|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
НЕЙРОФИЗИОЛОГИЯ СЛУХОВОЙ СИСТЕМЫ
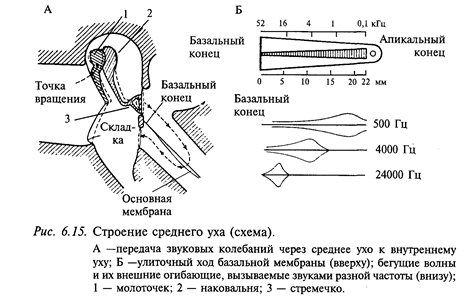
Мир, окружающий человека, полон звуков. Правда, человек слышит не все звуки этого мира, а только те, которые находятся в диапазоне от 20 до 20 000 Гц. Известно, что многие животные обладают значительно более широким диапазоном слышимых звуков. Например, дельфины «слышат» звуки частотой до 170 000 Гц, собаки и кошки также имеют более широкий диапазон слышимых звуков. Но слуховая система человека предназначена преимущественно для того, чтобы слышать речь другого человека, и в этом отношении ее совершенство нельзя даже близко сравнивать со слуховыми системами других млекопитающих. Наружное ухо включает ушную раковину и наружный слуховой проход. Рельеф ушной раковины играет значительную роль в восприятии звуков. Если, например, этот рельеф уничтожить, залив воском, человек заметно хуже определяет направление источника звука. Наружный слуховой проход человека в среднем имеет длину около 9 см. Есть данные, что трубка такой длины и схожего диаметра имеет резонанс на частоте около 1 кГц, другими словами, звуки этой частоты немного усиливаются. Среднее ухо отделено от наружного барабанной перепонкой, которая имеет вид конуса с вершиной, обращенной в барабанную полость. Среднее ухо человека состоит из барабанной полости, которая соединена евстахиевой трубой с полостью глотки, и барабанной перепонки с тремя слуховыми косточками (молоточком, наковальней и стремечком), сочлененных между собой (рис. 6.15). Слуховые косточки действуют как рычаги, улучшая передачу звуковых колебаний из воздушной среды наружного слухового прохода в более плотную водную среду внутреннего уха. Здесь имеются также две маленькие мышцы - одна прикреплена к ручке молоточка, а другая - к стремечку (на рис. 6.15 не показаны). Сокращение этих мышц предотвращает слишком большие колебания косточек, вызванных громкими звуками. Это так называемый акустический рефлекс. Таким образом, основной функцией акустического рефлекса является защита улитки от повреждающей стимуляции. Внутреннее ухо включает два рецепторных аппарата: вестибулярный и слуховой. Вестибулярный аппарат был уже рассмотрен выше. Опишем слуховой орган. Костный канал улитки разделен двумя мембранами - основной, или базилярной, и рейснеровой - на три отдельных канала, или лестницы: барабанную, вестибулярную и среднюю. Каналы внутреннего уха заполнены жидкостями, ионный состав которых в каждом канале специфичен. Средняя лестница заполнена эндолимфой с высоким содержанием ионов калия; она сообщается с эндолимфой вестибулярного органа. Две другие лестницы заполнены перилимфой, состав которой не отличается от тканевой жидкости. Внутри лестницы имеется еще один канал -кортиев туннель. По обе стороны от него находятся опорные клетки и ряды волосковых клеток (слуховые рецепторы), называемые соответственно внутренними и наружными клетками. Волосковые и опорные клетки, а также кортиев туннель покрыты соединительнотканной покровной (текториальной) мембраной, играющей важную роль в возбуждении слуховых рецепторов.
Эта структура (нейроэпителий и покровная мембрана), протянувшаяся по всей длине улитки, называется кортиевым органом (рис. 6.16). Вестибулярная и барабанная лестницы на вершине улитки соединяются через небольшое отверстие - геликотрему, средняя лестница заканчивается слепо (см. рис. 6.15). Средняя лестница, заполненная эндолимфой, заряжена положительно (до +80 мВ) относительно двух других лестниц. Если учесть, что потенциал покоя отдельных волосковых клеток около - 80 мВ, то в целом разность потенциала (эндокохлеарный потенциал) на участке средняя лестница - кортиев орган может составить около 160 мВ. Эндокохлеарный потенциал улитки чрезвычайно чувствителен к недостатку кислорода. Источником эндокохлеарного потенциала является, по-видимому, сосудистая полоска. Эндокохлеарный потенциал играет важную роль в возбуждении волосковых клеток. Предполагают, что волосковые клетки поляризованы этим потенциалом до критического уровня. В этих условиях минимальные механические воздействия могут вызвать возбуждение рецептора.
Звуковая волна действует на барабанную перепонку, и далее через систему косточек звуковое давление передается на овальное окно и воздействует на перилимфу вестибулярной лестницы (см. рис. 6.15). Поскольку жидкость несжимаема, перемещение перилимфы может передаваться через геликотрему в барабанную лестницу, а оттуда через круглое окно - обратно в полость среднего уха. Перилимфа может перемещаться и более коротким путем: рейснерова мембрана изгибается, и через среднюю лестницу давление передается на основную мембрану, затем в барабанную лестницу и через круглое окно в полость среднего уха. Именно в последнем случае раздражаются слуховые рецепторы. Колебания основной мембраны приводят к смещению волосковых клеток относительно покровной мембраны и, следовательно, к их деполяризации и далее к возбуждению волокон слухового нерва. Венгерский ученый Г. Бекеши (1951) предложил «теорию бегущей волны», позволяющую понять, как звуковая волна определенной частоты возбуждает волосковые клетки, находящиеся в определенном месте основной мембраны. Эта теория получила всеобщее признание. Основная мембрана расширяется от основания улитки к ее вершине примерно в 10 раз (у человека от 0, 04 до 0, 5 мм). Предполагается, что основная мембрана закреплена только по одному краю, остальная ее часть свободно скользит, что соответствует морфологическим данным. Теория Бекеши объясняет механизм анализа звуковой волны следующим образом: высокочастотные колебания проходят по мембране лишь короткое расстояние, а длинные волны распространяются далеко. Тогда начальная часть основной мембраны служит высокочастотным фильтром, а длинные волны проходят весь путь до геликотремы. Соединив все точки максимального перемещения колеблющейся мембраны, можно получить воображаемую поверхность, которую называют огибающей колебания. На рис. (6.15) показаны в поперечном разрезе огибающие колебаний основной мембраны для разных частот. Чистые тоны имеют огибающие с одним максимумом, от которого пологий спуск ведет к основанию, а более крутой - к верхушке улитки. Максимальные перемещения для разных частот происходят в разных точках основной мембраны: чем ниже тон, тем ближе его максимум к верхушке улитки. Таким образом, высота звука кодируется местом на основной мембране. Физиология путей и центров слуховой системы. Нейроны 1-го порядка (биполярные нейроны) находятся в спиральном ганглии, который расположен параллельно кортиеву органу и повторяет завитки улитки. Один отросток биполярного нейрона образует синапс на слуховом рецепторе, а другой направляется к головному мозгу, образуя слуховой нерв. Волокна слухового нерва выходят из внутреннего слухового прохода и достигают головного мозга в области так называемого мостомозжечкового угла (это анатомическая граница между продолговатым мозгом и мостом). -
Нейроны 2-го порядка образуют в продолговатом мозге комплекс слуховых ядер. При описании будем исходить из упрощенной анатомической схемы, согласно которой этот комплекс делят на дорсальный и вентральный, который, в свою очередь, состоит из переднелатеральной и заднелатеральной частей. В каждом из этих трех подразделений слуховых ядер имеется самостоятельное представительство кортиева органа (рис. 6.17). Как видно на рисунке, продвижение регистрирующего микроэлектрода от дорсального ядра к вентральному обнаруживает нейроны с постепенно убывающими значениями характеристической частоты (определение см. ниже). Это означает, что соблюдается принцип тонотопической организации. Таким образом, частотная проекция кортиева органа в целом упорядоченно повторяется в пространстве каждого из подразделений слухового комплекса ядер. Аксоны нейронов слуховых ядер поднимаются в лежащие выше структуры слухового анализатора как ипси -, так и контралатерально. Следующий уровень слуховой системы находится на уровне моста и представлен ядрами верхней оливы (медиальным и латеральным) и ядром трапециевидного тела. На этом уровне уже осуществляется бинауральный (от обоих ушей) анализ звуковых сигналов. Проекции слуховых путей на указанные ядра моста организованы также тонотопически. Большинство нейронов ядер верхней оливы возбуждаются бинаурально. Обнаружены две категории бинауральных нейронов. Одни возбуждаются звуковыми сигналами от обоих ушей (ВВ-тип), другие возбуждаются от одного уха, но тормозятся от другого (ВТ-тип). Ядро трапециевидного тела получает преимущественно контралатеральную проекцию от комплекса слуховых ядер, и в соответствии с этим нейроны реагируют преимущественно на звуковую стимуляцию контралатерального уха. В этом ядре также обнаруживается тонотопия. Аксоны клеток слуховых ядер моста идут в составе латеральной петли. Основная часть его волокон (в основном от медиальной оливы) переключается в нижнем двухолмии, другая часть идет в таламус и заканчивается на нейронах внутреннего (медиального) коленчатого тела, а также в переднем двухолмии (рис. 6.18). Кроме того, часть волокон латеральной петли иннервирует контралатеральный холмик нижнего двухолмия, образуя комиссуру Пробста. Нижнее двухолмие, расположенное на дорсальной поверхности среднего мозга, является важнейшим центром анализа звуковых сигналов. На этом уровне, по-видимому, заканчивается анализ звуковых сигналов, необходимых для ориентировочных реакций на звук. Основная часть клеточных элементов заднего холма локализована в центральном ядре. Аксоны клеток заднего холма направляются в составе его ручки к медиальному коленчатому телу. Однако часть аксонов идет к противоположному холму, образуя интеркаликулярную комиссуру. Медиальное коленчатое тело является таламическим центром слуховой системы. В нем различают крупноклеточную и мелкоклеточную (основную) части. Аксоны нейронов мелкоклеточной части коленчатого тела образуют акустическую радиацию и направляются в слуховую область коры. Крупноклеточная часть внутреннего коленчатого тела получает проекции от нижнего двухолмия. В этом таламическом ядре также прослеживается тонотопия: низкая частота представлена в латеральной, а высокая- в медиальной части ядра. Слуховая кора представляет высший центр слуховой системы ц располагается в височной доле. У человека в ее состав входят поля 41, 42 и частично 43. В каждой из зон имеет место тонотопия, т. е полное представительство нейроэпителия кортиева органа. Пространственное представительство частот, в слуховых зонах сочетается с колончатой организацией слуховой коры, особенно выраженной в первичной слуховой коре.
Частотно-пороговые характеристики нейронов слуховой системы. Как было описано выше, все уровни слуховой системы млекопитающих имеют тонотопический принцип организации. Другая важная характеристика нейронов слуховой системы - способность избирательно реагировать на определенную высоту звука.
У всех животных имеется соответствие между частотным диапазоном издаваемых звуков и аудиограммой, которая характеризует слышимые звуки. Частотную избирательность нейронов слуховой системы описывают частотно-пороговой кривой (ЧПК), отражающей зависимость порога реакции нейрона от частоты тонального стимула. Частота, при которой порог возбуждения данного нейрона минимальный, называется характеристической частотой. ЧПК волокон слухового нерва имеет V-образную форму с одним минимумом, который соответствует характеристической частоте данного нейрона. ЧПК слухового нерва имеет заметно более острую настройку по сравнению с амплитудно-частотными кривыми основной мембраны (рис. 6.19). Предполагают, что в обострении частотно-пороговой кривой участвуют эфферентные влияния уже на уровне слуховых рецепторов (вспомните, что волосковые рецепторы являются вторично-чувствующими и получают эфферентные волокна). Кодирование интенсивности звука. Для большинства нейронов разных уровней слуховой системы характерно увеличение их реакций при росте интенсивности стимула в широком диапазоне (от. О до 120 дБ). Если учесть, что одиночные нейроны широко распределены по порогам возникновения реакций, то можно считать, что интенсивность звука кодируется не только частотой импульсации, но и количеством возбужденных нейронов. Поэтому считают, что плотность потока импульсации является нейрофизиологическим коррелятом громкости. Вместе с тем в центральных отделах слуховой системы обнаружены нейроны, обладающие определенной избирательностью к интенсивности звука, т.е. реагирующие на довольно узкий диапазон интенсивности звука. Нейроны с такой реакцией впервые появляются на уровне слуховых ядер. На более высоких уровнях слуховой системы их количество возрастает. Диапазон выделяемых ими интенсивностей суживается, достигая минимальных значений у нейронов коры. Предполагают, что такая специализация нейронов отражает последовательный анализ интенсивности звука в слуховой системе. Вопросы 1. Строение и физиология наружного и среднего уха. 2. Строение и физиология внутреннего уха. 3. Нейроанатомическая схема слуховой системы человека. 4. Частотно-пороговые характеристики нейронов слуховой системы. Литература Гильберт С. Слух. Введение в психологическую и физиологическую акустику. М.: Медицина, 1984. Популярное:
|
Последнее изменение этой страницы: 2016-03-22; Просмотров: 1011; Нарушение авторского права страницы