|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Математические модели в экологии
Основные вопросы, рассматриваемые на лекции: 1. Классификация моделей. 2. Экологическое прогнозирование и мониторинг.
Классификация моделей При экологическом исследовании, которое обычно проводится на определенном количестве особей, изучаются природные явления во всем их многообразии: общие закономерности, присущие макросистеме, ее реакции на изменения условий существования и др. Но каждая особь, индивидуум неодинаковы, отличны друг от друга. Кроме того, выбор особи из всей популяции носит случайный характер. И лишь применение методов математической статистики дает возможность по случайному набору различных вариантов определить достоверность тех или иных результатов (степень отклонения их от нормы, случайны отклонения или закономерны) и получить объективное представление обо всей популяции. Однако, все биологические системы, обладают способностью к саморегуляции и ограничиваться только методами математической статистики стало невозможно. Поэтому в современной экологии широко применяются методы теории информации и кибернетики, тесно связанные с такими областями математики, как теория вероятности, математическая логика, дифференциальные и интегральные исчисления, теория чисел, матричная алгебра. В последнее время широкое распространение получило моделирование биологических явлений, т. е. воспроизведение в искусственных системах различных процессов, свойственных живой природе. Так, в «модельных условиях» были осуществлены многие реакции, протекающие в растении при фотосинтезе. Примером биологических моделей может служить и аппарат искусственного кровообращения, искусственная почка, искусственные легкие, протезы, управляемые биотоками мышц, и др. В различных областях биологии широко применяются так называемые живые модели. Несмотря на то, что различные организмы отличаются друг от друга сложностью структуры и функции, многие биологические процессы у них протекают практически одинаково. Поэтому изучать их удобно на более простых существах. Они то и становятся живыми моделями. В качестве примера можно привести зоохлореллу, которая служит моделью для изучения обмена веществ; моделью для исследования внутриклеточных процессов являются гигантские растительные и животные клетки и т. д. Основной задачей биологического моделирования является экспериментальная проверка гипотез относительно структуры и функции биологических систем. Сущность этого метода заключается в том, что вместе с оригиналом, т. е. с какой-то реальной системой, изучается его искусственно созданное подобие — модель. В сравнении с оригиналом модель обычно упрощена, но свойства их сходны. В противном случае полученные результаты могут оказаться недостоверными, не свойственными оригиналу. Т. Г. Гильманов (1980) указывает: «Одно из достоинств метода моделирования состоит в возможности построения моделей с «удобной реализацией», ибо удачный выбор реализации делает исследование модели несравненно более легким, чем исследование оригинала, и в то же время позволяет сохранить существенные черты его состава, структуры и функционирования». В зависимости от особенностей оригинала и задач исследования применяются самые разнообразные модели (рисунок 1). Реальные (натурные, аналоговые) модели, если таковые удается создать, отражают самые существенные черты оригинала. Например, аквариум может служить моделью естественного водоема. Однако создание реальных моделей сопряжено с большими техническими трудностями, так как пока еще не удается достичь точного воспроизведения оригинала. Знаковая модель представляет собой условное отображение оригинала с помощью математических выражений или подробного описания. Наибольшее распространение в современных экологических исследованиях получили концептуальные и математические модели и их многочисленные разновидности. Разновидности концептуальных моделей характеризуются подробным описанием системы (научный текст, схема системы, таблицы, графики и т.д.). Математические модели являются более эффективным методом изучения экологических систем, особенно при определении количественных показателей. Математические символы, например, позволяют сжато описать сложные экологические системы, а уравнения дают возможность формально определить взаимодействия различных их компонентов.
Рисунок 1- Классификация моделей (по В. Д. Федорову и Т. Г. Гильманову, 1980)
Процесс перевода физических или биологических представлений о любой экологической системе в ряд математических зависимостей и операции над ними называются системным анализом, а сама математическая система — моделью. Следует отметить, что математические модели являются неполным абстрактным отображением реального мира. По типу реализации различаются реальные и знаковые модели. Реальная модель отражает существенные черты оригинала уже по самой природе своей физической реализации. Например, аквариум с его растительностью, животным и микробным населением воспроизводит некоторые черты обитаемых природных водоемов уже потому, что он сам является населенным водоемом, хотя и значительно меньших размеров. Одна из наиболее сложных проблем, с которой приходится сталкиваться при работе с реальными (натурными) моделями, заключается в трудности установления степени адекватности модели оригиналу и, следовательно, в обосновании возможности применения результатов моделирования к исходной системе-оригиналу. В отличие, например, от аэро- или гидродинамики, где разработаны количественные критерии адекватности моделей (критерий Рейнольдса и т.п.), в результате чего стало возможным успешное применение натурных моделей для решения широкого круга научных и конструкторских задач в указанных областях, при натурном моделировании экосистем вопросы обоснования адекватности еще очень далеки от удовлетворительного решения. Кроме того, создание и использование натурных моделей экосистем связано с известными трудностями технического характера, преодоление которых, из-за отсутствия гарантий адекватности, отнюдь не всегда приводит к решению поставленной задачи. В отличие от реальной, знаковая модель представляет собой условное описание системы-оригинала с помощью данного алфавита символов и операций над символами, и результате чего получаются слова и предложения некоторого языка, которые с помощью определенного кода интерпретируются как образы некоторых свойств элементов системы-оригинала и связей между ними. Как отмечает И. А. Полетаев (1966), знаковые модели несравненно богаче возможностями, чем реальные, ибо они почти не связаны ограничениями физической реализации. Наибольшее значение для экологии имеют две разновидности знаковых моделей: это, во-первых, так называемые концептуальные и, во-вторых, математические модели. Концептуальная модель представляет собой несколько более формализованный и систематизированный вариант традиционного естественнонаучного описания изучаемой экосистемы, состоящей из научного текста, сопровождаемого блок-схемой системы, таблицами, графиками и прочим иллюстративным материалом. Несколько тавтологичный, но широко применяемый термин “концептуальная модель” подчеркивает, что назначение этой модели — служить ясным, обобщенным и в то же время достаточно полным выражением знаний и представлений исследователя об изучаемой системе в рамках и средствами определенной научной концепции. Например, в рамках “энергетической” или “биогеохимической” концепции соответствующие концептуальные модели принимают форму блок-схем трофических связей или потоков вещества в экосистеме, которые сопровождаются поясняющим текстовым, табличным и графическим материалом, раскрывающим состав, структуру и некоторые аспекты функционирования экосистемы. В то же время наряду с такими общеизвестными достоинствами концептуальных моделей, как универсальность, гибкость, богатство средств выражения и др., благодаря которым этот метод применяется к самым разным системам, ему свойственны и недостатки, как, например, высокая неоднозначность интерпретации и известная статичность, затрудняющая описание динамических систем. При количественном изучении динамики экосистем гораздо более эффективны методы математического моделирования. В качестве примера рассмотрим идеализированную систему, состоящую из одной популяции, которая существует в условиях изобилия корма и при отсутствии врагов и паразитов. Предположим, что в этих условиях прирост популяции пропорционален достигнутой численности, причем удельная скорость прироста r зависит от единственного внешнего фактора — температуры окружающей среды, которая на рассматриваемом промежутке времени t0≤ t≤ tN считается известной. Чтобы построить математическую модель такой системы, рассмотрим множество V, состоящее из одного элемента — входной функции v(t), задающей динамику температуры окружающей среды при t0≤ t≤ tN, а также множество X, тоже состоящее из одного элемента, действительной переменной х(t), обозначающей численность популяции в момент времени t. Структуру модели образуют три математических соотношения:
Первое выражает линейную зависимость скорости роста популяции от ее численности с меняющимся во времени коэффициентом удельного прироста r(t). Второе служит математическим выражением зависимости r от температуры окружающей среды v (функция Для нахождения функции модели, т. е. разрешающего оператора F системы (1), подставим в первое уравнение выражение r через v с помощью функции
Разделяя переменные:
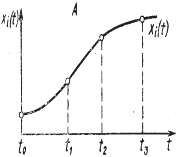
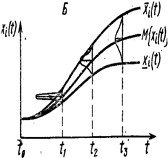
и интегрируя от х0 до х и от t0 до t соответственно, где х0=х(t0) и х=х(t), получаем ln|x|-ln|х0|=и(v(t))dt (4) После потенцирования получаем окончательный результат: х(t)=х0exp{ Таким образом, функционирование модели выражается оператором х(t)=F(v, х0, t)=х0exp{ В зависимости от свойств разрешающего оператора F математические модели динамичных систем классифицируются по разным признакам. Так, если для оператора F найдено точное аналитическое выражение, позволяющее для любых входных функций и начальных условий непосредственно определять значение переменных состояния x1, …, xn в любой нужный момент t, то модель принято называть аналитической. Аналитические модели обладают многими благоприятными свойствами, облегчающими их исследование и применение. Однако в подавляющем большинстве случаев нахождение аналитического выражения для разрешающего оператора F оказывается затруднительным или в принципе невозможным. Процедуру численного решения этих уравнений на ЭВМ, в результате чего получается реализация оператора F в виде машинной программы, с помощью которой по входным и начальным данным рассчитываются значения переменных состояний x1(t), …, xn(t) на интервале t0≤ t≤ tN. Такие модели называются численными, или, чаще, имитационными. В зависимости от степени определенности предсказания траектории (x1(t), …, xn(t)) оператором F модели делятся на детерминированные и стохастические (вероятностные). Если в детерминированной модели значения переменных состояния определяются однозначно (с точностью до ошибок вычисления) (рис. 2, А), то стохастическая модель для каждой переменной xi, дает распределение возможных значений, характеризуемое такими вероятностными показателями, как математическое ожидание М{xi}, среднее квадратическое отклонение σ (xi) и т. п. (рис. 2, Б).
Рисунок 2. Детерминированная (А) и стохастическая (Б) модели динамики переменной xi(t).
Для любого момента времени t детерминированная модель предсказывает единственное значение переменной xi(t), а стохастическая модель показывает интервал [xi(t), xi(t)], содержащий величину xi(t) и ее распределение на этом интервале (в частности, математическое ожидание М{xi(t)}, среднее квадратическое отклонение σ {xi(t), } и другие статистические показатели). По характеру временного описания динамики переменных состояния xi(t) различаются дискретные и непрерывные модели. Дискретная модель описывает поведение системы на фиксированной последовательности моментов времени t0< t1< …< ti< tN, тогда как в непрерывной модели значения переменных состояния могут быть рассчитаны для любой точки t рассматриваемого интервала [t0, tN]. Среди дискретных выделяются модели с фиксированным шагом по времени Δ t= ti-ti-1, который не может быть изменен без глубокой перестройки всей модели. Например, в моделях динамики популяции организмов с непрерывающимися поколениями, сменяющимися только один раз в год, принимается Δ t= 1 год (Варли, Градуэлл, Хасселл, 1978). В отличие от них существуют дискретные модели, в которых шаг по времени Δ t может неограниченно уменьшаться (в пределах возможностей используемой ЭВМ), вследствие чего такие модели по детальности описания временных изменений приближаются к непрерывным. В частности, таковы модели, получающиеся в результате дискретизации непрерывного описания изучаемой системы в процессе приближенного численного решения дифференциальных уравнений. Следующий важный признак, которому различаются математические модели экосистем, - это характер описания их пространственного строения. Модели, в которых пространственное строение экосистемы не рассматривается, т. е. в качестве переменных состояния фигурируют зависящие только от времени переменные xi(t),..., n, принято называть моделями с сосредоточенными значениями (или точечными моделями), в отличие от моделей с распределенными значениями, в которых переменные состояния xi зависят не только от времени, но и от пространственных координат (одной или нескольких). Например, при моделировании водной экосистемы в качестве переменных состояния можно взять усредненные по площади и суммированные по глубине значения биомасс популяций, запасов биогенных элементов и т. д., выраженные, например, в граммах под площадью зеркала воды в 1 м2 — это будет модель с сосредоточенными значениями (Алексеев, 1975). Если в модели учитывается гетерогенность по глубине (координата z), т. е. xi=xi(z, t) то получается (очевидно, более детальная) модель с распределенными значениями по глубине, но все еще осредненными по плоскости (х, у). При описании мелкого, хорошо перемешиваемого по вертикали, но гетерогенного по плоскости водоема целесообразно в качестве переменных состояния использовать функции вида xi=xi(x, y, t). Наконец, вводя все три пространственные координаты xi=xi(x, y, z, t), получают модель с пространственно распределенными значениями. Завершая краткое введение в метод моделирования, остановимся на некоторых способах визуального представления результатов моделирования. Наиболее часто используется так называемый способ развертки во времени, который состоит в построении таблиц или графиков изменения входных переменных и переменных состояния как функций времени t (рисунок 3, A ). При всех его достоинствах этот способ не всегда (особенно при большом числе переменных) дает наглядное представление взаимосвязей между переменными. Поэтому в дополнение к нему нередко используется так называемый способ фазовых портретов, когда на график наносится изображение траектории системы в пространстве состояний (при n=2 или 3) или проекции этой траектории на координатные плоскости (xi, xj), образованные различными парами координат при n> 3 (рисунок 3, Б). Время присутствует на фазовом портрете неявно, через указание тем или иным способом направления движения изображающей точки вдоль траектории (например, с помощью стрелок или отметок времени вдоль траектории). Соотношение способов развертки во времени и фазового портрета поясняет рисунок 3, В).
Рисунок 3 - Представление результатов моделирования способами развертки по времени (А) и фазового портрета (Б), соотношение этих двух способов в случае двухмерного фазового пространства (В).
В качестве примера рассмотрим простейшее дифференциальное уравнение, описывающее рост популяции какого-либо вида: dх/dt=rх, где х — плотность популяции в момент времени t; r — истинная скорость роста, принятая постоянной. Решением этого уравнения является функция х=х0еrt, где х0 — плотность популяции в момент времени t=0. Следовательно, если проводить наблюдение за большой выборкой особей в течение короткого периода времени Δ t, то доля особей, размножившихся в течение этого периода, будет равна rΔ t. Значит, возрастной состав популяции не изменяется во времени. Но это справедливо лишь для ограниченного периода времени, так как возрастная структура популяции будет приближаться к устойчивой лишь тогда, когда специфичные для каждой возрастной группы рождаемость и смертность остаются постоянными. Такого состояния популяция может достичь только при постоянстве условий среды, избытке необходимых ресурсов и отсутствии эволюционных процессов (изменений). Но в естественных условиях этого никогда не наблюдается. Как видно, при отсутствии реальных моделей математический подход становится все более отвлеченным. При исключении же математического подхода часто бывает трудно уловить общий смысл реальной модели. Вот почему в современной экологии реальные и знаковые модели используются параллельно, дополняя и обогащая друг друга. Первыми математическими моделями простейших экологических систем хищник — жертва и паразит — хозяин были теоретические разработки В. Вольтерры, сделанные в 1931 г. Они послужили толчком для построения более сложных моделей процессов пищевых отношений популяций в биоценозах. С появлением быстродействующих электронно-вычислительных машин возникли возможности моделирования еще более сложных саморегулирующихся систем с обратной связью — популяций, биоценозов и биогеоценозов. На основе математического моделирования успешно изучаются микробные популяции и популяции одноклеточных водорослей, выращиваемых в культиваторах, исследуются явления внутривидовой конкуренции и различные формы межвидовых взаимоотношений. Важное место занимает попытка математического моделирования в экологических исследованиях, направленных на использование ресурсов природы так, чтобы в ней осуществлялось естественное их самовоспроизводство. Для этого нужно не только знать сущность основных взаимодействий и процессов, протекающих в биогеоценозах, в биосфере в целом, но и с помощью математических методов прогнозировать их хотя бы на ближайшее будущее. Экологическое прогнозирование и мониторинг В современных условиях моделирование занимает основное место в работах по экологическому прогнозированию. Как отмечает А. А. Ляпунов, «задачи изучения математических моделей биогеценозов состоят в том, чтобы сопоставить кинетику явлений, подсказываемую моделью, с той, которая наблюдается в действительности, и, опираясь на это сопоставление, довести модель до достаточно хорошего согласия с действительностью. Тогда такие модели станет возможным использовать для практических прогнозов, а также для выбора рационального вмешательства человека в жизнь природы, с тем чтобы обеспечить такое использование природных ресурсов, при котором бы они должным образом самовоспроизводились». В основу математического моделирования при экологическом прогнозировании положен принцип представления сложной биологической системы в виде отдельных подсистем (блоков, модулей, камер), связанных между собой функциональными связями, имитирующими либо поток веществ (в том числе и загрязняющих), либо регулирующиеся воздействия, либо пространственные миграции, либо развитие организмов и т. д. Например, в водной экосистеме выделяются как отдельные подсистемы: рыбы разного возраста, разные виды фитопланктона, зоопланктона, бентос, донные отложения, поступающие в водоем стоки и т. д. Кроме того, можно выделять и функциональные зависимости: плодовитость рыб в норме и при определенных воздействиях, например свинца или других веществ; плодовитость и продолжительность жизни фито- и зоопланктона, бентоса; зависимость от температуры среды выклевывания мальков из икры, времени нереста и т. д. При этом экологический прогноз с помощью математических моделей возможен только при наличии данных о «нулевой точке отсчета», т. е. о не нарушенной природой системе. В последние годы среди прогнозистов широко распространилось понятие «мониторинг», которое было предложено еще в 1920-х гг. по отношению к окружающей среде. Часто за мониторингом признается только наблюдение за состоянием окружающей среды. В более широком смысле в понятие «мониторинг» включают не только наблюдение, а также контроль и управление состоянием окружающей среды, т. е. то или иное целенаправленное воздействие на нее. Таким образом, мониторинг слагается из двух последовательных этапов: от наблюдения до контроля и управления. Ограничение мониторинга наблюдением придает ему лишь пассивное, информационное значение. Задачей современного мониторинга является прогноз. Программа фонового экологического мониторинга реализуется в биосферных заповедниках. Схема фонового мониторинга загрязнения природной среды и его экологических последствий четко подразделяется на геохимический, геофизический и биологический мониторинги. Цель биохимического и геофизического мониторингов — получение информации об уровне загрязнения окружающей среды, об интенсивности и характере воздействий загрязнения на биологические системы, а также прогнозирование уровней загрязнения биоты в будущем. Цель биологического мониторинга — оценка и прогноз реакции экосистем в ответ на фоновые воздействия загрязнителей. В его основу положены разработки математических моделей экосистем, которые могли бы служить инструментом прогнозов изменения состояния экосистем на фоновом глобальном уровне. Такие модели позволяют строить прогнозы о том, когда и под влиянием каких уровней воздействия загрязнителей исчезнут наиболее чувствительные к загрязнениям виды, установить очередность их исчезновения или существенного снижения численности популяций в той или иной экосистеме. Примером успешного применения математической модели для прогнозирования является разработанная В. В. Меншуткиным модель динамики численности лососевых рыб. Анализ модели на ЭВМ позволил выяснить и понять важнейшие закономерности жизнедеятельности камчатской нерки. На модели был проведен ряд экспериментов. В результате установили, что на нерестилищах нерки наблюдается повышенная смертность мальков, а в открытом море катастрофически возрос промысел лососей. Модель явилась официальным документом при составлении прогноза. Ограничения вылова лососей в открытом море позволили сохранить стадо этих ценных рыб. В настоящее время модели чаще всего используются для решения конкретных сегодняшних задач: применять или не применять пестициды, какую природную среду требуется контролировать в первую очередь, какие антропические нагрузки допустимы и т. д. Хотя все эти сегодняшние проблемы невозможно решать без обоснованного прогноза на далекую перспективу.
Лекция № 4 Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 2030; Нарушение авторского права страницы

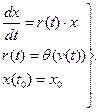 (1)
(1) (v) считается известной). Третье задает начальную численность популяции при t=t0
(v) считается известной). Третье задает начальную численность популяции при t=t0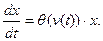 (2)
(2) (3)
(3)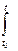 и(v(t))dt} (5)
и(v(t))dt} (5)