
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Структура и функции внутреннего уха
Во внутреннем ухе находится улитка, содержащая слуховые рецепторы. Улитка представляет собой костный спиральный канал, который по всей длине разделен вестибулярной и основной мембранами на три хода: верхний, средний и нижний (рис. 4.7). Полость среднего канала не сообщается с полостью других каналов и заполнена эндолимфой, а верхний и нижний каналы сообщаются друг с другом и заполнены перилимфой. Внутри среднего канала улитки на основной мембране расположен спиральный (кортиев) орган, содержащий рецепторные клетки, которые трансформируют механические колебания в электрические потенциалы.
Рис. 4.7. Поперечный разрез завитка улитки с увеличенной частью спирального (кортиева) органа, очерченной сверху прямоугольником
Колебания мембраны овального окна вызывают колебания перилимфы в верхнем и нижнем каналах, кроме того, начинает колебаться и основная мембрана. На ней расположены два вида рецепторных волосковых клеток: внутренние и наружные.
4.3.3. Анализ частоты звука (высоты тона)
При действии звуков разной частоты возбуждаются разные рецепторные клетки кортиева органа. В улитке сочетаются два типа кодирования высоты звука: пространственный и временной [Сомьен, 1975]. Пространственное кодирование основано на определенном расположении возбужденных рецепторов на основной мембране. При действии низких и средних тонов кроме пространственного осуществляется и временное кодирование: частота следования импульсов в волокнах слухового нерва повторяет частоту звуковых колебаний. Нейроны всех уровней слуховой системы настроены на определенную частоту и интенсивность звука. Для каждого нейрона может быть найдена оптимальная частота звука, на которую порог его реакции минимален. Частотно-пороговые кривые разных клеток не совпадают, в совокупности перекрывая весь частотный диапазон слышимых звуков, что обеспечивает их полноценное восприятие. Анализ интенсивности звука. Сила звука кодируется частотой импульсации и числом возбужденных нейронов. При слабом стимуле в реакцию вовлекается лишь небольшое количество наиболее чувствительных нейронов, а при усилении звука в реакции участвует все большее количество дополнительных нейронов с более высокими порогами.
Слуховые ощущения
Тональность (частота) звука. Человек воспринимает звуковые колебания с частотой от 16 до 20 000 Гц. Этот диапазон соответствует 10-11 октавам. Верхняя граница частоты воспринимаемых звуков зависит от возраста: она постепенно понижается (в старости часто не слышат высоких тонов). Различение частоты звука характеризуется тем минимальным различием по частоте двух близких звуков, которое еще улавливается человеком. При низких и средних частотах человек способен заметить различия в 1-2 Гц. Встречаются люди с абсолютным слухом: они способны точно узнавать и обозначать любой звук даже при отсутствии звука сравнения.
Рис. 4.8. Область звукового восприятия человека. Зависимость пороговой интенсивности звука (ось ординат — звуковое давление в дин/см) от частоты тональных звуков (ось абсцисс в Гц). Кривая AEFGD — абсолютные пороги восприятия; ABCD — пороги болевого ощущения при действии громких звуков
Громкость звука. Кажущуюся громкость звука следует отличать от его физической силы. Ощущение громкости не идет строго параллельно нарастанию интенсивности звучания. Единицей громкости звука является бел. Эта единица представляет собой десятичный логарифм отношения действующей интенсивности звука/ к пороговой его интенсивности /0. На практике обычно используется в качестве единицы громкости децибел (дБ), т. е. 0, 1 бела. Дифференциальный порог по громкости в среднем диапазоне слышимых частот (1000 Гц) составляет всего 0, 59 дБ, а на краях шкалы частот доходит до 3 дБ. Максимальный уровень громкости звука, вызывающий болевое ощущение, равен 130-140 дБ над порогом слышимости человека. Громкие и длительные звуки (например, рок-музыка, рев реактивного двигателя) приводят к поражению рецепторных клеток и к снижению слуха. Адаптация. Если на ухо долго действует тот или иной звук, то чувствительность к нему падает. Степень этого снижения чувствительности (адаптации) зависит от длительности, силы звука и его частоты. Участие в слуховой адаптации нейронных механизмов типа латерального и возвратного торможения несомненно. Известно также, что сокращения мышц среднего уха могут изменять энергию сигнала, передающуюся на улитку. Бинауральный слух. Человек и животные обладают пространственным слухом, т. е. способностью определять положение источника звука в пространстве. Это свойство основано на наличии бинаурального слуха, или слушания двумя ушами. Острота бинаурального слуха у человека очень высока: положение источника звука определяется с точностью порядка 1 углового градуса. Основой этого служит способность нейронов слуховой системы оценивать различия времени прихода звука на правое и левое ухо и интенсивности звука на каждом ухе. Если источник звука находится в стороне от средней линии головы, то звуковая волна приходит на одно ухо несколько раньше и имеет большую силу, чем на другом ухе. Оценка удаленности источника звука от организма связана с ослаблением звука и изменением его тембра. При раздельной стимуляции правого и левого уха через наушники задержка между звуками уже в 11 мкс или различие в интенсивности двух звуков на 1 дБ приводят к кажущемуся сдвигу локализации источника звука от средней линии в сторону более раннего или более сильного звука. В слуховых центрах имеются нейроны с острой настройкой на определенный диапазон интерауральных различий по времени и интенсивности. Найдены также клетки, реагирующие лишь на определенное направление движения источника звука в пространстве.
Вестибулярная система
Вестибулярная система играет важную роль в пространственной ориентации человека. Она получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной системы не возбуждаются. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры, что обеспечивает сохранение равновесия тела.
Вестибулярного аппарата
Периферическим отделом вестибулярной системы является вестибулярный аппарат, расположенный в пирамиде височной кости. Он состоит из преддверия и трех полукружных каналов. Полукружные каналы (рис. 4.9) располагаются в трех взаимно перпендикулярных плоскостях. Один из концов каждого канала расширен (ампула). Вестибулярный аппарат включает в себя также два мешочка. В них на возвышениях находится отолитовый аппарат: скопления рецепторных клеток. Выступающая в полость мешочка рецепторная клетка оканчивается длинным подвижным волоском и 60-80 склеенными неподвижными волосками. Они пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция — отолиты. Волосковые клетки возбуждаются при скольжении отолитовой мембраны по волоскам, т. е. при их сгибании.
Рис. 4.9. Строение отолитового аппарата: 1 — отолиты; 2 — отолитовая мембрана; 3 — волоски рецепторных клеток; 4 — рецепторные клетки; 5 — опорные клетки; б — нервные волокна
Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, активируют нейроны бульбарного вестибулярного комплекса (ядра: преддвер-ное верхнее Бехтерева, преддверное латеральное Дейтер-са, Швальбе и др.). Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору мозга, ретикулярную формацию и вегетативные ганглии.
Популярное:
|
Последнее изменение этой страницы: 2016-03-26; Просмотров: 1416; Нарушение авторского права страницы

 Механизмы слуховой рецепции. При колебаниях основной мембраны длинные волоски рецепторных клеток касаются текторинальной мембраны и несколько наклоняются. Это приводит к натяжению тончайших нитей, которые открывают ионные каналы в мембране рецептора. Пресинапти-ческое окончание волосковой клетки деполяризуется, что приводит к выходу в синаптическую щель нейромедиа-тора (глутамата или аспартата). Воздействуя на постси-наптическую мембрану афферентного волокна, медиатор вызывает в нем генерацию возбуждающего постсинаптического потенциала и импульсов, которые распространяются в нервные центры.
Механизмы слуховой рецепции. При колебаниях основной мембраны длинные волоски рецепторных клеток касаются текторинальной мембраны и несколько наклоняются. Это приводит к натяжению тончайших нитей, которые открывают ионные каналы в мембране рецептора. Пресинапти-ческое окончание волосковой клетки деполяризуется, что приводит к выходу в синаптическую щель нейромедиа-тора (глутамата или аспартата). Воздействуя на постси-наптическую мембрану афферентного волокна, медиатор вызывает в нем генерацию возбуждающего постсинаптического потенциала и импульсов, которые распространяются в нервные центры. Передача в мозг акустической информации. Сигналы от волосковых клеток поступают в мозг по 32 000 афферентных нервных волокон, входящих в состав кохлеарной ветви восьмого черепно-мозгового нерва. Они являются дендритами ганглио-ных нервных клеток спирального ганглия. По волокнам слухового нерва даже в тишине следуют спонтанные импульсы с частотой до 100 имп./с. При звуковом раздражении частота импульсации в волокнах увеличивается и остается повышенной в течение всего периода, когда действует звук. Степень учащения разрядов различна у разных волокон и связана с интенсивностью и частотой звукового воздействия. В центральных отделах слуховой системы много нейронов, возбуждение которых длится в течение всего периода действия звука, а в слуховой коре разряды ряда нейронов длятся десятки секунд после его прекращения.
Передача в мозг акустической информации. Сигналы от волосковых клеток поступают в мозг по 32 000 афферентных нервных волокон, входящих в состав кохлеарной ветви восьмого черепно-мозгового нерва. Они являются дендритами ганглио-ных нервных клеток спирального ганглия. По волокнам слухового нерва даже в тишине следуют спонтанные импульсы с частотой до 100 имп./с. При звуковом раздражении частота импульсации в волокнах увеличивается и остается повышенной в течение всего периода, когда действует звук. Степень учащения разрядов различна у разных волокон и связана с интенсивностью и частотой звукового воздействия. В центральных отделах слуховой системы много нейронов, возбуждение которых длится в течение всего периода действия звука, а в слуховой коре разряды ряда нейронов длятся десятки секунд после его прекращения. Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютным порогом слуховой чувствительности. Пороги слышимости сильно зависят от частоты звука. В области частот от 1000 до 4000 Гц слух человека максимально чувствителен. В этих пределах слышен звук, имеющий ничтожную энергию. При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука в 1 млн раз выше (нижняя кривая AEFGD на рис. 4.8). При усилении звука можно дойти до возникновения неприятного ощущения давления и даже боли в ухе.1 Звуки такой силы характеризуют верхний предел слышимости (кривая ABCD на рис. 4.8) и ограничивают область нормального слухового восприятия. Внутри этой области лежат и так называемые речевые поля, в пределах которых распределяются звуки речи.
Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютным порогом слуховой чувствительности. Пороги слышимости сильно зависят от частоты звука. В области частот от 1000 до 4000 Гц слух человека максимально чувствителен. В этих пределах слышен звук, имеющий ничтожную энергию. При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука в 1 млн раз выше (нижняя кривая AEFGD на рис. 4.8). При усилении звука можно дойти до возникновения неприятного ощущения давления и даже боли в ухе.1 Звуки такой силы характеризуют верхний предел слышимости (кривая ABCD на рис. 4.8) и ограничивают область нормального слухового восприятия. Внутри этой области лежат и так называемые речевые поля, в пределах которых распределяются звуки речи.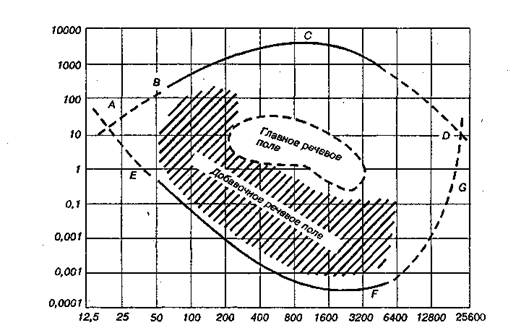
 4.4.1. Строение и функции рецепторного
4.4.1. Строение и функции рецепторного 
 В перепончатых полукружных каналах, заполненных эндолимфой, рецепторные волосковые клетки сконцентрированы в ампулах. Во время угловых ускорений эн-долимфа приходит в движение, волоски сгибаются и волосковые клетки возбуждаются. При противоположно направленном движении они тормозятся. Это связано с тем, что отклонение волоска в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении закрывает каналы и гиперполяризует рецептор. В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.
В перепончатых полукружных каналах, заполненных эндолимфой, рецепторные волосковые клетки сконцентрированы в ампулах. Во время угловых ускорений эн-долимфа приходит в движение, волоски сгибаются и волосковые клетки возбуждаются. При противоположно направленном движении они тормозятся. Это связано с тем, что отклонение волоска в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении закрывает каналы и гиперполяризует рецептор. В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.