
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Нервные пути и связи в зрительной системе
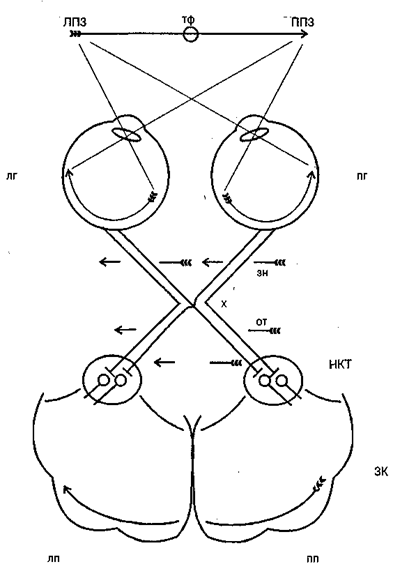
Из сетчатки зрительная информация по волокнам зрительного нерва устремляется в мозг. Нервы от двух глаз встречаются у основания мозга, где часть волокон переходит на противоположную сторону (зрительный перекрест, или хиазма). Это обеспечивает каждое полушарие мозга информацией от обоих глаз: в затылочную долю правого полушария поступают сигналы от правой половины каждой сетчатки, а в левое полушарие — от левой (рис. 4.3). После хиазмы зрительные нервы называются оптическими трактами и основное количество их волокон приходит в подкорковый зрительный центр — наружное коленчатое тело (НКТ). Отсюда зрительные сигналы поступают в первичную проекционную область зрительной коры (стриарная кора, или поле 17 по Бродма-ну). Зрительная кора состоит из ряда полей, каждое из которых обеспечивает свои, специфические функции, получая как прямые, так и опосредованные сигналы от сетчатки и в общем сохраняя ее топологию, или ретинотопию (сигналы от соседних участков сетчатки попадают в соседние участки коры).
Рис. 4.3. Схема зрительных путей от сетчатки глаза до первичной зрительной коры: ЛПЗ — левое поле зрения; ППЗ — правое поле зрения; тф — точка фиксации взора; лг — левый глаз; т — правый глаз; зн — зрительный нерв; х — зрительный перекрест, или хиазма; от — оптический тракт; НКТ — наружное коленчатое тело; ЗК — зрительная кора; лп — левое полушарие; пп — правое полушарие
Электрическая активность центров зрительной Системы
При действии света в рецепторах, а затем и в нейронах сетчатки генерируются электрические потенциалы, отражающие параметры действующего раздражителя (рис. 4.4, А, а). Суммарный электрический ответ сетчатки глаза на свет называют электроретинограммой (ЭРГ). Она может быть зарегистрирована от целого глаза: один электрод помещают на поверхность роговой оболочки, а другой — на кожу лица вблизи глаза (либо на мочку уха). В ЭРГ хорошо отражаются интенсивность, цвет, размер и длительность действия светового раздражителя. Поскольку в ЭРГ отражена активность почти всех клеток сетчатки (кроме ганглиозных клеток), этот показатель широко используется для анализа работы и диагностики заболеваний сетчатки.
Рис. 4.4. Электроретинограмма (Л) и вызванный светом потенциал (ВП) зрительной коры (Б): a, b, c, d нa A — волны ЭРГ; стрелками указаны моменты включения света. P1-P5 - позитивные волны BП, N1-N5 — негативные волны ВП на Б
Возбуждение ганглиозных клеток сетчатки приводит к тому, что по их аксонам (волокнам зрительного нерва) в мозг устремляются электрические импульсы. Ганглиозная клетка сетчатки — это первый в сетчатке нейрон «классического» типа, генерирующий распространяющиеся импульсы. Описано три основных типа ганглиозных клеток: отвечающие на включение света (оn-реакция), его выключение (оff-реакция) и на то и другое (оn-off-реакция). В центре сетчатки рецептивные поля ганглиозных клеток маленькие, а на периферии сетчатки они значительно больше по диаметру. Одновременное возбуждение близко расположенных ганглиозных клеток приводит к их взаимному торможению: ответы каждой клетки становятся меньше, чем при одиночном раздражении. В основе этого эффекта лежит латеральное или боковое торможение (см. гл. 3). Благодаря круглой форме рецептивные поля ганглиозных клеток сетчатки производят так называемое поточечное описание сетчаточного изображения: оно отображается очень тонкой дискретной мозаикой, состоящей из возбужденных нейронов. Нейроны подкоркового зрительного центра (НКТ) возбуждаются, когда к ним приходят импульсы из сетчатки по волокнам зрительного нерва. Рецептивные поля этих нейронов также круглые, но меньшего размера, чем в сетчатке. Пачки импульсов, генерируемые ими в ответ на вспышку света, короче, чем в сетчатке. На уровне НКТ происходит взаимодействие афферентных сигналов, пришедших из сетчатки, с эфферентными сигналами из зрительной коры, а также из ретикулярной формации от слуховой и других сенсорных систем. Это взаимодействие помогает выделять наиболее существенные компоненты сигнала и, возможно, участвует в организации избирательного зрительного внимания (см. гл. 9). Импульсные разряды нейронов НКТ по их аксонам поступают в затылочную часть полушарий головного мозга, в которой расположена первичная проекционная область зрительной коры (стриарная кора). Здесь у приматов и человека происходит значительно более специализированная и сложная, чем в сетчатке и в НКТ, переработка информации. Нейроны зрительной коры имеют не круглые, а вытянутые (по горизонтали, вертикали или по диагонали) рецептивные поля (рис. 4.5) небольшого размера [Хьюбел, 1990].
Рис. 4.5. Рецептивное поле нейрона зрительной коры мозга кошки (А) и ответы этого нейрона на вспыхивающие в рецептивном поле световые полоски разной ориентации (Б): А — плюсами отмечена возбудительная зона рецептивного поля, а минусами — две боковые тормозные зоны; Б — видно, что этот нейрон наиболее сильно реагирует на вертикальную и близкую к ней ориентацию
Благодаря этому они способны выделять из изображения отдельные фрагменты линий с той или иной ориентацией и расположением и избирательно на них реагировать (детекторы ориентации). В каждом небольшом участке зрительной коры по ее глубине сконцентрированы нейроны с одинаковой ориентацией и локализацией рецептивных полей в поле зрения. Они образуют ориентационную колонку нейронов, проходящую вертикально через все слои коры. Колонка — пример функционального объединения корковых нейронов, осуществляющих сходную функцию. Группа соседних ориентационных колонок, нейроны которых имеют перекрывающиеся рецептивные поля, но разные предпочитаемые ориентации, образует так называемую сверхколонку. Как показывают исследования последних лет, функциональное объединение отдаленных друг от друга нейронов зрительной коры может происходить также за счет синхронности их разрядов. Недавно в зрительной коре найдены нейроны с избирательной чувствительностью к крестообразным и угловым фигурам, относящиеся к детекторам второго порядка. Таким образом, начала заполняться «ниша» между описывающими пространственные признаки изображения простыми ориентационными детекторами и детекторами высшего порядка (лица), найденными в височной коре. В последние годы хорошо исследована так называемая «пространственно-частотная» настройка нейронов зрительной коры [Глезер, 1985; Физиол. зрения, 1992]. Она заключается в том, что многие нейроны избирательно реагируют на появившуюся в их рецептивном поле решетку из светлых и темных полос определенной ширины. Так, имеются клетки, чувствительные к решетке из мелких полосок, т. е. к высокой пространственной частоте. Найдены клетки с чувствительностью к разным пространственным частотам. Считается, что это свойство обеспечивает зрительной системе способность выделять из изображения участки с разной текстурой [Глезер, 1985]. Многие нейроны зрительной коры избирательно реагируют на определенные направления движения (дирекциональные детекторы) либо на какой-то цвет (цве-тооппонентные нейроны), а часть нейронов лучше всего отвечает на относительную удаленность объекта от глаз. Информация о разных признаках зрительных объектов (форма, цвет, движение) обрабатывается параллельно в разных частях зрительной коры.
Сравнение вызванного световой вспышкой ответа сетчатки (ЭРГ) и ВП коры позволяет оценить работу проекционного зрительного пути и установить локализацию патологического процесса в зрительной системе.
Световая чувствительность
Абсолютная чувствительность зрения. Чтобы возникло зрительное ощущение, свет должен обладать некоторой минимальной (пороговой) энергией. Минимальное количество квантов света, необходимое для возникновения ощущения света в темноте, колеблется от 8 до 47. Одна палочка может быть возбуждена всего 1 квантом света. Таким образом, чувствительность рецепторов сетчатки в наиболее благоприятных условиях световос-приятия предельна. Одиночные палочки и колбочки сетчатки различаются по световой чувствительности незначительно. Однако количество фоторецепторов, посылающих сигналы на одну ганглиозную клетку, в центре и на периферии сетчатки различно. Количество колбочек в рецептивном поле в центре сетчатки примерно в 100 раз меньше количества палочек в рецептивном поле на периферии сетчатки. Соответственно и чувствительность палочковой системы в 100 раз выше, чем у колбочковой.
Зрительная адаптация
При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной системы к условиям яркой освещенности называется световой адаптацией. Обратное явление (темновая адаптация) наблюдается, когда из светлого помещения человек переходит в почти не освещенное помещение. В первое время он почти ничего не видит из-за пониженной возбудимости фоторецепторов и зрительных нейронов. Постепенно начинают выявляться контуры предметов, а затем различаются и их детали, так как чувствительность фоторецепторов и зрительных нейронов в темноте постепенно повышается. Повышение световой чувствительности во время пребывания в темноте происходит неравномерно: в первые 10 мин она увеличивается в десятки раз, а затем, в течение часа, — в десятки тысяч раз. Важную роль в этом процессе играет восстановление зрительных пигментов. Так как в темноте чувствительны только палочки, слабо освещенный предмет виден лишь периферическим зрением. Существенную роль в адаптации, помимо зрительных пигментов, играет переключение связей между элементами сетчатки. В темноте площадь возбудительного центра рецептивного поля ганглиозной клетки увеличивается из-за ослабления кольцевого торможения, что приводит к увеличению световой чувствительности. Световая чувствительность глаза зависит и от влияний, идущих со стороны мозга. Освещение одного глаза понижает световую чувствительность неосвещенного глаза. Кроме того, на чувствительность к свету оказывают влияние также звуковые, обонятельные и вкусовые сигналы.
Популярное:
|
Последнее изменение этой страницы: 2016-03-26; Просмотров: 1283; Нарушение авторского права страницы


 Для оценки передачи сигналов на разных уровнях зрительной системы часто используют регистрацию суммарных вызванных потенциалов (ВП), которые у человека можно одновременно отводить от сетчатки и от зрительной коры (см. рис. 4.4, Б).
Для оценки передачи сигналов на разных уровнях зрительной системы часто используют регистрацию суммарных вызванных потенциалов (ВП), которые у человека можно одновременно отводить от сетчатки и от зрительной коры (см. рис. 4.4, Б).