности и длительности физической нагрузки в организме наблюдаются изменения в обмене веществ, которые могут быть ограничены локальными сдвигами в энергетическом обмене или затрагивать метаболизм всего организма. В последнем случае процесс восстановления метаболизма до уровня покоя занимает значительно больше времени и требует большего периода отдыха. Долговременная адаптация организма к различным факторам внешней среды возможна благодаря явлению суперкомпенсации (сверхвосстановления). В области мышечной деятельности это явление первыми описали JI. И. Ямпольский (1949) и Н. Н. Яковлев (1949, 1955). В основе суперкомпенсации лежит взаимодействие между нагрузкой и восстановлением. Это циклический процесс и его пусковым стимулом является физическая нагрузка. После нагрузки возникают утомление и резкое снижение работоспособности спортсмена, что соответствует первой фазе цикла. Во второй фазе начинается процесс восстановления работоспособности. К концу второй фазы работоспособность спортсменов достигает исходного уровня. В третьей фазе (фазе суперкомпенсации) работоспособность превышает исходный уровень. Последняя, четвертая фаза характеризуется возвращением работоспособности к исходному уровню (рис. 6.1, а).
Рис. 6.1. Принцип суперкомпенсации (Ю. Хартманн, X. Тюннеманн, 1988) Очень наглядно эффект суперкомпенсации виден на примере восстановления энергетических запасов мышц. Перед началом тренировки в мышцах находится определенное количество энергетических веществ (например, креатинфосфата, гликогена и др.). В результате тренировки происходит снижение уровня этих веществ в мышцах. После окончания тренировки, в фазе восстановления, уровень энергетических веществ в мышечном волокне превышает исходный, то есть происходит суперкомпенсация. Долговременная адаптация возможна только в том случае, если достигаемые срочный и отставленный тренировочные эффекты от каждой тренировки будут суммироваться (рис. 6.1, б). Поэтому для получения определенного тренировочного эффекта и последующего повышения физической работоспособности очередную физическую нагрузку следует проводить в период преимущественно суперкомпенсации после предшествующей работы. Слишком частые (рис. 6.1, в) тренировки прерывают стадию восстановления до достижения эффекта суперкомпенсации. Вследствие этого возможны постепенное развитие процесса недовосстановления работоспособности и преждевременное наступление утомления, что негативно сказывается на результатах (М. И. Калин- ский, В. А. Рогозкин, 1989). Слишком редкие тренировки (рис. 6.1, г) в фазе сниженной суперкомпенсации не позволяют закрепить тренировочный эффект, так как каждая последующая тренировка проводится после возвращения функциональных возможностей организма к исходному уровню. Однако еще в начале XX века М. Е. Маршак (1931) установил, что процессы восстановления после тяжелой мышечной работы в различных вегетативных системах протекают гетерохронию, то есть с разной скоростью. Более того, даже в пределах одной и той же системы для разных показателей ее функции возвращаются к уровню покоя не одновременно. В последующем эти данные были подтверждены для многих физиологических и биохимических показателей. Следует отметить, что процессы восстановления энергетических веществ в мышцах также протекают с разной скоростью и завершаются в разное время. Вначале из скелетных мышц и крови удаляется молочная кислота, которая окисляется до С02 или включается в синтез гликогена, затем происходит ресинтез креатинфосфата, гликогена и жиров. Существует правило Энгельгарда, согласно которому интенсивность протекания восстановительных процессов и сроки восстановления энергетических запасов организма зависят от интенсивности их расходования во время выполнения упражнения. Следовательно, чем больше рас-ход энергетических запасов мышц при работе, тем интенсивнее идет их восстановление и тем значительнее превышение исходного уровня в фазе суперкомпенсации. Однако это правило применимо лишь в ограниченных пределах. На основании эмпирических исследований установлено, что во взаимоотношениях тренировочных раздражителей и адаптационных реакций лежат следующие закономерности: · подпороговые раздражители (30 % нагрузки от максимально возможной) не вызывают никаких адаптационных сдвигов; · слишком высокие раздражители (большой объем и интенсивность нагрузки) вызывают спад результатов; · оптимальные нагрузки приводят к оптимальным результатам. В процессы адаптации, возникающие при интенсивных физических нагрузках, вовлекаются все системы, обеспечивающие функционирование мышц. Наряду с увеличением синтеза сократительных белков мышц возрастает интенсивность функционирования систем, обеспечивающих их катаболизм. В экспериментах на животных, проведенных П. 3. Гудзем (1963), в которых животные вначале подвергались различным по длительности и интенсивности физическим нагрузкам, а затем помещались в тесные клетки, было установлено следующее. Если животные получали умеренные динамические нагрузки или повышенные статические, а затем были переведены в режим гиподинамии, длительное пребывание в тесных клетках не вызвало существенных деструктивных изменений в скелетных мышцах. Зато у животных, которые до этого получали повышенные физические нагрузки, через четыре недели пребывания в тесных клетках были обнаружены дистрофические и деструктивные изменения гипертрофированных мышечных волокон. Из этого можно сделать вывод, что адаптированные к высоким физическим нагрузкам системы, отвечающие за катаболизм белка, после резкого снижения физических нагрузок продолжают функционировать некоторое время с максимальной мощностью, что приводит к резкой дистрофии и деструктивным изменениям в мышечных волокнах. Третьим условием достижения прочных адаптационных сдвигов является постепенное увеличение физических нагрузок как по объему, так и по интенсивности. Если это условие не соблюдается, то по мере адаптации организма к тренировочным нагрузкам энерготраты будут постепенно снижаться и изменения метаболизма станут менее выраженными (М. И. Калинский, В. А. Рогозкин, 1989). 6.4. Виды гипертрофии мышечных волокон Различают два вида гипертрофии мышечного волокна в ответ на физическую нагрузку — саркоплазматическую и миофибриллярную. Саркоплазматическая гипертрофия МВ — увеличение объема мышечного волокна за счет увеличения объема его саркоплазмы. Известно, что саркоплазматическая гипертрофия проявляется как адаптация скелетных мышц к тренировке на выносливость. Существует мнение, что пусковым стимулом увеличения объема мышечных волокон по саркоплазматическому типу является уменьшение в них источников энергии. Выполнение высокоинтенсивной работы приводит к тому, что в течение первых 10 секунд исчерпываются запасы АТФ и КФ. Если работа продолжается далее, то в течение 100-110 секунд подходят к концу запасы гликогена в мышечных волокнах. Анаэробный гликолиз приводит к тому, что в мышцах накапливается молочная кислота и, как следствие, увеличивается количество ионов водорода (Н+). Ионы водорода, ингибируя выход ионов кальция (Са2+) из саркоплазматического ретикулума, а также прикрепление ионов кальция к тропонину, уменьшают количество взаимодействующих поперечных мостиков. Таким образом, образование ионов водорода ухудшает процесс сокращения мышцы и уменьшает ее силу. Ионы водорода также ингибируют активность фосфофруктокиназы — ключевого гликолитического фермента (М. Foss, S. Kateyian, 1998). В связи с этим теряется возможность компенсации энергии, необходимой мышцам, за счет гликолиза и возникает необходимость в подключении тканевого дыхания. В результате постоянного истощения запасов АТФ, КрФ, гликогена, а также увеличения активности ионов кальция в мышечных волокнах развиваются процессы адаптации, называемые суперкомпенсацией. В фазе восстановления происходит поворот к большему анаболизму и снижению уровня катаболических процессов (G. Goldspink, S. Harridge, 2003; P. A. Tesh, В. A. Alkner, 2003). По мнению V. М. Zatsiorsky, W. J. Kraemer (2006), такая гипертрофия должна быть распространена у бодибилдеров и бегунов на средние дистанции, в тренировочные занятия которых часто включаются подходы, выполняемые «до отказа». В результате этого повышается устойчивость к утомлению. В процессе гипертрофической силовой тренировки вследствие суперкомпенсации происходит значительное увеличение запасов энергетических субстратов: креатинфосфата, гликогена, а также веществ, необходимых для аэробного ресинтеза АТФ (жиров и углеводов). Миофибриллярная гипертрофия MB — увеличение объема мышечного волокна за счет увеличения количества и объема миофибрилл. Миофибриллярная гипертрофия проявляется как адаптация организма спортсмена к силовым нагрузкам при направленности тренировочного процесса на увеличение силы мышц. Так как при этом типе гипертрофии возрастают количество и объем миофибрилл, это в конечном счете приводит к возрастанию числа поперечных мостиков при сокращении мышцы, а следовательно, и силы мышцы. Рис. 6.2. Степени повреждения мышечного волокна (R. L. Lieber, L. Е. Thornell, J. Frieden, 1996): а — напряжение мышцы ведет к повышению концентрации ионов кальция (черные кружки); б — повышение концентрации ионов кальция приводит к активации калпаинов и избирательному повреждению элементов цитоскелета; в — последующая активность мышцы повреждает цитоскелет
Иногда сильные повреждения обнаруживаются и в миофибриллах: некоторые саркомеры могут быть более растянутыми по сравнению с другими. Это означает, что поврежден цитоскелет мышечного волокна, то есть продольные филаменты, связывающие соседние саркомеры в одной миофибрилле, а также поперечные филаменты, связывающие соседние миофибриллы между собой и с сарколеммой. Повреждение ци- тоскелета неизбежно приводит к незначительным повреждениям сарколеммы и, как следствие, к активации деятельности клеток-сателлитов. Многочисленными исследованиями доказано, что в результате выполнения силовых упражнений в первую очередь повреждаются Z-диски, соединяющие саркомеры друг с другом. Напомним, что к Z-дискам прикрепляются тонкие филаменты. Считается, что Z-диски являются «слабым звеном» миофибриллы. При этом возможен как полный разрыв миофибрилл в области Z-дисков, так и растягивание Z-дисков с сохранением целостности миофибриллы (рис. 6.3). Возможно также повреждение М-дисков, к которым прикрепляются толстые филаменты (J. Frieden, R. L. Lieber, 1992). Так как миофибриллы в области Z-дисков соединены с сарколеммой посредством костамерных белков, происходят нарушение ее целостности и активация клеток-сателлитов.
Рис. 6.3. Электронная фотография мышечного волокна человека после выполнения эксцентрических упражнений (J. G. Yu, L. Carlsson, L. Е. Thomell, 2004): * — миофибриллы с разрушенными Z-дисками; О - миофибрилла с неповрежденным Z-диском Повреждение саркомеров и миофибрилл приводит также к повреждениям саркоплазматического ретикулума, нарушениям гомеостаза Са2+, что обусловливает активацию протеаз, играющих важную роль в запуске расщепления белков скелетных мышц, в воспалительных процессах и процессе регенерации. Теория повреждения позволяет объяснить, почему в первую очередь повреждаются мышечные волокна типа II. J. Frieden, R. L. Lieber (1992) предположили, что более сильные повреждения мышечных волокон типа II по сравнению с мышечными волокнами типа I связаны с тем, что в этих волокнах по сравнению с волокнами типа I Z-диски и М-диски значительно тоньше. Более тонкие Z-диски волокон будут легче повреждаться и, следовательно, запускать комплекс процессов, ведущих к повышенному синтезу белка в мышечных волокнах. Активация клеток-сателлитов позволяет объяснить, за счет каких ресурсов осуществляется повышенный синтез белка в мышечных волокнах. Этот ресурс — увеличение количества миоядер за счет деления клеток-сателлитов. Показано, что при силовой тренировке в мышечном волокне возрастает количество активных клеток-сателлитов и миоядер. Анализируя перестройки, происходящие в иннервации скелетных мышц у животных, подвергнутых высокоинтенсивным нагрузкам и работе до отказа, П. 3. Гудзь (1963) обращает внимание на то, что на утолщенных мышечных волокнах нервные окончания разрастаются, становятся шире. Зачастую на одном мышечном волокне образуются поля концевых пластинок, состоящие из 2—3 двигательных нервных окончаний. Это обеспечивает лучшую передачу нервных импульсов. Некоторые нервные окончания разрастаются вдоль мышечного волокна, образуя густую сеть терминалей. КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Сформулируйте понятия адаптации и стресса. 2. Опишите виды адаптации. 3. Какие виды тренировочного эффекта вы знаете? 4. Перечислите условия адаптации. 5. Дайте определения саркоплазматической и миофибриллярной гипертро- фий мышечных волокон. 6. Дайте определение и кратко опишите гипотезы миофибриллярной гипертрофии. 7. Опишите энергетическую гипотезу миофибриллярной гипертрофии мышечных волокон. 8. Опишите гипотезу ацидоза. 9. Опишите гипотезу гипоксии. 10. Дайте характеристику гипотезы механического повреждения мышечных волокон как основного стимула, приводящего к миофибриллярной гипертрофии.
Строение молекулы белка Белки — высокомолекулярные соединения, состоящие из аминокислот. В состав белковых молекул могут входить десятки, сотни и тысячи аминокислот, однако все белки синтезируются на основе 20 различных аминокислот. Аминокислоты, соединяясь друг с другом пептидной связью, образуют длинные неразветвленные цепи — полипептиды. Пептидные связи обладают высокой прочностью. Первый уровень пространственной организации белковой молекулы называется первичной структурой и представляет собой последовательность расположения аминокислот в полипептидных цепях. Каждый белок имеет уникальную первичную структуру (рис. 7.1).
Рис. 7.1. Структуры I—IV белковой молекулы Второй уровень пространственной организации — вторичная структура — задает пространственную форму полипептидных цепей. Выделяют три типа вторичной структуры: а-спираль, слоистоскладчатую спираль и коллагеновую спираль. При образовании а-спирали витки полипептидной цепи периодически повторяются. Слоистоскладчатая структура белка представляет собой линейные полипептидные цепи, расположенные параллельно и прочно связанные водородными связями. Такая структура является основой белка миозина. Коллагеновая спираль характеризуется сложной укладкой полипептидных цепей. Отдельные цепи спирализованы и закручены одна вокруг другой, образуя суперспираль. Такая структура характерна для белка коллагена, входящего в состав сухожилий мышц. Коллагеновая спираль обладает высокой жесткостью и прочностью. Третичная структура белка — это пространственная укладка спира- лизованных или линейных участков полипептидной цепи в виде фибриллы (нитевидная) или глобулы (шарообразная). В мышце фибриллярными белками являются коллаген (входит в состав сухожилий) и миозин. Эти белки не растворяются в воде. Глобулярным белком является сократительный белок мышц — актин (g-актин). Глобулярные белки растворяются в воде. Глобулярные белки могут превращаться в фибриллярные. Так, тонкие филаменты мышц образованы фибриллярным белком актином, который состоит из большого количества глобулярного актина. Четвертичной структурой обладают только некоторые белки. Четвертичная структура — это сложное надмолекулярное образование, состоящее из нескольких белков, имеющих свою собственную первичную, вторичную и третичную структуры. Связи белковых молекул в четвертичной структуре весьма слабые, поэтому они легко распадаются. Следует отметить, что из всех структур белка кодируется только первичная. Высшие структуры (вторичная, третичная и четвертичная) возникают самопроизвольно в соответствии со строением полипептидов. 7.3. Переваривание и всасывание белков Посредством питания организм получает необходимое ему количество белков. Установлено, что при потребности увеличения мышечной массы организм человека должен получать в день не менее 1, 5— 2 г белка на 1 кг массы тела. Расщепление пищевых белков начинается в желудке. В желудочном соке содержатся соляная кислота и большое количество протеолитиче- ских ферментов. Основным протеолитическим ферментом желудочного сока, расщепляющим белки в желудке, является фермент пепсин. Под воздействием соляной кислоты, содержащейся в желудочном соке, происходят набухание и частичная денатурация белков. Под воздействием протеолитических ферментов в пищевых белках расщепляются пептидные связи между аминокислотами, находящиеся в глубине их молекул. В результате такого воздействия белковые молекулы превращаются в смесь полипептидов различной длины. В двенадцатиперстной кишке образовавшиеся полипептиды подвергаются дальнейшему расщеплению при участии ферментов, содержащихся в соке поджелудочной железы и кишечном соке. В результате полипептиды расщепляются на более простые соединения — низкомолекулярные пептиды (олигопептиды) и аминокислоты. Окончательное расщепление низкомолекулярных пептидов до аминокислот происходит в тонком кишечнике под воздействием ферментов кишечного сока. Эти ферменты локализуются в микроворсинках кишечника, поэтому расщепление олигопептидов до аминокислот происходит именно там. Всасывание аминокислот в кишечнике осуществляется путем их транспорта через стенку кишечника. После этого аминокислоты по системе воротной вены поступают в печень, а затем — в большой круг кровообращения. К мышцам аминокислоты попадают посредством кровеносных капилляров, которыми окружено каждое мышечное волокно. Весь процесс переваривания белков в же- лудочно-кишечном тракте занимает в среднем 8—12 часов после принятия пищи и требует затрат энергии. Рис. 7.2. Этапы синтеза белка: а - транскрипция; б - трансляция Первый этап синтеза белка (транскрипция) протекает в миоядрах. Транскрипция — это процесс синтеза молекулы информационной РНК (иРНК) на участке молекулы ДНК (гене). Установлено, что транскрипция начинается с разрыва водородных связей между двумя цепями ДНК. Затем происходит «раскручивание» участка спирали ДНК. Специальный фермент — РНК-полимераза, — двигаясь по цепи ДНК, подбирает по принципу комплементарности нуклеотиды и соединяет их в цепочку. В результате на одной из цепей ДНК синтезируется молекула иРНК (рис. 7.2). Таким образом происходит переписывание (транскрипция) информации о структуре синтезируемого белка с участка ДНК. Затем иРНК выходит из ядра в саркоплазму мышечного волокна и перемещается в область рибосом. ДНК восстанавливает свою структуру. Доказано, что силовая тренировка приводит к увеличению количества миоядер (F. Kadi et al., 1999). Это увеличение связано с усиленным делением клеток-сателлитов при повреждении мышечного волокна. Возрастание количества миоядер создает предпосылки для увеличения синтеза белка, так как увеличивается количество иРНК, синтезируемой в ядрах мышечных волокон. Второй этап синтеза белка — рекогниция — проходит в саркоплазме мышечного волокна. В процессе рекогниции транспортные РНК (тРНК) соединяются с аминокислотами и транспортируют их в область рибосом. Так как существуют 20 основных аминокислот, то насчитывается более 20 видов тРНК.
Второй этап синтеза белка — рекогницию — называют еще этапом активации аминокислот. Установлено, что для каждой аминокислоты имеются свои специфические ферменты, которые участвуют в ее активации. Эти ферменты проявляют высокую активность в присутствии ионов магния (Mg2+). Молекулы тРНК имеют по два специфических триплета. Один из них — кодон, с которым связывается аминокислота, другой — антико- дон, который соответствует кодону данной аминокислоты в иРНК (см. рис. 7.2). Благодаря этому аминокислоты при синтезе белка располагаются в последовательности, диктуемой последовательностью кодонов виРНК. Третий этап синтеза белка — трансляция — происходит на рибосомах. Из аминокислот, перемещаемых тРНК в область рибосом, формируется пептидная цепь (белковая молекула) в соответствии с информацией, закодированной в иРНК. Молекула иРНК передвигается между двумя субъединицами рибосомы — малой и большой. К малой субъединице присоединяется иРНК, к большой — фермент, синтезирующий белок. При передвижении иРНК между двумя субъединицами рибосом кодо- ны иРНК взаимодействуют с антикодонами тРНК по принципу комплементарное™. Завершение синтеза белка обеспечивается кодонами терминации (стоп-сигналом) в иРНК, с которыми не может связаться ни одна тРНК. Поэтому процесс завершения синтеза белка называется тер- минацией. Затем включается фактор освобождения и полипептидная цепь белка отделяется от рибосом. Таким образом образуется первичная структура белка. Затем вновь синтезированный белок приобретает определенную пространственную вторичную структуру. Сложная третичная структура белка формируется самопроизвольно в саркоплазме и определяется характером первичной структуры белка, а также окружающими условиями. Аналогичным образом осуществляется синтез молекул основных сократительных белков (миозина и актина). Следует отметить, что синтез белка требует огромных затрат энергии. Так, только для присоединения одной аминокислоты к полипептидной цепи синтезируемого белка используется по меньшей мере пять молекул АТФ, поэтому процесс синтеза белка во многом зависит от скорости восстановления уровня АТФ в мышечных волокнах. Миофибриллогенез Под термином миофибриллогенез понимается формирование в мышечном волокне специальных органелл — миофибрилл. Итак, после третьего этапа синтеза сократительных белков в мышечном волокне синтезируются молекулы различных белков: миозина, акти- * Триплет — комбинация из трех последовательно расположенных нуклеотидов в молекуле нуклеиновой кислоты. на, тропомиозина, тропонина и многих других. Каким же образом из этих молекул формируется структурная единица саркомера, состоящая из одного толстого и шести тонких филаментов? Каким образом увеличивается число миофибрилл, а также площадь их поперечного сечения? Установлено, что толстые и тонкие филаменты формируются путем самосборки. В растворах слабой ионной силы молекулы миозина начинают агрегировать (собираться) в более сложные структуры — толстые филаменты. При этом миозиновые молекулы объединяются «хвост в хвост» и образуются биполярные нити. Этот процесс продолжается за счет добавления молекул миозина на каждом конце нити до тех пор, пока их длина не достигнет 1, 5 мкм. К этому времени они становятся похожими на толстые филаменты. Следует заметить, что процесс образования такой сложной структуры, как толстый филамент, очень чувствителен к показателю рН. Вне области физиологических значений рН (7, 2—7, 5) агрегация (самосборка) часто происходит нерегулярно, так что возникают разветвляющиеся структуры и очень длинные агрегаты (Дж. Бендолл, 1970). Из этого следует, что смещение рН в мышцах в кислую сторону, что имеет место при накоплении молочной кислоты в мышцах, отодвигает сборку миофибрилл до того момента, пока рН мышц не достигнет физиологических значений. Тонкие филаменты также формируются путем самосборки. После того как на рибосомах синтезируется полипептидная цепь белка актина, она автоматически сворачивается в G-форму, которая представляет собой третичную структуру белка. При этом молекулы глобулярного актина (G-актина) в присутствии АТФ, ионов магния (Mg2+) и калия (К+) полимеризуются в длинные нити фибриллярного актина (F-актина), составляющего основу тонкого филамента. После этого нити фибриллярного актина скручиваются в двойную спираль. Если в растворе находятся толстые и тонкие филаменты, то они вступают друг с другом в связь и возникает так называемый актомио- зиновый комплекс {сот). В результате взаимодействия между толстыми и тонкими филаментами возникают структуры с периодическим чередованием толстых и тонких филаментов, сходные с натуральными по- перечно-полосатыми миофибриллами. При этом значительно повышается вязкость раствора. В 40-х годах XX века Альберт Сент-Дьерди показал, что возрастание вязкости уменьшается, если в раствор добавить молекулы АТФ. Было доказано, что добавление АТФ в раствор вызывает диссоциацию акто- миозинового комплекса на актин и миозин. Если же актомиозиновый комплекс помещался в раствор, содержащий ионы калия, магния и молекулы АТФ, актомиозиновые нити сокращались. В тех же условиях нити, образованные только из миозина, не сокращались. Эти опыты позволили установить, что мышечное сокращение возникает в результате взаимодействия миозина, актина и АТФ (Л. Страйер, 1985). На последней стадии морфогенеза миофибриллярного аппарата приводится в соответствие поперечная исчерченность соседних миофибрилл (Е.К. Жуков, 1974), то есть формируется цитоскелет мышечного волокна. Ответственными за поперечную исчерченность являются цитоскелетные белки, к одним из которых относится десмин. Этот белок связывает Z-диски соседних миофибрилл. Показано, что в процессе регенерации миофибрилл после эксцентрических упражнений увеличивается синтез десмина и происходит перестройка цитоскелета мышечного волокна (J. Friden, U. Kjorell, L.E. Thornell, 1984; J. Frieden, R.L. Lieber, 1992). Таким образом, общая картина морфогенеза миофибриллярного аппарат следующая. На первой стадии происходит сборка исходных белковых составных частей в неисчерченные нитевидные структуры миофиламентов. На второй стадии осуществляется распределение образовавшихся компонентов в поперечно-полосатые структуры — миофибриллы. На третьей стадии происходит приведение в соответствие расположение саркомеров в соседних миофибриллах (Е. К. Жуков, 1974). При рассмотрении параметров, влияющих на поперечное сечение мышечного волокна, было показано, что его увеличение может осуществляться за счет миофибриллярной гипертрофии — возрастания количества миофибрилл и их объема. Теория повреждения миофибрилл позволяет объяснить этот феномен. В детском возрасте миофибриллы «растут» с увеличением длины конечностей (J. D. MacDougall, 2003). Так как при росте конечностей мышца находится в постоянном растяжении, миофибриллы постепенно увеличивают свою длину. Это происходит за счет того, что на концах миофибрилл осуществляется их «достройка» посредством сборки дополнительных саркомеров. Таким способом увеличивается длина миофибрилл. Возможен также полный поперечный разрыв миофибриллы. При этом каждая «культя» увеличивает свою длину посредством достройки (сборки дополнительных саркомеров). В результате вместо одной миофибриллы получаются две.
Рис. 7.3. Продольное расщепление миофибриллы двуглавой мышцы плеча мыши: а — начало расщепления миофибриллы в области двух Z-дисков; б — расщепленные миофибриллы (G. Goldspink, 1971) Доказано также продольное расщепление миофибрилл. Это происходит тогда, когда объем миофибриллы (или ее поперечное сечение) становится очень большим и достигает критических значений. Чаще всего расщепление миофибриллы начинается в области Z-дисков (рис. 7.3). При этом в центре диска появляется небольшое отверстие, которое затем расширяется. В результате образуются две «дочерние» миофибриллы (G. Goldspink, 1970, 1971). Причину продольного расщепления миофибриллы G. Goldspink (1971) видит в том, что тонкие филаменты прикрепляются к Z-диску под небольшим углом. При развитии напряжения мышцы этот угол увеличивается, сила тяги тонких филаментов возрастает и повреждает Z-диск. Если напряжение мышцы происходит очень быстро, что имеет место в быстрых мышечных волокнах, Z-диски повреждаются еще быстрее и чаще происходит расщепление миофибрилл. Поэтому, как считает G. Goldspink (1971), миофибриллы в быстрых мышечных волокнах, как правило, небольшие, в то время как в медленных — более крупные. Новые миофибриллы могут возникать в результате деления и объединения клеток сателлитов. Таким способом может увеличиваться число миофибрилл в мышечном волокне. Поперечное сечение миофибрилл возрастает за счет добавления синтезированных толстых и тонких филаментов на их поверхности (Е. К. Жуков, 1974; J. D. MacDougall, 2003). Однако плотность упаковки миофиламентов не возрастает, расстояние между толстым и тонкими филаментами не меняется. Так как увеличение количества миофибрилл и их объема влияет на объем мышечного волокна, возникает гипертрофия мышечных волокон по миофибриллярному типу. КОНТРОЛЬНЫЕ ВОПРОСЫ 1. Укажите основные функции ДНК и РНК. 2. Какие виды РНК вы знаете? 3. Сколько полинуклеотидных цепей в ДНК и РНК? 4. Что представляет собой ген? 5. Что представляют собой первичная, вторичная, третичная и четвертичная структуры белка? 6. Охарактеризуйте процессы переваривания и всасывания белков. 7. Как протекает катаболизм белков в мышечных волокнах? 8. Что представляет собой антикатаболический эффект? 9. Дайте краткую характеристику этапам синтеза белка в MB. 10. Опишите первый этап синтеза белка — транкрипцию. 11. Опишите второй этап синтеза белка — рекогницию. 12. Опишите третий этап синтеза белка — трансляцию. 13. Что представляет собой миофибриллогенез? 14. Дайте характеристику гиперплазии у животных и человека. 15. Опишите механизмы возникновения новых мышечных волокон в скелетной мышце. 16. Опишите концепции, объясняющие повышенный синтез белка в скелетных мышцах человека при выполнении силовых тренировок. 17. Как влияют занятия силовыми упражнениями на размер миоядерного домена?
Рис. 8.1. Взаимоотношения между различными механизмами увеличения мышечной силы (J. Hoffman, 2002) На этапе, соответствующем среднему уровню спортивного мастерства, «вклад» морфометрических показателей (площади поперечного сечения мышц) в максимальную силу мышц увеличивается. Однако по мере роста мастерства увеличение силы и площади поперечного сечения мышц замедляется.
Рис. 8.2. Развитие максимальной силы за счет улучшения внутримышечной координации (метод кратковременных максимальных напряжений) (Ю. Хартманн, X. Тюннеманн, 1988) На этапе высшего спортивного мастерства прирост силы и гипертрофия мышц резко уменьшаются, поэтому, чтобы увеличить силовые и морфометрические показатели мышц, некоторые спортсмены прибегают к приему анаболических стероидов. По данным S. Е. Alway et al. (1992), у бодибилдеров, имеющих стаж тренировочных занятий не менее пяти лет, не обнаружено достоверных изменений в силовых и морфометрических показателях двуглавой мышцы плеча после двух лет тренировки (табл. 8.1). Таблица 8.1 Влияние занятий бодибилдингом в течение двух лет на морфометрические показатели двуглавой мышцы плеча элитных бодибилдеров (S. Е. Alway et ah, 1992)
Рис. 8.3. Степень проявления запаздывающих болезненных ощущений после выполнения эксцентрических упражнений (Р. М. Clarkson, I. Tremblay, 1988) Существуют несколько гипотез о причинах появления запаздывающих болезненных ощущений в мышцах: локальный спазм мышцы (ишемия); накопление в мышцах продуктов распада (в частности, молочной кислоты); механические повреждения мышечных волокон и соединительной ткани. Н. A. deVriez (1966) выдвинул теорию «спазма» (ишемии) на основе зарегистрированной им повышенной электрической активности в расслабленных мышцах, в которых имеются запаздывающие болезненные ощущения. Согласно этой теории, последовательность событий, приводящих к DOMS, такова. После тяжелых для организма упражнений в мышцах возникает локальная ишемия, которая вызывает боль. Боль приводит к увеличению активности мышц, росту напряжения и еще большему увеличению ишемии. Однако теория «спазма» не получила подтверждения в последующих исследованиях. Вторая популярная гипотеза связывает DOMS с накоплением молочной кислоты в мышцах. Накопление в мышцах молочной кислоты происходит в результате функционирования мышцы в анаэробных условиях (гликолитический путь ресинтеза АТФ). Накопление молочной кислоты в саркоплазме мышечных волокон вызывает изменение осмотического давления. Вода из тканевой жидкости поступает внутрь мышечных волокон, вызывая их набухание. В процессе набухания происходят сдавливание нервных окончаний и, как следствие, возникновение болезненных ощущений (М. И. Калинский, В. А. Рогозкин, 1989; В. И. Морозов, Г. А. Сакута, М. И. Калинский, 2006). Согласно этой гипотезе, появление в крови креатинкиназы интерпретируется как увеличенная проницаемость или разрушение мембраны мышечного волокна вследствие деструкции липидов под действием перекисного окисления. Согласно третьей гипотезе, основной причиной возникновения запаздывающих болезненных ощущений в мышцах являются механические повреждения мышечных волокон. Еще в 50-х годах прошлого века Е. Асмуссен (Е. Asmussen, 1952, 1956) установил, что запаздывающие болезненные ощущения непосредственно связаны с эксцентрическим компонентом упражнений. В настоящее время доказано (М. J. Gibala et al., 1995), что при выполнении силовых упражнений происходит повреждение мышечных волокон. Более сильные повреждения наблюдаются при сокращении мышцы в эксцентрическом режиме. Механические повреждения мышечных и коллагеновых волокон приводят к запуску целого ряда процессов дегенерации и регенерации мышечной и соединительной тканей, в результате чего человек испытывает болезненные ощущения. Установлено, что вначале боль локализуется в проксимальных и дистальных сухожильно-мышечных соединениях, затем распространяется на всю мышцу. Это позволяет предположить, что ультраструктурные повреждения локализуются прежде всего в дистальных и проксимальных частях мышцы. Согласно гипотезе механического повреждения, понижение уровня силы мышц связано с повреждением большого количества мышечных волокон. Появление в крови креатинкиназы обусловлено с разрушением мембраны мышечного волокна вследствие ее механического повреждения (J. Frieden, R. L. Lieber, 2001). Появление в крови легкой изоформы миозина связано с большим повреждением мышечных волокон типа II (М. Guerrero et al., 2008). Следует отметить, что гипотеза о связи запаздывающих болезненных ощущений с накоплением в мышцах продуктов катаболизма (например, молочной кислоты) также не отвергается. Предполагается, что начальная фаза повреждения — механическая, за которой следует вторичное метаболическое повреждение. Молочная кислота стимулирует нервные окончания, что ощущается человеком как боль. Кроме того, увеличение в мышечных волокнах молочной кислоты вызывает их набухание и снижение рН саркоплазмы. Не исключается, что недостаток кислорода и повышенная кислотность в мышечных волокнах способны стимулировать дегенеративные процессы, возникающие в результате их повреждения. Рис. 8.5. График вовлечения в работу медленных и быстрых мышечных волокон (Дж. X. Уилмор, Д. Л. Костилл, 1997) Если в одном сете выполняются многократные повторения упражнений с небольшим отягощением (менее 50 % от максимально возможного), рекрутируются только ДЕ типа S, в состав которых входят мышечные волокна типа I. Эти мышечные волокна способны сокращаться длительное время. При этом уровень силы, развиваемый мышцей, будет небольшим, так как отягощения небольшие. Для преодоления субмаксимальных или максимальных отягощений мышца должна развить наибольшую силу. Поэтому в сокращение последовательно вовлекаются все типы мышечных волокон, особенно типа IIВ (рис. 8.5). Нетренированные люди не способны вовлекать в работу достаточно большое количество мышечных волокон. С повышением уровня мастерства улучшается управление мышцами, поэтому количество вовлеченных в работу мышечных волокон значительно увеличивается. Мышечные волокна типа IIB, вносящие максимальный вклад в поперечное сечение мышцы, повреждаются в первую очередь, так как у этих волокон по сравнению с волокнами типа I более тонкие Z- и М-диски. При этом происходит растяжение или разрыв миофибрилл. Повреждение миофибрилл в волокнах типа IIВ приводит к запуску процессов регенерации, следствием которой является миофибриллярная гипертрофия мышечных волокон. Рис. 8.6. Изменение максимума силы мышц, измеренной в изометрическом режиме 7, уровня 2 запаздывающих болезненных ощущений и уровня 3 креатинкиназы в плазме крови после выполнения серии эксцентрических упражнений мышцами- сгибателями предплечья (J. A. Faulkner, S. V. Brooks, J. A. Opiteck, 2. Тренировка в динамическом (концентрическом и эксцентрическом) режиме приводит к удлинению мышечных волокон и укорочению сухожилий, возрастанию количества миофибрилл и ядер овальной формы, увеличению концевых пластинок. Как показало компьютерное моделирование (U. Proske, D. L. Morgan, 2001), увеличение длины мышечных волокон осуществляется за счет возрастания количества саркомеров, что приводит к изменению оптимальной длины мышцы при развитии напряжения. 3. Тренировка в динамическом режиме приводит к большей гипертрофии мышц по сравнению с изометрическим режимом (P. J. Rasch и L. J. Morehouse, 1957). 4. Тренировка в эксцентрическом режиме приводит к несколько большей гипертрофии и жесткости мышц по сравнению с тренировкой в концентрическом режиме (Е. Higbie et al., 1996). 5. Гипертрофия мышечных волокон при работе в эксцентрическом режиме больше, чем при концентрическом, при этом гипертрофия быстрых мышечных волокон при выполнении быстрых эксцентрических сокращений больше, чем медленных. 6. Растягивание пассивной мышцы не приводит к ее гипертрофии (R. Fowles et al., 2000). Рис. 8.9. Группы ДЕ, активированных во время выполнения силовых упражнений при поднятии субмаксимальных отягощений (V. М. Zatsiorsky, W. J. Kraemer, 2006) 1 — ДЕ рекрутированные, но не утомленные; 2 — ДЕ рекрутированные и утомленные (тренировочный коридор); 3 — ДЕ нерекрутированные Согласно гипотезе V. М. Zatsiorsky, W. J. Kraemer (2006), если ДЕ не утомлены, они не тренируются, поэтому воздействие на медленные мышечные волокна, входящие в первую группу, посредством этого метода минимально. Вторую группу составляют ДЕ, которые активированы и сильно утомлены (истощены). По мнению V. М. Zatsiorsky, W. J. Kraemer (2006), только эти ДЕ испытывают тренировочное воздействие. В этой группе не представлены медленные ДЕ, а также сильные и быстрые. В третью группу входят самые быстрые и высокопороговые ДЕ, которые не активированы и, следовательно, не испытывают тренировочного воздействия. Картина активности ДЕ в стандартном цикле представлена на рис. 8.9.
Рис. 8.10. Группы ДЕ, активированных во время выполнения силовых упражнений: а — отказной цикл; б — при методе максимальных усилий (Y. М. Zatsiorsky, W. J. Kraemer, 2006) 1—3 — те же, что и на рис. 8.9 В последнем, «отказном» цикле картина меняется. К этому времени рекрутируется большое количество ДЕ, в том числе и самые большие ДЕ, которые не были активированы в стандартных циклахРекрутирование (рис. 8.10). «Если атлет может поднять штангу 12 раз, но поднимает только 10 — упражнение бесполезно для тренировки самых больших ДЕ. Эти ДЕ не активируются в этом подходе, так как они активируются только в последних двух подходах, которые не выполняются» (V. М. Zatsiorsky, W. J. Kraemer, 2006. С. 85). Однако концепция, предлагаемая V. М. Zatsiorsky, W. J. Kraemer (2006), не учитывает факта повреждения MB во время выполнения упражнения.Гкпотеза, учитывающая повреждение мышечных волокон. На основе анализа экспериментальных данных (А. В. Самсонова, 2011) была высказана гипотеза, учитывающая, что при выполнении силовых упражнений происходит повреждение мышечных волокон. Если работа «до отказа» выполняется с небольшим внешним отягощением, например 40 % от максимума, то к последнему, «отказному» циклу большая часть мышечных волокон дезактивируется из-за уменьшения запасов энергии и понижения активности MB типа II в кислой среде (в мышцах накапливается много молочной кислоты). Работа в таком режиме в большей степени ведет к развитию силовой выносливости, чем силы. Гипертрофируются в первую очередь медленные мышечные волокна типа I, при этом имеет место саркоплазматическая гипертрофия мышечных волокон. Работа «до отказа» с внешней нагрузкой 70—80 % от максимума (жим штанги ногами лежа на тренажере) выглядит следующим образом. При выполнении жима штанги ногами лежа на тренажере, спортсмен на выполнение восьми циклов движений с 80%-ным отягощением затрачивает в среднем 20—30 с. При выполнении каждого стандартного цикла, начиная с первого, при котором мышца попеременно работает в эксцентрическом и концентрическом режимах, небольшая часть ДЕ (а следовательно, и мышечных волокон) прекращает функционировать из-за их повреждения. Следует отметить, что при работе с нагрузкой, составляющей 80 % от максимальной, активны все типы ДЕ. Чтобы движение выполнялось в том же темпе и мышца продолжала развивать необходимое усилие, требуется постоянная активация дополнительных ДЕ, до этого не принимавших участия в работе. Перед окончанием работы «до отказа» в предпоследнем и последнем циклах количество мышечных волокон, способных развивать необходимое усилие, резко уменьшается — частично из-за повреждения, а также из-за того, что к 20—30 с запасы КрФ в мышце резко уменьшаются. Поэтому ЦНС в последних циклах движения «бросает в бой» свой резерв — самые большие и сильные ДЕ, которые еще не принимали участие в работе и потому сохранили достаточное количество КрФ. Этим можно объяснить увеличение амплитуды ЭМГ в предпоследнем или последнем цикле. Однако падение силы к последнему, «отказному» циклу в мышце из-за повреждения и истощения большого количества ДЕ таково, что мышца уже не способна развить требуемый импульс силы (произведение силы мышцы на длительность ее активности), чтобы сообщить внешнему отягощению необходимую скорость после его остановки в нижнем положении. Так как уровень силы, развиваемый мышцей, резко падает, накопление необходимого импульса силы происходит за счет значительного увеличения длительности «отказного» цикла. При этом скорость сокрн щения мышцы в концентрическом режиме резко уменьшается, и мыш ца начинает функционировать практически в изометрическом режиме, который, по сравнению с концентрическим, как было показано выше, обладает большим повреждающим воздействием на мышечные волок на. Поэтому при медленном выполнении последнего, «отказного» цик ла происходит значительное повреждение большого количества мы шечных волокон, подобно тому как под действием огромной внешней нагрузки, привносимой ураганом, постепенно рвутся стальные жилы канатов Байтового моста. К концу отказного цикла оставшиеся активными ДЕ уже не способ ны развить необходимый импульс силы, чтобы сообщить огромному внешнему отягощению (80 % от максимума) нужную скорость. Однако развиваемый мышцей импульс силы достаточен для того, чтобы обе спечить прыжок вверх с места, что подтверждается фактами, полученными Е. Б. Мякинченко (1997). При этом спортсмен испытывает силь нейшие болезненные ощущения из-за резкого возрастания в мышцах уровня лактата, происходит «отказ» от выполнения упражнения. Через несколько минут отдыха в части истощенных, но не поврежденных мышечных волокон запасы креатинфосфата частично восстанавливаются и они могут снова развивать достаточное усилие. Поэтому спортсмен способен снова повторить подход «до отказа». Но так как часть мышечных волокон уже повреждена, количество циклов в подходе «до отказа» уменьшается. КОНТРОЛЬНЫЕ ВОПРОСЫ 4. Опишите изменение силы и гипертрофии скелетных мышц на начальном этапе силовых тренировок. 5. Опишите изменение силы и гипертрофии скелетных мышц на этапе, со ответетвующем среднему уровню спортивного мастерства. 6. Опишите изменение силы и гипертрофии скелетных мышц на этапе высшего спортивного мастерства. 7. Опишите причины острых и запаздывающих болезненных ощущений. 8. Объясните механизм влияния различных отягощений на силу и гипертрофию скелетных мышц. 9. Опишите срочные тренировочные эффекты воздействия на силу и гипертрофию скелетных мышц различных режимов их функционирования. 10. Опишите отставленные тренировочные эффекты воздействия на силу и гипертрофию скелетных мышц различных режимов их функционирования. 11. Опишите кумулятивные тренировочные эффекты воздействия на силу и гипертрофию скелетных мышц различных режимов их работы. 12. Дайте характеристику гипотезам, объясняющим большее повреждение мышечных волокон при работе в уступающем (эксцентрическом) режиме мышечного сокращения. 13. Охарактеризуйте достоинства и недостатки метода «до отказа». 14. Опишите срочные тренировочные эффекты воздействия на силу и гипертрофию скелетных мышц метода «до отказа» с различными отягощениями. 15. Опишите отставленные тренировочные эффекты воздействия на силу и гипертрофию скелетных мышц метода «до отказа» с различными отягощениями. 16. Опишите кумулятивные тренировочные эффекты воздействия на силу и гипертрофию скелетных мышц различных режимов их функционирования. 17. Дайте характеристику гипотезам, объясняющим значительное повреждение мышечных волокон при использовании метода «до отказа». 18. Охарактеризуйте влияние прекращения тренировки и последующего ее возобновления на гипертрофию скелетных мышц. 19. Охарактеризуйте влияние порядка упражнений и различных протоколов тренировки на силу и гипертрофию скелетных мышц.
ГЛОССАРИЙ Адаптация — структурно-функциональная перестройка организма, позволяющая обеспечить его жизнеспособность в изменяющихся условиях и представляющая собой процесс адекватного приспособления организма к окружающей среде. Аденозинтрифосфат (АТФ) — основное макроэргическое соединение организма. В условиях организма при расщеплении одного моля АТФ (506 г) выделяется 10-12 ккал (42-50 кДж). Актин — основной белок, входящий в состав тонкого филамента. Аминокислоты — обязательные составные части белков. Анатомический поперечник — площадь, соответствующая разрезу, перпендикулярному длине мышцы. Атрофия — уменьшение объема органа под влиянием различных факторов. Ацетилхолин — нейромедиатор, который выделяют нервные окончания в синаптическую щель при поступлении по аксону мотонейрона нервного импульса. Ацидоз — повышение кислотности крови, сопровождающееся снижением значения рН и обусловленное накоплением органических кислот. При напряженной мышечной работе ацидоз приводит к развитию утомления. Аэробный путь ресинтеза АТФ (синонимы: тканевое дыхание, аэробное или окислительное фосфорилирование) — основной способ образования АТФ с использованием различных энергетических источников (жиров, углеводов, аминокислот и др.) при участии кислорода. Базалъная мембрана — внешняя оболочка мышечного волокна. Биопсия мышцы (от греч, bios — жизнь, opsis — вид, зрелище) — извлечение крошечного кусочка ткани из брюшка мышцы в целях последующего его исследования. Вторичная структура белка — пространственная форма полипептидных цепей. Ген — участок молекулы ДНК, содержащий информацию об одном белке. Гидролиз АТФ — химическая реакция взаимодействия АТФ с водой, в результате которой выделяется энергия. Гиперплазия миофибрилл — увеличение количества миофибрилл в мышечном волокне. Гиперплазия мышечных волокон — увеличение количества мышечных волокон в мышце. Гипертрофия — увеличение объема органа под воздействием различных факторов. Гипертрофия мышечного волокна — увеличение объема мышечного волокна под воздействием различных факторов. Гипоксия — состояние кислородного голодания тканей. Гистогенез (от греч. histos — ткань и...генез — развитие тканей) — совокупность закономерно протекающих процессов, обеспечивающих возникновение, существование и восстановление тканей животных организмов с их специфическими в разных органах свойствами. Гликолитический путь ресинтеза АТФ (гликолиз) — анаэробный путь ресинтеза АТФ, использующий гликоген, содержащийся в мышечных волокнах, и глюкозу крови. Двигательная единица (ДЕ) — функциональная система, состоящая из мотонейрона и иннервируемых им мышечных волокон. ДЕтипа S (slow) — медленная, устойчивая к утомлению ДЕ. ДЕтипа FR (fast resistant)— быстрая, устойчивая к утомлению ДЕ. ДЕтипа /^(fast fatigable) — быстрая, быстроутомляемая ДЕ. Дифференцировка — приобретение клеткой определенной структуры для выполнения конкретной функции. Долговременная адаптация — структурно-функциональная перестройка, происходящая в организме в ответ на длительное или многократное воздействие физической нагрузки. Изометрический режим сокращения мышцы — режим сокращения, при котором длина мышцы не изменяется. Инвазивные методы оценки композиции MB — повреждающие (прямые) методы оценки композиции мышц (биопсия с последующим гистохимическим исследованием мышечных волокон). Запаздывающие болезненные ощущения (DOMS) — мышечные боли, возникающие через один—три дня после изнурительной физической нагрузки. Катаболизм белков (протеолиз) — расщепление белков под воздействие различных факторов. Катепсины — ферменты, расщепляющие белки в мышечных волокнах. Клетки-сателлиты — стволовые клетки, расположенные между ба- зальной мембраной и сарколеммой мышечного волокна. Кодон (триплет) — сочетание трех азотистых оснований, последовательно расположенных в молекуле ДНК, соответствующее определенной аминокислоте. Комплекс Голъджи — мембранная органелла мышечного волокна, имеющая вид плоских цистерн, на периферии которых имеются многочисленные мелкие пузырьки. Композиция мышечных волокон — процентное соотношение в мышце волокон различных типов. Компьютерная томография — метод неразрушающего послойного исследования внутренней структуры объекта, основанный на измерении и сложной компьютерной обработке степени ослабления рентгеновского излучения различными по плотности тканями. Конституция человека — совокупность функциональных и морфологических особенностей организма, сложившихся на основе наследственных и приобретенных свойств, которые определяют своеобразие реакции организма на внешние и внутренние раздражители. Концевая пластинка — окончание нейрона, которое выполняет функцию контакта с мышечным волокном. Креатин — вещество, синтезирующееся из аминокислот. При взаимодействии с АТФ креатин превращается в креатинфосфат. Креатинфосфат (КрФ) — макроэргическое соединение организма. Креатинфосфатный путь ресинтеза АТФ — анаэробный путь ресин- теза АТФ, использующий креатинфосфат (КрФ), содержащийся в мышечных волоьснах. Кумулятивный тренировочный эффект выражается через показатели, характеризующие долговременную адаптацию организма спортсмена к тренировочной нагрузке. Лизосомы — мембранные органеллы мышечного волокна, представляющие собой микроскопические пузырьки размером 0, 2—0, 4 мкм. В лизосомах содержатся гидролитические ферменты, расщепляющие белки, нуклеиновые кислоты и некоторые полисахариды. Магниторезонансная томография (МРТ) — метод исследования внутренних органов и тканей человека посредством физического явления ядерного магнитного резонанса. Мезенхима — эмбриональная соединительная ткань, источник происхождения клеток всех соединительных тканей. Метаболизм — совокупность химических реакций, протекающих во внутренней среде организма. Миозин — белок, из которого состоит толстый филамент. Миотом — слой клеток, из которого впоследствии образуются скелетные мышцы. Миотонометрия — методика неинвазивной оценки композиции MB, разработанная М. С. Цветковым. Миосимпласт — мышечное волокно. Миофибриллогенез — формирование в мышечном волокне специальных органелл — миофибрилл. Миофибриллы — органеллы специального назначения, представляющие собой тонкие нити, идущие вдоль мышечного волокна. Миофибриллярная гипертрофия мышечного волокна — увеличение объема мышечного волокна за счет увеличения количества и объема миофибрилл. Миоядерный (мионуклеарный) домен (DNA-unit) — объем мышечного волокна (саркоплазмы), контролируемый одним ядром. Митохондрии — мембранные органеллы мышечного волокна размером до 2—3 мкм. В митохондриях протекает окисление углеводов, жиров и аминокислот до углекислого газа и воды с использованием кислорода воздуха. За счет энергии, выделяющейся при окислении, в митохондриях осуществляется синтез АТФ. Моль — единица измерения количества вещества. 1 моль равен количеству вещества, в котором содержится NA частиц. NA — постоянная Авогадро. NA = 6, 02214179x1023. М-ответ — суммарный электрический потенциал в ответ на одиночное раздражение двигательного или смешанного нерва. Мотонейрон {лат. motor — приводящий в движение + нейрон; синоним: нейрон двигательный) — крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса. Мотонейронный пул — совокупность мотонейронов, иннервирую- щих одну мышцу. Мочевина — конечный продукт обезвреживания аммиака, образующегося при распаде аминокислот (белков). Мышечные волокна типа /(MB типа I) — красные, медленные, устойчивые к утомлению окислительные мышечные волокна. Мышечные волокна типа IIA (MB типа Па) — промежуточные, быстрые, устойчивые к утомлению окислительно-гликолитические мышечные волокна. Мышечные волокна типа IIB (MB типа IIB) — белые, быстрые, бы- строутомляемые, гликолитические, анаэробные мышечные волокна. Неинвазивные методы оценки композиции MB — непровреждающие (косвенные) методы. Нейромедиатор — биологически активное химическое вещество, посредством которого осуществляется передача электрического импульса от мотонейрона к мышце. Олигопептид — продукт расщепления белков (пептидов). Органеллы общего назначения — обязательные элементы любой клетки. Острые болезненные ощущения — мышечные боли, возникающие в процессе и сразу после физической нагрузки. Отставленный тренировочный эффект выражается через различные показатели, характеризующие изменения в организме спортсмена в ближайшие дни после тренировки. Пепсин — основной протеолитический фермент желудочного сока, расщепляющий белки в желудке. Первичная структура белка — последовательность расположения аминокислот в полипептидных цепях. Перимизий — соединительно-тканная оболочка, окружающая пучки мышечных волокон. Площадь поперечного сечения мышцы (SM) — площадь фигуры, полученная при поперечном разрезе мышцы в области, соответствующей ее максимальному обхвату. Полипептиды — длинные неразветвленные цепи, состоящие из остатков аминокислот, соединенных пептидными связями. Потенциал действия — волна возбуждения, перемещающаяся по мембране мышечного волоьсна. Преинкубация препарата — выдерживание препарата в кислой или щелочной среде для определения композиции MB гистохимическими методами. Преодолевающий (концентрический) режим сокращения мышцы — режим, при котором длина мышцы уменьшается. Принцип размера (правило Хенеманна) — правило, согласно которому активируются ДЕ при развитии мышцей напряжения. Согласно принципу размера, при развитии мышцей напряжения вначале активируются ДЕ типа S, затем ДЕ типа FR, после чего ДЕ типа FF. Проприорецепторы — специальные образования опорно-двигательного аппарата, информирующие нервную систему о длине, скорости сокращения мышцы, а также о ее напряжении. Протеолиз — распад белков, приводящий к образованию аминокислот. Рекогниция (распознавание) — второй этап синтеза белка, в результате которого аминокислоты избирательно связываются со своими переносчиками — тРНК. Реперфузия — возобновление притока кислорода к тканям. Репликация — удвоение молекулы ДНК в процессе клеточного деления. РесинтезАТФ — образование АТФ в мышечных волокнах непосредственно во время физической работы. Рибосомы — очень мелкие (диаметром до 20 нм), многочисленные (до нескольких десятков тысяч) немембранные органеллы. Основная функция рибосом — синтез белков. Сарколемма — внутренняя оболочка мышечного волокна. Саркомер — структурный элемент миофибриллы. Саркопения — уменьшение массы мышц у людей по мере старения. Саркоплазма — гелеобразный коллоидный раствор, заполняющий объем внутри мышечного волокна. Саркоплазматическая гипертрофия мышечного волокна — увеличение объема мышечного волокна за счет возрастания объема саркоплазмы. Саркоплазматический ретикулум (гладкая эндоплазматическая сеть) — мембранная органелла мышечного волокна, представляющая собой систему трубочек и мешочков (цистерн), окружающих миофибриллы. Сила мышцы (или сила мышечной тяги) — сила, регистрируемая на ее конце (то есть количественная мера взаимодействия мышцы и регистрирующего прибора). Симпласт — крупное образование, состоящее из цитоплазмы и множества ядер. Синхронизация активности ДЕ — одновременное возбуждение ДЕ. Сократимость — способность мышцы укорачиваться при возбуждении, в результате чего возникает сила тяги. Сот — структурный элемент саркомера, представляющий собой систему, основными элементами которой являются один толстый и шесть тонких филаментов, расположенных по обеим сторонам от Z-диска. Срочная адаптация — структурно-функциональная перестройка, происходящая в организме спортсмена непосредственно во время выполнения физических упражнений. Срочный тренировочный эффект выражается через различные показатели, характеризующие срочную адаптацию организма спортсмена к тренировочной нагрузке. Стресс — неспецифическая (общая) реакция организма на воздействие, нарушающее его гомеостаз. Тензиомиография — методика неинвазивной оценки композиции мышечных волокон, разработанная V. Valencia Терминация — процесс завершения синтеза белка. Тканевая жидкость — жидкость, омывающая мышечные волокна и другие компоненты мышцы. Транскрипция — первый этап синтеза белка, в процессе которого происходит синтез молекулы иРНК на участке молекулы ДНК. Трансляция — третий этап синтеза белка, в процессе которого на рибосомах из аминокислот синтезируется молекула белка. Третичная структура белка — пространственная укладка спирали- зованных или линейных участков полипептидной цепи в виде фибриллы (нитевидная) или глобулы (шарообразная). Триплет см. Кодон. Тропонин — один из белков, входящих в состав тонкого филамента. Тропомиозин — один из белков, входящих в состав тонкого филамента. Физиологический поперечник — площадь, соответствующая разрезу, перпендикулярному ходу мышечных волокон. Угол перистости — угол, под которым пучки мышечных волокон располагаются относительно длинника мышцы. Ультразвуковое исследование (УЗИ) — неинвазивное исследование организма человека или животного с помощью ультразвуковых волн. Уступающий режим сокращения мышцы — режим сокращения мышцы, при котором длина мышцы увеличивается. Цитоскелет мышечного волокна — система немембранных органелл, необходимых для создания определенной внутриклеточной структуры. Частота разрядов ДЕ — частота импульсации мотонейрона (МН), иннервирующего мышечные волокна. Четвертичная структура — сложное надмолекулярное образование, состоящее из нескольких белков, имеющих свою собственную первичную, вторичную и третичную структуру. Хромосомы — нуклеопротеидные структуры в ядре клетки, в которых сосредоточена большая часть наследственной информации и которые предназначены для ее хранения, реализации и передачи. Шероховатая эндоплазматическая сеть — мембранная органелла мышечного волокна, окружающая миоядра. Эндомизий — соединительно-тканная оболочка, окружающая мышечные волокна. Эпимизий — слой плотной соединительной ткани, окружающий мышцу. Ядра (миоядра) — образования овальной формы, длиной 10-20 мкм, расположенные под сарколеммой мышечного волокна. Z-диск — диск, разделяющий саркомеры в миофибрилле. [1] Пампинг — метод силовой тренировки, заключающийся в использовании большого количества повторений с небольшими отягощениями (20 % от максимума). [2] Молекула миозина состоит из двух тяжелых цепей и четырех легких цепей. Тяжелая цепь миозина состоит из хвоста и головки. У каждой головки имеются по две легких цепи. [3] Преинкубация — выдерживание в растворе. [4] Если значение рН меньше 7 — среда кислая, если больше 7 — среда щелочная, если значение рН равно 7 — нейтральная. [5] Гомеостаз — относительное, колеблющееся в строго очерченных границах постоянство внутренней среды — крови, лимфы и тканевой жидкости — и устойчивость основных физиологических функций: артериального давления, ЧСС и др. [6] Катаболизм — совокупность химических реакций, за счет которых крупные молекулы превращаются в молекулы меньшего размера. [7] Анаболизм — совокупность химических реакций синтеза. [8] Гипоксия - состояние кислородного голодания тканей. [9] Реперфузия — возобновление притока кислорода к тканям. [10] Да (Дальтон) — внесистемная единица массы, применяемая для масс молекул, атомов, атомных ядер и элементарных частиц. СОДЕРЖАНИЕ 1 ГИПЕРТРОФИЯ СКЕЛЕТНЫХ МЫШЦ ЧЕЛОВЕКА И МЕТОДЫ ЕЕ ОЦЕНКИ 2 1.1 Классификации скелетных мышц. 2 1.2 Особенности функционирования веретенообразных и перистых мышц. 4 1.3 Морфологические показатели, характеризующие степень гипертрофии скелетных мышц, и методы их оценки. 5 1.3.1. Площадь поперечного сечения и объем скелетных мышц. 5 1.3.2. Методы оценки морфологических показателей скелетных мышц. 6 1.3.3. Обхватные размеры тела человека и способы их измерения. 7 1.4. Факторы, влияющие на гипертрофию скелетных мышц. 9 1.4.1. Локализация гипертрофической реакции мышцы.. 9 1.4.2. Расположение мышц. 10 1.4.3. Влияние пола на гипертрофию скелетных мышц. 11 1.4.4. Влияние возраста но гипертрофию скелетных мышц. 12 1.5. Влияние силовой тренировки на морфологические характеристики скелетных мышц. 12 КОНТРОЛЬНЫЕ ВОПРОСЫ... 12 2 ФАКТОРЫ, ВЛИЯЮЩИЕ НА ГИПЕРТРОФИЮ СКЕЛЕТНОЙ МЫШЦЫ... 14 2.1. Скелетная мышца как орган. 14 2.2. Соединение мышечных и сухожильных волокон. 19 2.3. Соединение мышечного волокна и двигательного нерва. 20 2.4. Управление активностью мышцы со стороны цнс. 20 2.5. Биохимия процессов сокращения на уровне мышцы.. 21 2.6. Параметры, определяющие объем скелетных мышц. 21 2.7. Методы оценки параметров, определяющих объем скелетных мышц. 23 2.8. Влияние силовой тренировки на параметры, определяющие объем скелетных мышц. 25 КОНТРОЛЬНЫЕ ВОПРОСЫ... 27 3 ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ГИПЕРТРОФИЮ СКЕЛЕТНОЙ МЫШЦЫ С УЧЕТОМ ТИПОВ МЫШЕЧНЫХ ВОЛОКОН.. 29 3.1. Типы мышечных волокон. 29 3.2. ТИПЫ ДВИГАТЕЛЬНЫХ ЕДИНИЦ.. 30 3.3. Регуляция силы и скорости сокращения мышцы центральной нервной системой. 31 3.4. Параметры, определяющие объем мышцы с учетом типов мышечных волокон. 33 3.5. Факторы, влияющие на площадь поперечного сечения мышечных волокон различных типов. 33 3.6. Влияние силовой тренировки на площадь поперечного сечения мышечных волокон различных типов. 35 3.7. Факторы, определяющие композицию мышечных волокон в скелетных мышцах. 36 3.8. Методы оценки композиции мышечных волокон в скелетных мышцах. 41 3.8.1. Инвазивные методы оценки композиции мышечных волокон в скелетных мышцах 41 3.8.2. Неинвазивные методы оценки композиции мышечных волокон в скелетных мышцах. 42 КОНТРОЛЬНЫЕ ВОПРОСЫ... 48 4 ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ГИПЕРТРОФИЮ СКЕЛЕТНОЙ МЫШЦЫ НА УРОВНЕ МЫШЕЧНОГО ВОЛОКНА.. 49 4.1. Состав мышечного волокна. 49 4.2. Строение мышечного волокна. 54 4.3. Сокращение и расслабление мышечного волокна. 56 4.4. Биохимические процессы, происходящие в мышечном волокне при сокращении и расслаблении мышцы.. 58 4.5. Состав, строение и морфофункционадьная характеристика мышечных волокон различных типов. 61 4.6. Параметры, определяющие объем мышечного волокна. 64 4.7. Влияние тренировки на параметры, определяющие гипертрофию мышечного волокна. 64 4.8. Гистогенез мышечных волокон. 66 4.9. Регенерация мышечных волокон. 67 КОНТРОЛЬНЫЕ ВОПРОСЫ... 70 5 ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ГИПЕРТРОФИЮ СКЕЛЕТНЫХ МЫШЦ НА УРОВНЕ МИОФИБРИЛЛЫ... 71 5.1. Состав и структура миофибриллы.. 71 5.2. Состав и структура саркомера. 71 5.2.1. Состав и структура толстого филамента. 74 5.2.2. Состав и структура тонкого филамента. 74 5.2.3. Состав и структура Z-диска. 75 5.2.4. Состав и структура М-диска. 76 5.3. Модель сокращения мышцы на уровне саркомера. 76 5.4. Параметры, определяющие объем миофибриллы.. 78 5.5. Влияние силовой тренировки на параметры миофибрилл. 79 КОНТРОЛЬНЫЕ ВОПРОСЫ... 80 6 ГИПЕРТРОФИЯ СКЕЛЕТНЫХ МЫШЦ КАК ПРОЯВЛЕНИЕ ДОЛГОВРЕМЕННОЙ АДАПТАЦИИ ЧЕЛОВЕКА К ФИЗИЧЕСКИМ НАГРУЗКАМ.. 81 6.1. Понятие адаптации организма человека к физическим нагрузкам.. 81 6.2. Виды адаптации и тренировочного эффекта. 81 6.3. Условия адаптации. 83 6.4. Виды гипертрофии мышечных волокон. 87 6.5. Гипотезы миофибриллярной гипертрофии мышечных волокон. 88 6.6. Механическое повреждение мышечных волокон как стимул повышенного синтеза белка в мышцах. 91 КОНТРОЛЬНЫЕ ВОПРОСЫ... 95 7 МЕТАБОЛИЗМ БЕЛКОВ В ОРГАНИЗМЕ ЧЕЛОВЕКА.. 97 7.1. Строение и функции нуклеиновых кислот. 97 7.2. Строение молекулы белка. 99 7.3. Переваривание и всасывание белков. 100 7.4. Катаболизм белков в мышечных волокнах. 101 7.5. Синтез белков в мышечных волокнах. 103 7.6. Миофибриллогенез. 105 7.7. Формирование новых мышечных волокон и их гиперплазия. 109 7.8 Концепции, объясняющие повышенный синтез бедка в скелетных мышцах при выполнении силовых тренировок. 111 КОНТРОЛЬНЫЕ ВОПРОСЫ... 115 8 ВЛИЯНИЕ РАЗЛИЧНЫХ ПАРАМЕТРОВ ТРЕНИРОВКИ НА ГИПЕРТРОФИЮ СКЕЛЕТНЫХ МЫШЦ.. 116 8.1. Факторы, сопутствующие миофибриллярной гипертрофии скелетных мышц. 116 8.1.1. Увеличение силы скелетных мышц. 116 8.1.2. Мышечные боли, возникающие ори выполнении силовых упражнений. 118 8.2. Влияние силовой тренировки с отягощениями различной массы на гипертрофию скелетных мышц. 121 8.2.1. Характеристика силовой тренировки с отягощением различной массы.. 121 8.2.2. Гипотеза, объясняющая тренировочные эффекты воздействия на скелетные мышцы отягощений различной массы.. 122 8.3. Влияние силовой тренировки, выполняемой в различных режимах мышечного сокращения, на гипертрофию скелетных мышц. 123 8.3.1. Тренировочные эффекты воздействия на скелетные мышцы работы в различных режимах. 123 8.3.2. Гипотезы, объясняющие тренировочные эффекты воздействияно скелетные мышцы роботы в различных режимах. 125 8.4. Влияние тренировки методом «до отказа» на гипертрофию скелетных мышц. 128 8.4.1. Характеристика метода «до отказа». 128 8.4.2. Тренировочные эффекты воздействия на скелетные мышцы тренировки методом «до отказа». 130 8.4.3. Гипотезы, объясняющие тренировочные эффекты воздействия но скелетные мышцы роботы методом «до отказа». 131 8.5. Влияние прекращения тренировки и последующего ее возобновления на гипертрофию скелетных мышц. 135 8.6. Влияние порядка упражнений, используемых в тренировке, на силу и гипертрофию скелетных мышц. 136 8.7. Влияние различных программ тренировки на силу и гипертрофию скелетных мышц. 137 КОНТРОЛЬНЫЕ ВОПРОСЫ... 138 ГЛОССАРИЙ... 140
ГИПЕРТРОФИЯ СКЕЛЕТНЫХ МЫШЦ ЧЕЛОВЕКА И МЕТОДЫ ЕЕ ОЦЕНКИ |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Последнее изменение этой страницы: 2019-04-09; Просмотров: 479; Нарушение авторского права страницы
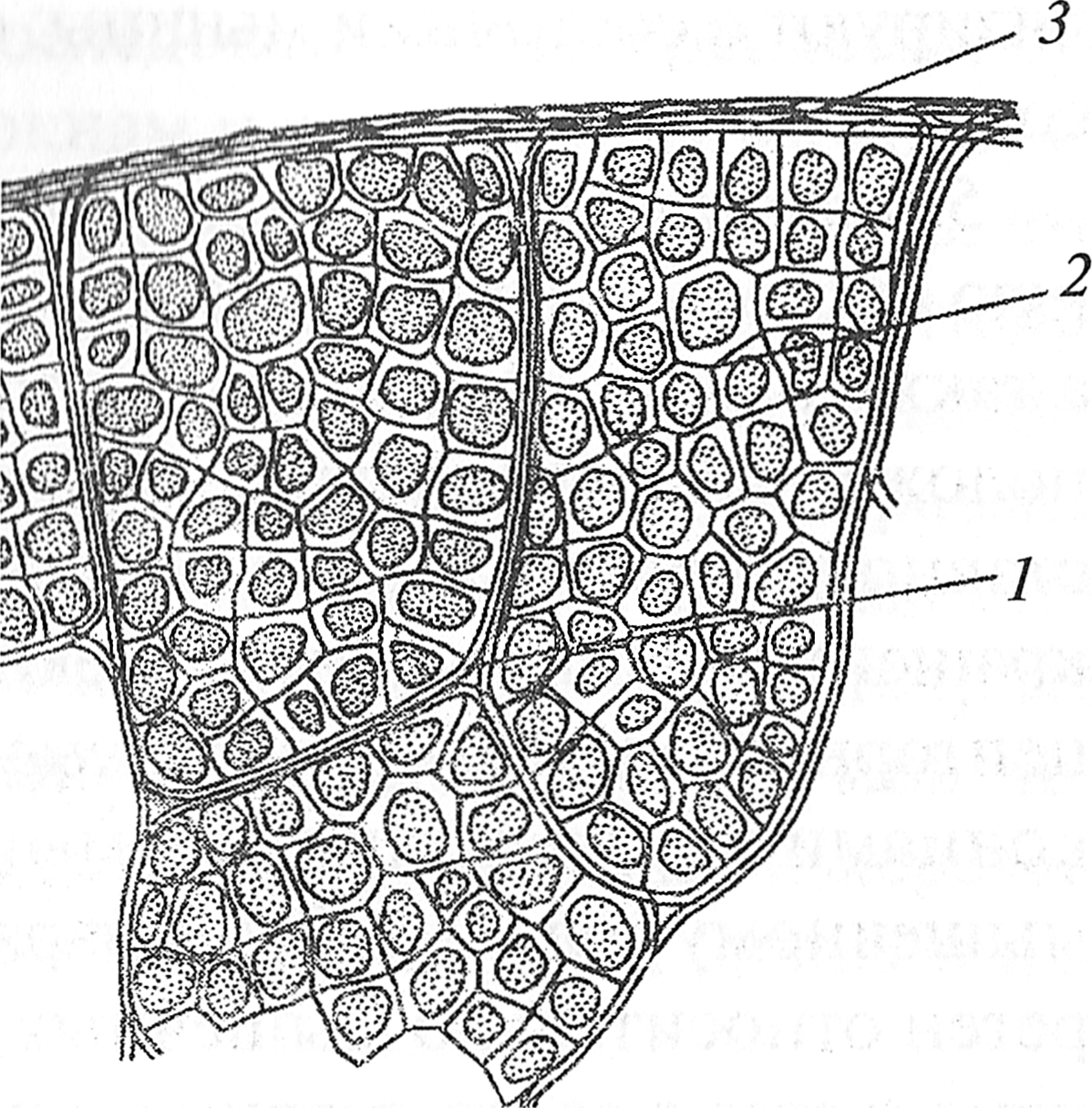
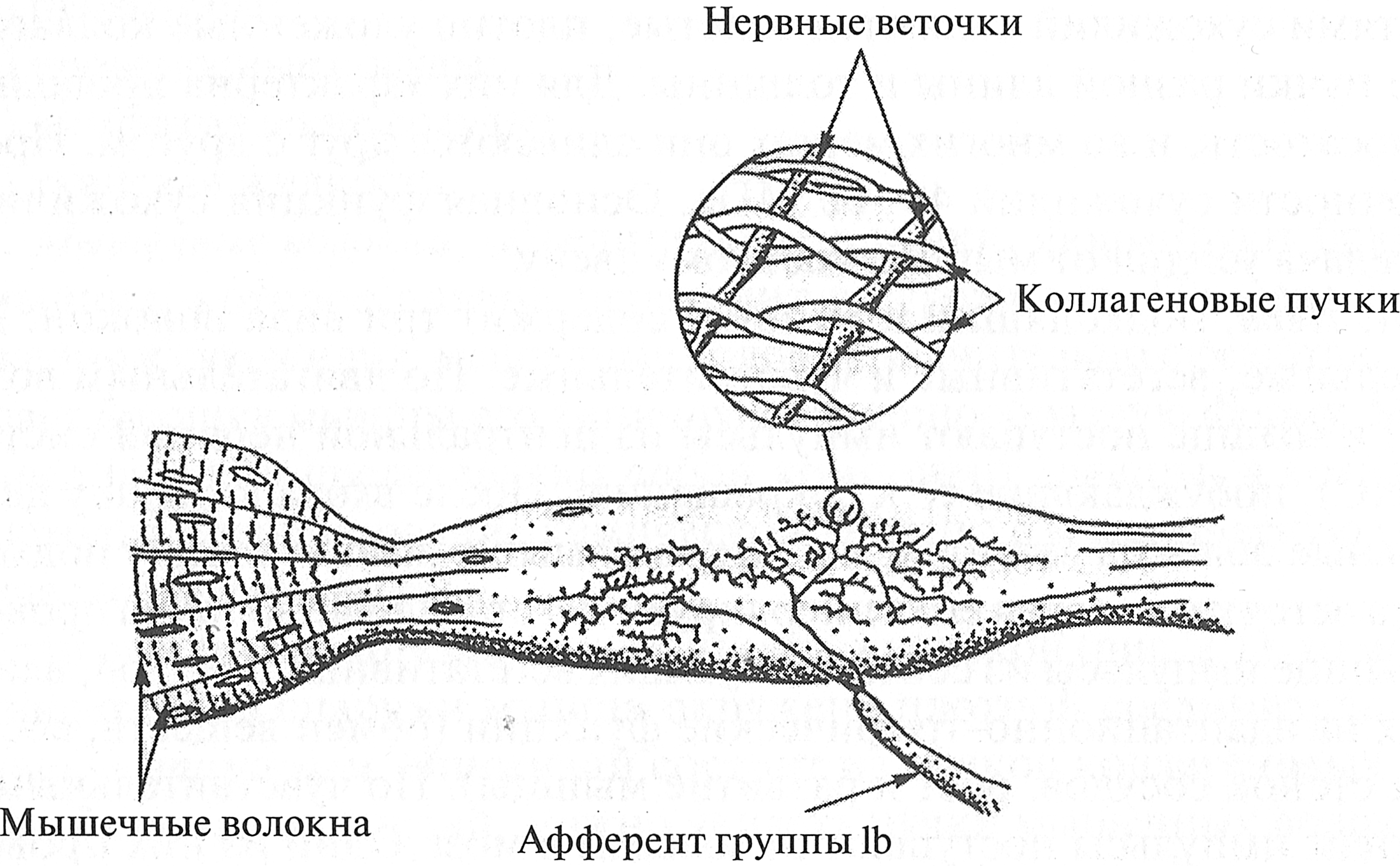


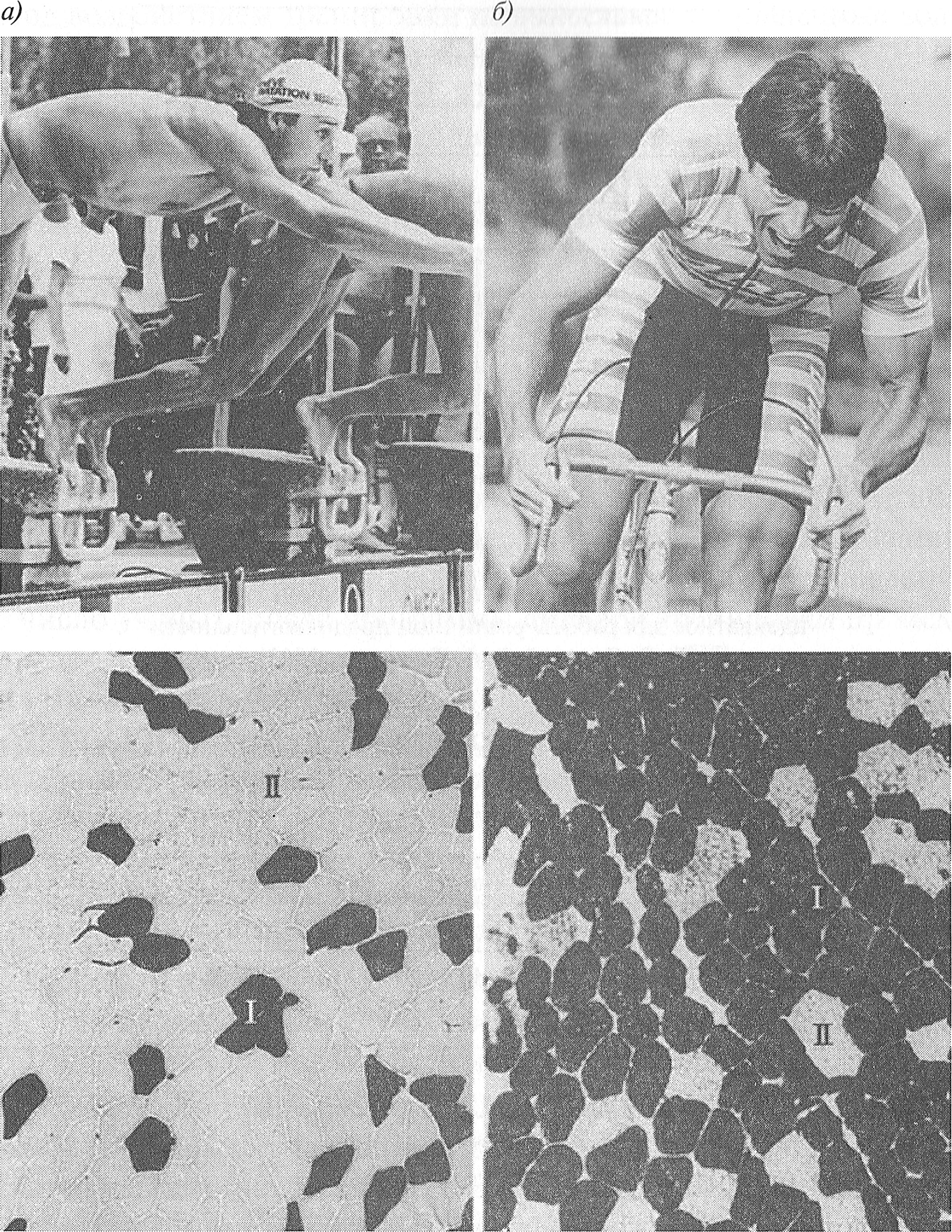
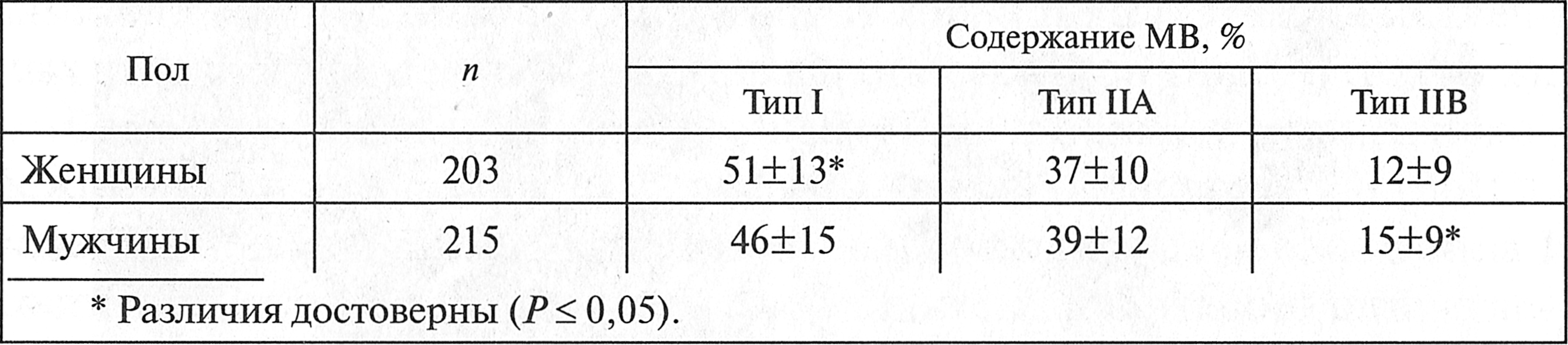

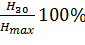
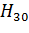 — среднее арифметическое значение высоты тридцать первого, тридцать второго и тридцать третьего прыжков;
— среднее арифметическое значение высоты тридцать первого, тридцать второго и тридцать третьего прыжков; 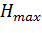 — среднее арифметическое высоты трех первых прыжков.
— среднее арифметическое высоты трех первых прыжков.