|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
К настоящему времени идентифицировано несколько механизмов возникновения болезни геномного импринтинга.
1. Однородительская дисомия (ОРД) — наличие у больного двух хромосом с импринтинговыми участками, полученными от одного из родителей. В этом случае человек имеет нормальный кариотип (46 хромосом), однако, обе хромосомы одной из пар получены им от одного из родителей. Причин возникновения ОРД несколько, но наиболее вероятны из них четыре: а) нерасхождение хромосом во втором мейотическом делении с образованием совершенно идентичных хроматид; б) нерасхождение хромосом в первом делении мейоза с возникновением негомологичных хромосом; в) исправление возникшей при делении клеток трисомии или моносомии по хромосомам, содержащим импринтированные гены; г) соматическая рекомбинация хромосом, т.е. обмен между несестринскими хроматидами гомологичных хромосом в соматических клетках. 2. Хромосомные перестройки в импринтинговых участках, содержащих экспрессирующиеся гены. Наиболее распространены микроделеции, выявляемые специальными цитогенетическими методами (прежде всего FISH-анализом), а также транслокации и инверсии определенного хромосомного региона. 3. Делеции в области импринтинговых центров, контролирующих процессы метилирования хромосом. Точные механизмы развития геномного импринтинга пока не установлены, но предполагается участие в этом процессе специфического метилирования цитозиновых оснований ДНК, образующихся антисмысловых транскриптов РНК ( антисмысловой траскрипт РНК - получаемый искусственно или природный полирибонуклеотид, комплементарный определенной мРНК и подавляющий ее биологическую активность за счет образования с ней дуплекса, что препятствует трансляции мРНК на рибосомах). Импринтинг генов осуществляется с помощью процесса метилирования ДНК. Синдром Прадера-Вилли - редкое наследственное заболевание, причиной которого Является отсутствие отцовской копии участка хромосомы 15q11-13 ( в участке хромосомы 15).
Делеция этого же района материнской 15-й хромосомы обусловливает развитие синдрома Ангельмана (комплекс врожденных психических расстройств). Синдром Ангельмана легко отличим от синдрома Прадера-Вилли.
Образование однояйцевых и разнояйцевых близнецов. Сиамские близнецы Близнецов человека разделяют на две основные группы: монозиготные (однояйцовые, идентичные) и дизиготные (двуяйцовые, неидентичные) близнецы. Дизиготные близнецы развиваются в результате двух независимых процессов оплодотворения, тогда как монозиготные происходят из одного зародыша, клетки которого каким-то образом разделились. Последнее означает, что изолированный бластомер млекопитающего способен образовать целый зародыш. Такое заключение было подтверждено опытом разрушения одного бластомера у 2-клеточного зародыша кролика (Seidel. 1952). Оставшийся бластомер был способен к формированию полноценного взрослого организма. Даже один бластомер 8-клеточного зародыша мыши может успешно развиться в полноценную взрослую особь (Kelly, 1977). Было также показано, что при инъекции клеток внутренней клеточной массы (но не трофобласта) в бластоцисту они способны принимать участие в образовании нового зародыша (Gardiner. Rossant. 1976). Таким образом, идентичные близнецы возникают путем разделения бластомеров на ранних стадиях или даже разделения ВКМ на две части в пределах одной бластоцисты. По-видимому, именно так оно и происходит при развитии примерно 0, 25% детей. Около 33% идентичных близнецов имеют два полноценных независимых хориона, что указывает на разделение зародыша еще до образования ткани трофобласта на 5-е сутки беременности. У оставшихся 67% идентичных близнецов хорион общий; в этих случаях разделение ВКМ произошло, очевидно, после образования трофобласта. К 9 суткам зародыш человека завершает формирование другого внезародышевого образования - амниона. Из этой ткани образуется амниотический мешок (или водная оболочка), заполненный амниотической жидкостью, который окружает зародыш и защищает его от обезвоживания и последствий резких движений. Если бы разделение зародыша произошло между образованием хориона на пятые сутки и амниона на девятые. то зародыши должны были бы иметь один хорион и два амниона. Так обстоит дело приблизительно у двух третей идентичных близнецов. Небольшой процент идентичных близнецов имеет общий хорион и общий амнион. Это означает, что разделение зародыша произошло после 9 сут беременности. Такие новорожденные рискуют оказаться сросшимися между собой («сиамские близнецы»).
Способность образовать целый зародыш из клеток, которые в норме участвовали бы в образовании только его части, называется регуляцией. Регуляцией является также способность двух и большего числа ранних зародышей образовать одну аллофенную мышь, а не двойню, тройню или многоголового урода. Имеются данные в пользу того, что аллофенная регуляция может происходить и у человека (Chappelle el al., 1974; Mayr et al., 1979). Описаны индивидуумы, в организме которых имеются клетки двух разных генетических типов (XX и XY), каждый из которых обладает собственным набором генетических характеристик. Самое простое объяснение существования такого явления заключается в том, что эти индивидуумы возникли в результате слияния двух зародышей мужского и женского пола, которые развивались одновременно. Если такое объяснение правильно, то в этих случаях два близнеца, брат и сестра, слились для образования одной химерной особи.
Рисунок, иллюстрирующий зависимость между временем разделения зародыша человека, приводящего к образованию монозиготных (идентичных, однояйцовых) близнецов, и их взаимоотношением с зародышевыми оболочками. А. Разделение происходит до образования трофэктодермы, при этом каждый из близнецов имеет собственные хорион и амнион. Б. Разделение происходит после образования трофэктодермы, но до образования амниона: в результате каждый из близнецов имеет собственный амниотический мешок, но хорион у них общий. В. При разделении после образования амниона близнецы находятся в общем амниотическом мешке и имеют общий хорион.
15. Имплантация у человека После оплодотворения зигота постепенно спускается по маточной трубе в полость матки. В этот период, в течение примерно трех дней, зигота проходит стадию клеточного деления, известную как дробление. При дроблении число клеток увеличивается, но общий их объем не меняется, так как каждая дочерняя клетка мельче, чем исходная. Первое дробление происходит примерно через 30 часов после оплодотворения и дает две совершенно одинаковые дочерние клетки. Второе дробление наступает через 10 часов после первого и приводит к образованию четырехклеточной стадии. Примерно через 50-60 часов после оплодотворения достигается стадия т.н. морулы - шара из 16 и более клеток. По мере продолжения дробления наружные клетки морулы делятся быстрее, чем внутренние, в результате наружный клеточный слой (трофобласт) отделяется от внутреннего скопления клеток (т.н. внутренней клеточной массы), сохраняя с ними связь только в одном месте. Между слоями образуется полость, бластоцель, которая постепенно заполняется жидкостью. На этой стадии, наступающей через три-четыре дня после оплодотворения, дробление заканчивается и эмбрион называют бластоцистой, или бластулой. В течение первых дней развития, эмбрион получает питание и кислород из секрета (выделений) маточной трубы. Примерно через пять-шесть дней после оплодотворения, когда бластула находится уже в матке, трофобласт образует пальцевидные ворсинки, которые, энергично двигаясь, начинают внедряться в ткань матки. В то же время, по-видимому, бластула стимулирует выработку ферментов, способствующих частичному перевариванию слизистой (эндометрия) матки. Примерно на 9-10 день эмбрион имплантируется (врастает) в стенку матки и оказывается полностью окруженным ее клетками; с имплантацией эмбриона прекращается менструальный цикл. В дополнение к своей роли в имплантации, трофобласт участвует также в образовании хориона - первичной мембраны, окружающей эмбрион. В свою очередь хорион содействует образованию плаценты, губчатой по структуре мембраны, через которую эмбрион в дальнейшем получает питание и выводит продукты обмена.
Имплантация - процесс врастания зародыша в слизистую оболочку матки, который осуществляется на 7-8-е сутки развития. К этому времени зародыш под влиянием перистальтических сокращений, движения ресничек и перемещения секрета эпителиоцитов маточной трубы проходит от ампульной части последней в полость матки. Это, так называемый, трубный период жизни зародыша (первая-четвертая сутки). В то время, когда зародыш перемещается по яйцеводу по направлению к матке, бластоциста увеличивается в объеме. В плазматических мембранах клеток трофэктодермы имеется натриевый насос (Na+/К+-АТФаза), который переносит ионы натрия в центральную полость. Такое накопление ионов натрия вызывает поступление в полость воды осмотическим путем, в результате чего размеры полости бластоцисты увеличиваются. В период перемещения зародыша большое значение имеет тот факт, что прозрачная оболочка предотвращает прилипание бластоцисты к стенкам яйцевода. Когда у человека такое прилипание происходит, наблюдается «трубная беременность». Это опасное явление, поскольку имплантация зародыша в яйцевод может вызвать угрожающее жизни кровотечение. Однако, когда зародыш достигает матки, он должен освободиться от прозрачной оболочки для того, чтобы прикрепиться к стенке матки. *Бластоциста мыши освобождается от оболочки путем лизиса в ней небольшого отверстия, через которое она выдавливается при увеличении cвoего объема. Гистологические данные свидетельствуют о том, что одна из клеток трофобластической стенки (в части, не прилежащей к внутренней клеточной массе) образует вырост, который приходит в контакт с прозрачной оболочкой. На плазматической мембране этого выроста локализована трипсиноподобная протеаза, стрипсин, которая лизирует отверстие в фибриллярном матриксе оболочки. Освободившись от оболочки, бластоциста может вступить в непосредственный контакт с маткой. Здесь трофобласт будет секретировать другую трипсиноподобную протеазу, активатор плазминогена. Полагают, что этот расщепляющий белок фермент необходим для разрушения ткани матки, чтобы бластоциста могла погрузиться в ее стенку. С пятой до семи суток в полости матки находится зародыш в виде свободной бластоцисты (не связанной с определенным участком эндометрия). Питание зародыша в названные периоды осуществляется частично за счет питательных веществ овоцита, частично - за счет секрета маточных труб и желез эндометрия. Седьмую-восьмые сутки, когда бластоциста вступает в контакт со слизистой оболочкой матки, считают критическим периодом развития, поскольку существует угроза, что в результате тех или иных причин врастания в эндометрий не осуществится и зародыш погибнет. Различают две фазы имплантации - адгезию и инвазию. Различают две стадии имплантации: адгезию (прилипание), когда зародыш прикрепляется к внутренней поверхности матки, и инвазию (погружение) - внедрение зародыша в ткани слизистой оболочки матки. На 7-е сут в трофобласте и эмбриобласте происходят изменения, связанные с подготовкой к имплантации. Бластоциста сохраняет оболочку оплодотворения. В трофобласте увеличивается количество лизосом с ферментами, обеспечивающими разрушение (лизис) тканей стенки матки и тем самым способствующими внедрению зародыша в толщу ее слизистой оболочки. Появляющиеся в трофобласте микроворсинки постепенно разрушают оболочку оплодотворения. Зародышевый узелок уплощается и превращается в зародышевый щиток, в котором начинается подготовка к первой стадии гаструляции. Имплантация продолжается около 40 ч. Одновременно с имплантацией начинается гаструляция (образование зародышевых листков). Это первый критический период развития. В первой стадии трофобласт прикрепляется к эпителию слизистой оболочки матки, и в нем формируются два слоя - цитотрофобласт и симпластотрофобласт (в конспекте синцититрофобласт - это одно и тоже? Я хз ) Во второй стадии симпластотрофобласт, продуцируя протеолитические ферменты, разрушает слизистую оболочку матки. Формирующиеся при этом ворсинки трофобласта, внедряясь в матку, последовательно разрушают ее эпителий, затем подлежащую соединительную ткань и стенки сосудов, и трофобласт вступает в непосредственный контакт с кровью материнских сосудов. Образуется имплантационная ямка, в которой вокруг зародыша появляются участки кровоизлияний. Питание зародыша осуществляется непосредственно из материнской крови (гематотрофный тип питания). Из крови матери зародыш получает не только все питательные вещества, но и кислород, необходимый для дыхания. Одновременно в слизистой оболочке матки из клеток соединительной ткани, богатых гликогеном, происходит образование децидуальных клеток. После полного погружения зародыша в имплантационную ямку отверстие, образовавшееся в слизистой оболочке матки, заполняется кровью и продуктами разрушения тканей слизистой оболочки матки. В последующем дефект слизистой оболочки исчезает, эпителий восстанавливается путем клеточной регенерации. Гематотрофный тип питания, сменяющий гистиотрофный, сопровождается переходом к качественно новому этапу эмбриогенеза - второй фазе гаструляции и закладке внезародышевых органов.
Давал термин:
Окно имплантации – период второй (лютеиновой) фазы менструального цикла, (6-7 суток от зачатия) когда в эндометрии формируются наиболее благоприятные условия для прикрепления эмбриона. Сочетание множества этих условий – залог успешной имплантации и течения беременности без акушерских осложнений. Из моего коснпекта: Материнский организм осуществляет имуннодепрессивную реакцию. Иммунодепрессоры снижают реакцию на имплантацию. 16.
Определение пола у человека — процесс развития половых различий у людей. Этот процесс определяется как развитие фенотипических структур в результате воздействия гормонов, которые вырабатываются в зависимости от развития гонад. Развитие половых различий, или половая дифференциация, включает в себя развитие гениталий и внутренних половых путей, молочных желёз, волос на теле и играет роль при гендерной идентификации[2]. Формирование половых различий начинается с формирования гоносом (половых хромосом). За формирование фенотипических различий между мужским и женским организмами из недифференцированной зиготы отвечают сложные механизмы. Женским считается организм, имеющий две X-хромосомы, мужским — организм, имеющий одную Y-хромосому и одну X-хромосому. На ранних этапах эмбрионального развития все человеческие организмы имеют одинаковую внутреннюю структуру. Она включает в себя мезонефральные и парамезонефрические протоки. Присутствие гена SRY на Y-хромосоме вызывает развитие яичек в мужском организме и последующее производство гормонов, под воздействием которых исчезают парамезонефрические протоки. В женском организме исчезают мезонефральные протоки. У человека есть первичные и вторичные половые признаки Первичные: анатомические и физиологические особенности, связанные с формированием половых клеток и их встреча в процессе оплодотворения (половые железы, наружные половые органы) Вторичные: характер жировых отложений, строение таза, тип распределения волос по телу, высота голоса. Гонады способны развиваться в двух направлениях: либо в яичнике, либо в семеннике. До конца 8-й недели гонада не проявляет свой пол. У млекопитающих, в том числе и человека, развитие мужского организма невозможно без наличия Y-хромосомы, то есть прежде всего половая принадлежность у человека определяется сочетанием половых хромосом в его кариотипе. Однако установлено, что для развития организма мужского пола недостаточно только наличия гена SRY на Y-хромосоме, который определяет дифференцировку мужских половых желёз и синтез ими тестостерона. Ткани-мишени, на которые действует тестостерон, должны быть чувствительны к нему. Для этого необходим белок-рецептор, являющийся продуктом особого гена, локализованного в Х-хромосоме.. Он обеспечивает проникновение гормона в клетки нужных тканей. Если в этом гене происходит мутация, нарушающая образование нормального белка-рецептора, то ткани-мишени становятся нечувствительны к тестостерону. Не использовав возможность формирования мужского фенотипа на определённом этапе онтогенеза, организм осуществляет развитие по женскому типу. В итоге формируется особь с кариотипом XY, но внешне схожая с женщиной. Такие субъекты бесплодны, поскольку их семенники недоразвиты, а выводные протоки нередко формируются по женскому типу (недоразвитая матка, влагалище). Вторичные половые признаки характерны для женского пола. Такое нарушение у человека известно как синдром Морриса. Таким образом, пол у человека формируется как результат комплементарного взаимодействия неаллельных генов. Зародыши млекопитающих развиваются по пути самки, если организм не содержит яичек, в противоположном случае, зародыш развивается по пути самца. В случае, если половые железы удалены до формирования яичек или яичников, организм также развивается в самку, независимо от того, какие хромосомы содержит геном. Для развития женских половых органов требуются и другие факторы, например, яичники млекопитающих не развиваются без сигнального белка Wnt4. Введение гена SRY в геном ХХ-зиготы мыши приводит к развитию зародыша по пути самца, хотя остальные гены Y-хромосомы у такого эмбриона отсутствуют. Подобные мыши с «обращенным полом» не способны к сперматогенезу, так как их геном не содержит других генов Y-хромосомы, которые необходимы для образования спермы. Ген SRY экспрессируется в соматических клетках развивающихся половых желез и вызывает их дифференцировку этих клеток в клетки Сертоли. Последние направляют развитие организма по мужскому пути, в том числе, стимулируют превращение клеток яичка в клетки Лейдига, секретирующие тестостерон в кровь. Белок SRY связывает соответствующие участки ДНК и запускает транскрипцию других генов, принимающих участие в формировании клеток Сертоли. Одним из важных генов, активируемых SRY, является ген SOX9, который экспрессируется у всех самцов позвоночных. В отсутствие генов SRY или SOX9, у XY-зародышей млекопитающих вместо яичек формируются яичники, а вместо клеток Сертоли формируются фолликулярные клетки. Остальные клетки половых желез вместо клеток Лейдига превращаются в клетки теки, которые, в пубертатный период начинают секретировать эстроген. · Ген SRY (англ. Sex-determining Region Y) располагается на Y-хромосоме большинства млекопитающих, и принимает участие в развитии организма по мужскому типу. · SOX9 - расположен на 17 хромосоме, экспрессируется на стадии эмбриональной закладки желез. · WNT-4 - сигнал на образование яичника, репрессирует развитие мужского пола. · WT1- ген WT1 человека является геном-супрессором опухолей, мутационные нарушения которого наблюдают при злокачественных новообразованиях Вилмса, возникающих в почках в детском возрасте. Четыре белка, кодируемые этим геном, возникают в результате альтернативного сплайсинга, и, по крайней мере, один из них является фактором транскрипции (был дан в конспекте, не знаю зачем) *Когда Джост (Jost, 1953) удалял еще недифференцированные гонады у плода кролика, то независимо от генотипа (XX или XY) развившаяся особь всегда оказывалась женской. Все подопытные кролики имели яйцеводы, матки, влагалища и были лишены мужских половых органов. В отсутствие гонад развитие идет по женскому пути. Следовательно, определение пола у млекопитающих зависит от какого-то продукта Y-хромосомы или ею регулируется. Если Y-хромосома отсутствует, то зачатки гонад развиваются в яичники. Эстрогены, продуцируемые яичником, обусловливают развитие мюллерова протока во влагалище, шейку матки, матку и яйцеводы. При наличии Y-хромосомы последняя продуцирует или регулирует формирование фактора (до сих пор еще не охарактеризованного), который детерминирует развитие семенников, т.е. вынуждает развивающуюся гонаду вступать на путь, ведущий к развитию семенника, а не яичника. Коль скоро семенник сформировался, он начинает секретировать два важных гормона. Первый из них – гормон, направленный против мюллерова канала, или антимюллеровский гормон (АМГ) разрушает ткань, способную в альтернативном случае дать начало матке, яйцеводу, шейке матки и верхней части влагалища. Другой гормон – тестостерон – приводит к маскулинизации плода, вызывая формироване пениса, мошонки и других частей мужской анатомии, одновременно стимулируя редукцию зачатков молочных желез. Итак, фенотип организма останется женским, если его не изменят два гормона, вырабатываемые семенниками плода. Перейдем к более подробному анализу этих событий.
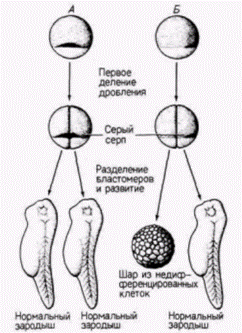
Вопрос 17 Эмбриональная индукция (прогрессивная детерминация) Шпеман провел опыт и доказал, что ядра ранних бластомеров тритона идентичны, т. е. каждое из них способно обеспечить развитие целой личинки. Пользуясь волоском ребенка, он перевязывал им яйцо тритона вскоре после оплодотворения в плоскости первого деления дробления. Затем он несколько стягивал петлю так, что все деления ядер происходили лишь в одной из половин. Наконец на стадии 16 бластомеров одно ядро смогло проскользнуть через перетяжку в безъядерную половину. Дробление начиналось и в этой половине, а петлю, накинутую на яйцо. Шпеман стягивал все сильнее, пока не разделял яйцо на две изолированные половины. В результате развивались два зародыша-близнеца, причем один был немного старше другого. Результаты этого опыта позволили Шпеману сделать вывод, что ядра ранних зародышей амфибий идентичны и каждое способно обеспечить развитие целого организма.
Но когда Шпеман проделал сходный опыт с перетягиванием яйца перпендикулярно к плоскости первого деления дробления (т.е. разделял яйцо не на левую и правую половины, а на будущую спинную и брюшную стороны), он получил совершенно другой результат! По обе стороны от перетяжки ядра продолжали делиться, но лишь из клеток одной стороны образовалась нормальная личинка. Из другой половины возникала только неорганизованная масса ткани, названная Шпеманом «кусок живота». Эта масса тканей представляла собой шарик эпидермальных клеток (эктодерма), содержащий внутри кровь и мезенхиму (мезодерма) и клетки кишки (энтодерма), но в нем не было дорсальных структур, таких, как нервная система, хорда или сомиты. Почему описанные выше два опыта дали разные результаты? В яйцах амфибий после оплодотворения происходит резкое смещение кортикального слоя цитоплазмы и у некоторых видов амфибий такое движение приводит к образованию серого серпа в области, прямо противоположной месту проникновения спермия в яйцо. Кроме того, плоскость первого деления дробления обычно делит эту область поровну между двумя бластомерами, из которых, если их отделить друг от друга, развиваются две нормальные личинки. Однако, если плоскость первого деления отклоняется от середины серого серпа (в редких случаях спонтанно или в опыте, в котором исследователь перетягивает яйцо волосяной петлей перпендикулярно плоскости нормального деления), то материал серого серпа может попасть только в один из двух бластомеров. Шпеманн обнаружил, что когда эти два бластомера разделены полностью, нормально развивается только тот бластомер, который содержит материал серого серпа. В дальнейшем из серпа образуются клетки, которые инициируют гаструляцию и формируют спинную губу бластопора. Клетки спинной губы бластопора запрограммированы, чтобы инвагинировать внутрь зародыша и, таким образом, начинать гаструляцию и формирование архентерона. Поскольку все будущее развитие амфибий зависит от взаимодействия клеток, перемещающихся в процессе гаструляции, Шпеманн предположил, что материал серого серпа играет решающую роль в инициации гаструляции и что в период гаструляции в развитии зародыша происходят кардинальные изменения. Позднее Шпеманн доказал, что в период гаструляции действительно резко меняется потенция клеток. Он обнаружил, что клетки ранней гаструлы еще не детерминированы к их конечной дифференцировке, тогда как на стадии поздней гаструлы судьба клеток уже определена (смотрим вопрос 20, часть про тритонов). Детерминация (лат. determinatio - ограничение, определение) - предопределение судьбы клеток, чтобы в конце из них развился совершенно определенный клеточный тип. Клеточный материал считают детерминированным, начиная со стадии, когда он впервые при пересадке в чуждое место дифференцируется в тот тип клеток, который из него образуется при нормальном развитии. Вопрос 18 Ближние тканевые взаимодействия Есть два основных типа ближних тканевых взаимодействий. К первому типу относятся разрешающие (пермессивные) взаимодействия. При этом отвечающая ткань потенциально готова к экспрессии и нуждается только в определенных условиях, которые разрешили бы экспрессию ее признаков. Другой тип ближних тканевых взаимодействий - инструктивные взаимодействия. При такого рода взаимодействиях изменяется тип отвечающей ткани, т.к. происходит включение новых генетических программ, определяющих спецификацию клеток. Можно вспомнить образование хрусталика. Ближние клеточные взаимодействия лежат в основе механизма, обеспечивающего координированное развитие органов, поскольку отвечающая ткань также может стать индуцирующей тканью. В каскаде индукций, приводящих к образованию глаза, хорда индуцирует возникновение нервной трубки: выпячивание нервной трубки – глазной пузырь – в результате контакта с эктодермой инструктирует ее клетки к превращению в хрусталик. В свою очередь хрусталик инструктирует образование роговицы в эпидермисе, который располагается непосредственно над ним. Индукция (лат. inductus – пробуждение) – межклеточное взаимодействие индуктора и реагирующей системы в результате которого изменяется направление развития последней. Индуктором могут быть ткань, зачаток клетка, которые вырабатывают сигнальные молекулы, действие которых вызывает изменение потенции развития ткани-мишени. Реагирующая ткань (мишень) – это особым способом дифференцированная ткань, которая характеризуется компетенцией. Компетенция (лат. competo – быть способным) – способность ткани отвечать на индуцирующий стимул.
Вопрос 19 Нейруляция и образование осевого комплекса зачатков
Первым указанием на то, что данный участок эктодермы предназначен стать нервной тканью, является изменение формы клеток. Удлинение дорсальных эктодермальных клеток приводит к тому, что эти проспективные нейральные области приподнимаются над окружающей их эктодермой, образуя нервную пластинку. Вскоре после образования нервной пластинки ее края утолщаются и движутся вверх, образуя нервные валики, а в центре нервной пластинки возникает U-образный нервный желобок, разделяющий зародыша на будущие правую и левую половины. Нервные валики, приподнимаясь, движутся навстречу друг другу и, в конечном счете, сливаются по среднеспинной линии зародыша. В результате нервная пластинка превращается в лежащую под эктодермальным эпителием нервную трубку. Клетки в области, соединяющей этот эпителий с нервной трубкой, являются клетками нервного гребня. Позднее эти клетки мигрируют по всему зародышу, давая начало нескольким клеточным популяциям, в том числе пигментным клеткам и клеткам периферической нервной системы. Формирование нервной трубки происходит неодновременно во всей эктодерме. У раннего зародыша млекопитающих нервная трубка на всем ее протяжении прямая. Но нейруляция в головной области идет быстрее, тогда как в хвостовом отделе еще продолжается гаструляция. Регионализация нервной трубки является также результатом изменений ее формы. В головном конце (где будет формироваться головной мозг) стенка трубки широкая и толстая. В ней рядом вздутий и сужений обозначены различные отделы головного мозга. Сначала появляются три первичных мозговых пузыря: первичный передний мозг, средний мозг и первичный задний мозг. Первичный передний мозг затем подразделяется на передний конечный мозг и промежуточный мозг. Получается 5 мозговых пузырей. Каудальнее головного отдела она сохраняет вид простой, сужающейся по направлению к хвосту трубки. Два открытых конца нервной трубки называются соответственно передним и задним нейропором.
Вопрос 20 Первичная эмбриональная индукция и ее региональная специфичность Индукция – процесс, в течении которого одна область зародыша заставляет другую дифференцироваться в определенном направлении (определение с лекции). Первичная эмбриональная индукция – ключевое взаимодействие, в котором дорсальная мезодерма индуцирует эктодерму к дифференцировке в нейральные структуры. Открытие организатора Показав, что в период органогенеза предопределение судьбы зачатков происходит в результате взаимодействия (вспоминаем развитие хрусталика), Шпеманн попытался распространить свою методологию анализа эмбрионального развития на более ранние стадии. Он трансплантировал различные зачатки гаструлы тритонов 2 разных видов (не будем забивать голову их названиями) на стадиях ранней и поздней гаструлы. Яйца этих видов отличаются своей пигментацией: у одного яйца светлые, у другого – темные. Применяя реципрокные трансплантации (туда сюда, кусочки поменял местами), Шпеманн мог прослеживать судьбу пересаженных естественно маркированных кусочков. Шпеманн установил, что трансплантированные зачатки ранней гаструлы развивались в соответствии с новыми условиями окружающей их среды: презумптивная нейроэктодерма развивалась на брюшной стороне зародыша как эпидермис, тогда как презумптивный эпидермис в области нейральной пластинки развивался как нервная ткань. Из этих наблюдений следовал вывод, что на стадии ранней гаструлы исследованные зачатки не были детерминированы и развивались зависимо, принимая судьбу окружающих их тканей. Иная картина наблюдалась спустя несколько часов, когда трансплантацию проводили на стадии поздней гаструлы. Теперь при трансплантации материала нейроэктодермы на брюшную сторону формировались структуры, характерные для нервной системы. Презумптивная эпидермальная ткань, пересаженная в область нейроэктодермы, также сохраняла свои свойства в новом окружении. Из этих экспериментов следовало, что на стадии гаструляции происходит детерминация свойств эпидермальной и нервной ткани. самый сок вопроса Особое значение имели результаты, полученные Шпеманном и его ученицей Хильдой Мангольд в опытах по трансплантации зачатка спинной губы бластопора, взятого на стадии ранней гаструлы. Пересадка спинной губы бластопора на вентральную сторону зародыша вызывала полную реорганизацию морфогенетических процессов, в результате которой в этой области происходило развитие дополнительной хорды, нервной трубки, сомитов. Дорсальная губа бластопора предстала, таким образом, организатором развития осевых структур и при трансплантации вызывала формирование вторичного зародыша (рисунок на стр 65, том 2). Из экспериментов Шпеманна и его учеников следовало, что при нормальном развитии материал спинной губы бластопора образует хордомезодерму, которая контактирует с эктодермой и индуцирует в последней процессы, ведущие к возникновению нейральной ткани и нервной системы. Процесс образования нейральной ткани из компетентной эктодермы под действием организатора был назван Шпеманном первичной эмбриональной индукцией. Дальнейшее развитие зародыша, по Шпеманну, представлялось цепью индукционных событий, в результате которых происходила последовательная дифференциация зародыша. Вскоре во многих лабораториях мира было показано, что гомологичные спинной губе бластопора амфибий зачатки зародышей рыб, рептилий, птиц и млекопитающих обладают такими же свойствами организатора и способны индуцировать развитие осевых структур. Региональная специфичность Внешне однородная мезенхима, взятая из разных областей тела зародыша, обладает способностью при взаимодействии с одним и тем же видом эпителиальной ткани индуцировать разные органы. Кожа цыпленка дает начало трем типам покровных структур, почти полностью образованных эктодермальными клетками. Это широкие перья крыла, узкие перья голени и чешуи и когти ног. Отделив эмбриональные эпителий и мезенхиму друг от друга, можно затем совмещать их друг с другом различными способами. За специфичность индукции отвечает мезенхима. Тот же самый тип компетентной эктодермы развивается в соответствии с областью происхождения мезодермы. Таким образом, мезенхима играет инструктивную роль, включая в игру разные наборы генов отвечающих клеток.
ДЛИННЫЙ ВАРИАНТ я сначала его сделала, жалко удалять Одним из наиболее любопытных явлений в первичной эмбриональной индукции является региональная специфичность образующихся нейральных структур. Переднемозговая, заднемозговая и спинокаудальная области нервной трубки должны быть организованы в соответствующем порядке спереди назад. Следовательно, хордомезодермальная ткань крыши первичной кишки индуцирует не только нервную трубку в целом, но также и ее специфические отделы. Эта региональная специфичность индукции была обнаружена Отто Мангольдом в серии экспериментов, в которых участки крыши архентерона, взятые на стадии ранней нейрулы тритона, имплантировали в бластоцель ранней гаструлы тритона. У зародышей, только что завершивших гаструляцию, после удаления нервной пластинки из лежащей под ней крыши первичной кишки были вырезаны четыре последовательных участка (то есть кусочки мезодермы). Эти участки по отдельности имплантировали в бластоцель зародышам на стадии ранней гаструлы. Самая передняя часть крыши первичной кишки индуцировала образование балансёров и частей ротового аппарата. Следующая за ней передняя часть индуцировала различные структуры головы, в том числе нос, глаза, балансёры и слуховые пузырьки (органы слуха). Третий участок индуцировал только слуховые пузырьки, а самый задний участок дорсальную туловищную и хвостовую мезодерму. Продолжая изучение феномена индукции, Гольтфретер изготовил «сандвичи» из материала спинной губы бластопора, который поместил между двумя лоскутами недифференцированной эктодермы и культивировал их in vitro. Губа бластопора, взятая на стадии ранней гаструлы, индуцировала главным образом архенцефалические структуры, тогда как материал спинной губы бластопора, взятый на все более поздних стадиях, вызывал дифференцировку все более задних нейральных структур. Существует гипотеза, что региональная спецификация обусловлена действием двух веществ, секретируемых клетками хордомезодермы. Высокая концентрация одного из этих веществ обусловливает развитие переднего мозга, тогда как высокая концентрация другого индуцирует формирование спинного мозга и туловищных структур. Смесь этих двух веществ приводит к образованию среднего и заднего мозга (вспоминаем модель французского флага). разные участки цитоплазмы. Эти цитоплазматические участки содержат морфогенетические детерминанты, контролирующие детерминацию данной клетки и образование клеток определенного типа. Её изучают на примере оболочников, т.к. сразу после оплодотворения цитоплазма сегрегируется на несколько по-разному окрашенные области. Эти области распределяются по разным бластомерам. Первое деление дробления делит яйцо на правую и левую половину, а последующие протекают в каждой половине синхронно. Проследив деление бластомеров, было выявлено, что у каждой окрашенной области своя судьба – каждый из них дает начало определенному типу тканей (органов, групп органов). Многие структуры развиваются изолированно, однако нейральные – являются исключением. Они развиваются из двух передних квадрантов бластомеров – анимального и вегетативного, а при разделении не дают начала нервным клеткам. Таким образом даже в строго детерминированном зародыше оболочников существуют индукционные взаимодействия бластомеров.
Вопрос 23. |
Последнее изменение этой страницы: 2019-06-08; Просмотров: 355; Нарушение авторского права страницы





 Генотипическое и фенотипическое определение пола у человека
Генотипическое и фенотипическое определение пола у человека