|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Кислородсодержащие производные 5-метилцитозина.
В результате последовательного окисления 5-метилцитозина ферментами семейства ТЕТ образуются кислородсодержащие производные – 5-гидроксиметилцитозин, 5-формилцитозин и 5-карбоксилцитозин. Эти 3 соединения были обнаружены в ДНК млекопитающих (Ito et al., 2011; Kriaucionis, Heintz, 2009; Tahiliani et al., 2009). В научной литературе появляется все больше сведений о собственных функциях кислородсодержащих производных 5-метилцитозина. 5-гидроксиметилцитозин был впервые обнаружен в геноме Т-четного бактериофага, где использовался для того, чтобы уходить от распознавания бактериальными рестриктазами (Wyatt, Cohen, 1953). В начале 1970-х годов он был найден в ДНК, выделенной из мозга крыс и мышей (Penn et al., 1972). Однако в течение последующих 30 лет этому открытию не уделялось должного внимания. В настоящее время описано наличие 5-гидроксиметилциозина в эмбриональных тканях, стволовых клетках, а также в мужских пронуклеусах оплодотворенных яйцеклеток мышей (Iqbal et al., 2011; Wossidlo et al., 2011). Хотя и известно, что уровень 5-гидроксиметилцитозина в эмбриональных стволовых клетках снижается во время дифференциации, до сих пор неизвестна причина изменения уровня 5-гидроксиметилцитозина в тканях взрослых организмов. Также как и 5-метилцитозин, 5-гидроксиметилцитозин находится после сайтов начала транскрипции, однако его обнаружили и во внутригенных областях (Pastor et al., 2011). Недавно полученные данные показали, что 5-гидроксиметилцитозин является стабильной эпигенетической модификацией, играющей свою роль в эпигенетической регуляции генома. Было показано, что 5-гидроксиметилцитозин снижает температуру плавления ДНК по сравнению с метилированной (Wanunu et al., 2011). Больше всего обогащены 5-гидроксиметилцитозином промоторные участки генов, что указывает на связь с регуляцией транскрипции (Pastor et al., 2011). Некоторые ДНК связывающие белки, такие как MeCP2 различают 5-метилцитозин и 5-гидроксиметилцитозин, в то время как другие белки, например, фактор поддерживающей метилтрансферазы Uhrf1, связывает оба модифицированных цитозина (Frauer et al., 2011). Кроме того, превращение 5-метилцитозина в 5-гидроксиметилцитозин способствует пассивному деметилированию ДНК, поскольку поддерживающая метилтрансфераза DNMT1 плохо узнаёт 5-гидроксиметилцитозин и после репликации не метилирует новую синтезированную нить (Valinluck, Sowers, 2007). Последующее окисление 5-гидроксиметилцитозина ферментами ТЕТ приводит к образованию следующих промежуточных продуктов активного деметилирования – 5-формилцитозина (5-fC) и 5-карбоксилцитозина (5-caC) (Ito et al., 2011). Высказываются предположения о том, что 5-fC и 5-caC также являются стабильными эпигенетическими модификациями и могут выполнять определенные функции. Биофизическими методами было выявлено, что присутствие 5-формилцитозина в гуанин-цитозиновых динуклеотидах приводит к изменению физических свойств двойной цепи ДНК. Сама молекула ДНК приобретает особый вид, F-конформацию, для которой характерно ослабленное закручивание спирали. По этой причине наличие 5-формилцитозина может влиять на взаимодействие с гистонами и структуру хроматина. Сильное формилирование гуанин-цитозиновых динуклеотидов, запускаемое ферментами семейства ТЕТ, может контролировать экспрессию генов путем изменения физических свойств ДНК (Raiber et al., 2015). Более того, удаление 5-формилцитозина или восстановление до 5-гидроксиметилцитозина вызывает обратное превращение ДНК в В-конформацию. Таким образом, физические свойства молекулы ДНК могут динамически изменяться под действием гликозилаз (удаляющих 5-fC) и ферментов семейства ТЕТ, окисляющих 5-гидроксиметилцитозин до 5-формилцитозина, влияя на экспрессию генов (Raiber et al., 2015). С помощью полнопротеомного анализа в эмбриональных стволовых клетках мыши были обнаружены белки, которые могут связывать или считывать 5-формилцитозин и 5-карбоксилцитозин. В числе таких белков гликозилазы, регуляторы транскрипции и структуры хроматина (Iurlaro et al., 2013; Spruijt et al., 2013). Формильная группа является химически более активной относительно карбоксильной и способна взаимодействовать с различными компонентами клетки (Song, He, 2013). Кроме того, 5-формилцитозин и 5-карбоксилцитозин, как показано, снижают скорость транскрипции и субстратную специфичность РНК полимеразы II (Kellinger et al., 2012). А накопление 5-fC/5-caC на дистальных регуляторных элементах в отсутствие TDG согласовано со связыванием транскрипционного коактиватора р300 (Song et al., 2013). р300 может служить мостом для транскрипционных факторов и всего аппарата транскрипции, способствуя, тем самым ремоделированию хроматина и активации транскрипции генов (Chen, Li, 2011). Кислородсодержащие формы 5-метилцитозина появляются не только ферментативным путем, но и в результате окислительного стресса. 5-mC может подвергаться реакциям окисления, опосредованным эндогенными и экзогенными формами активного кислорода, такими как гидроксильный анион-радикал (HO•− ), пероксид водорода (H2O2), супероксидный анион-радикал (O2•− ) и одноэлектронные окислители (Cadet, Wagner, 2013; Cadet et al., 2012). Облучение дальним ультрафиолетом или ионизирующим излучением 5-метилцитозина также вызывает формирование кислородсодержащих производных цитозина, главным образом, через образование активных форм кислорода в водном растворе, содержащем ДНК (Cadet, Wagner, 2013). Однако эффективность радикальных реакций и фотоионизации падает для каждого последующего окисления, поэтому образование 5-карбоксилцитозина в результате окислительного стресса маловероятно. Таким образом, кислородсодержащие формы 5-метилцитозина возможно являются не только промежуточными продуктами активного деметилирования, но и выполняют определенные функции в эпигенетической регуляции генома.
1.2. Основные этапы женского гаметогенеза и доимплантационного развития человека Онтогенез, или индивидуальное развитие, человека включает пренатальный (внутриутробный) период, который длится примерно 280 суток, и постнатальный (внеутробный) период, длительность которого варьирует у различных людей в зависимости от внутренних и внешних факторов. Пренатальный период начинается с момента оплодотворения яйцеклетки сперматозоидом и заканчивается рождением ребенка (Баранов, Кузнецова, 2007). Оплодотворению предшествует процесс формирования половых клеток - гаметогенез. В процессе гаметогенеза образуются яйцеклетки с одинаковым набором хромосом - 22 аутосомы и одна половую Х-хромосому. В результате мужского гаметогенеза образуются сперматозоиды с отличающимися наборами хромосом - 22 + Х и 22 + Y (Баранов, Кузнецова, 2007). Женский гаметогенез представляет собой сложный и многостадийный процесс, который занимает значительную часть онтогенеза, и чьи этапы значительно разнесены во времени. Женский гаметогенез начинается с того, что примордиальные (первичные) половые клетки заселяют гонады и приступают к делениям митозом. При митотических делениях образуются генетически одинаковые диплоидные клетки, называемые оогониями. Спустя некоторое количество делений, число оогониев достигает нескольких миллионов. Женский гаметогенез на этом этапе протекает во время внутриутробного развития (2-5 месяц) в эмбриональном яичнике. В этот период число половых клеток увеличивается с нескольких тысяч до 7 миллионов. Однако вскоре огромное число оогониев претерпевает естественную гибель в результате процесса, названного атрезией. Атрезия постоянно происходит в женском яичнике вплоть до наступления менопаузы. В результате, только 40 тысяч оогониев смогут вступить в мейоз, а из них лишь 400 дадут зрелые яйцеклетки, способные к оплодотворению (Carlson, 2009) (рис. 2 А ). Незадолго до рождения оогонии входят в мейоз и становятся первичными ооцитами. В первые месяцы после рождения ооцит входит в диплотену первого деления мейоза и наступает первый блок оогенеза. В этот период происходит накопление белков, рибосомных и других типов РНК. Это необходимо будущей зрелой яйцеклетке и зиготе, которая должна будет быстро совершить ряд делений дробления, для чего потребуется большое количество запасенной энергии и строительного материала. Также происходит накопление кортикальных гранул, главным образом за счет аппарата Гольджи, которые во время оплодотворения защитят яйцеклетку от полиспермии. С началом мейоза клетки яичника постепенно окружают первичные ооциты, формируя, таким образом, первичные фолликулы (рис. 2 А ). Ооциты и фолликулярные клетки образуют между собой множество микроворсинок и щелевых контактов, позволяющих обмениваться аминокислотами и глюкозой, необходимыми для роста ооцита. Также фолликулярные клетки секретируют фактор, подавляющий мейоз и поддерживающий блок на стадии диплотены. Первый блок оогенеза длится вплоть до полового созревания. За это время первичные фолликулы увеличиваются в размере, главным образом за счет роста самих ооцитов, а также увеличения числа фолликулярных клеток, после чего становятся вторичными фолликулами (рис. 2 А ). Также между ооцитом и фолликулярными клетками происходит формирование неклеточной мембраны, называемой блестящим слоем. Блестящий слой содержит рецепторы к сперматозоидам и другие компоненты, играющие важную роль в оплодотворении (Carlson, 2009). С начала полового созревания под действием фолликул-стимулирующего гормона (ФСГ) в фолликуле происходит образование полости (антрума), наполненной фолликулярной жидкостью. С образованием полости фолликулярные клетки разделяются на 2 группы: прилегающие непосредственно к ооциту клетки кумулюса, и находящиеся между полостью и фолликулярной мембраной пристеночные фолликулярные клетки (рис. 2 А ). В ответ на гормональное воздействие фолликул резко увеличивается в размере и давит на поверхность яичника. В таком состоянии фолликул называется третичным, или граафовым пузырьком. За 10-12 часов до овуляции возобновляется мейоз. Во время каждого менструального цикла от 10 до 30 первичных ооцитов завершают первое деление мейоза и начинают дальнейшее развитие, при этом остальные будут ждать своей очереди еще долгие годы. После первого деления мейоза образуется 2 неравные клетки – большой вторичный ооцит и малое первичное полярное тельце. Тогда же третичный фолликул начинает выпячивать поверхность яичника и еще больше увеличивается в размере, главным образом за счет протеогликанов и гиалуроновой кислоты. Теперь фолликул готов к овуляции и ожидает стимула в виде ФСГ и лютеинизирующего гормона (ЛГ), выделяемых передней долей гипофиза (Carlson, 2009). Вторичный ооцит, вступивший во второе деление мейоза, претерпевает блок оогенеза на стадии метафазы, снятие которого осуществляется только после оплодотворения сперматозоидом. Если оплодотворение не произошло, то вторичный ооцит не может завершить мейоз и подвергается атрезии. Если оплодотворение прошло успешно, ооцит завершает второе мейотическое деление, в результате которого образуется вторичное полярное тельце, а зигота приступает к формированию отцовского и материнского пронуклеусов и готовится к их слиянию (Carlson, 2009). Собственно процесс оплодотворения условно разделяют на три фазы – дистантного взаимодействия, контактного взаимодействия и фазу объединения генетического материала половых клеток (Данилов, Боровая, 2003). Во время фазы дистантного взаимодействия происходит обмен сигналами между половыми клетками в половых путях женщины. Важными механизмами дистантного взаимодействия являются положительные хемо- и реотаксис, чувствительность к изменениям температуры, а уже на близком расстоянии - электростатическое взаимодействие гамет (Bahat, Eisenbach, 2006). В фазе контактного взаимодействия сперматозоид начинает разрушать оболочки ооцита – сначала лучистый венец (corona radiata), затем блестящий слой (zona pellucida) и в конце - цитоплазматическую мембрану. На основе этого фазу контактного взаимодействия можно условно разделить на 3 этапа: прохождение лучистого венца, прохождение прозрачной зоны, слияние мембран ооцита и сперматозоида (Данилов, Боровая, 2003) (рис. 2 Б ). Таким образом, из многих миллионов сперматозоидов только 300-500 смогут достичь места оплодотворения. Из них лишь 1 оплодотворяет яйцеклетку, в том время как остальные помогают в прохождении барьеров, защищающих женскую половую клетку (Данилов, Боровая, 2003).
Рис 2. Объединенная схема: А).женского гаметогенеза; Б).оплодотворения яйцеклетки, формирования и слияния разнородительских пронуклеусов на стадии зиготы. (по Cаrlson Human embryology and developmental biology, 4th edition, 2009)
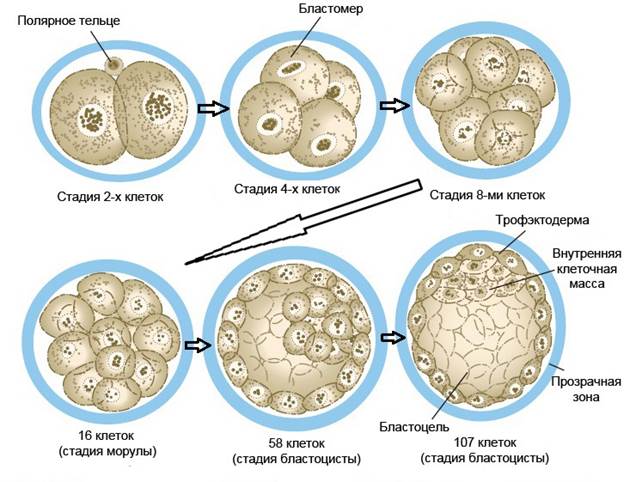
Доимплантационное развитие человека длится около 6–7 дней и включает морфогенетические процессы, такие как формирование пронуклеусов, дробление, компактизация и бластуляция. Во время фазы объединения генетического материала родительских половых клеток появляется диплоидная зигота. Однако сначала яйцеклетка должна завершить второе деление мейоза. Тем временем пронуклеус сперматозоида округляется и принимает вид интерфазного. В нем происходит репликация ДНК, в результате чего мужской пронуклеус имеет гаплоидный набор реплицированных хромосом. Женский пронуклеус после завершения мейоза претерпевает точно такие же изменения. Затем оба пронуклеуса переходят в профазу митоза. Центриоль, привнесенная сперматозидом, делится с образованием двух центросом. Они прикрепляются к веретену, образующемуся между мужским и женским наборами хромосом, что, таким образом, приводит к их совместному расположению в экваториальной плоскости – так осуществляется метафаза митоза зиготы. Далее следует ана- и телофаза – зигота завершает первое деление дробления, в результате чего формируются две дочерние клетки с диплоидным набором хромосом, называемые бластомерами (Данилов, Боровая, 2003). Дробление представляет собой ряд митотических делений оплодотворенной яйцеклетки, в результате которых происходит увеличение числа клеток при неизменной массе (рис.3). Для доимплантационного периода развития человека характерно полное неравномерное дробление, при котором бластомеры дробятся с различной скоростью, а также могут существенно различаться по размерам (Баранов, Кузнецова, 2007). Первое деление дробления длится в среднем около 30 часов. Последующие деления происходят примерно каждые 18 часов с образованием все более мелких бластомеров. Начиная с 2-клеточной стадии, возникает асинхронность ритма дробления бластомеров, что приводит к формированию зародышей с нечетным количеством бластомеров. В течение трех первых делений дробления все бластомеры имеют округлую форму, идентичны по своей морфологии и способностям к росту (Баранов, Кузнецова, 2007). Зародыш, состоящий из 8-16 бластомеров, называют морулой (от латинского слова morum, означающего ягоду тутового дерева). На этой стадии (3–5-й дни развития) начинается процесс компактизации, сопровождающийся поляризацией бластомеров. При этом бластомеры, расположенные снаружи, приобретают свойства эпителиальных клеток, между ними возникают плотные межклеточные контакты (десмосомы), а внутри зародыша начинает накапливаться жидкость (Баранов, Кузнецова, 2007; Carlson, 2009). Этот процесс был назван кавитацией, а полость, наполненная жидкостью – бластоцелем. Таким образом, на 5 день после оплодотворения морула превращается в бластоцисту. Бластоциста, окруженная блестящим слоем (zona pellucida), состоит из двух типов клеток: наружного слоя (трофэктодермы) и внутренней небольшой группы клеток эмбриобласта (внутренняя клеточная масса — ВКМ). Общее число клеток на этой стадии — более 32 (Carlson, 2009).
Рис.3. Первые стадии дробления зародыша человека.
Клетки трофэктодермы начинают активно синтезировать хорионический гонадотропин и различные лизирующие ферменты, с помощью которых приобретают инвазивные свойства, обеспечивая гистиотрофное питание зародыша и процесс имплантации. После имплантации клетки трофэктодермы образуют только экстраэмбриональные ткани, в которые также входит наружная часть плаценты (Баранов, Кузнецова, 2007; Carlson, 2009). Клетки ВКМ активно делятся и в дальнейшем развитии дадут начало всем тканям собственно эмбриона, а также его внезародышевым частям (хорион, внутренняя часть плаценты, желточный мешок, аллантоис, амнион). В ходе делений дробления зародыш продвигается по маточной трубе в направлении матки как за счет тока жидкости, так и вследствие перистальтических сокращений мускулатуры и движения эпителиальных ресничек. На 5–6-й день зародыш на стадии бластоцисты попадает в матку (Баранов, Кузнецова, 2007; Carlson, 2009). Первые деления дробления осуществляются за счет генетической информации, накопленной ооцитом еще в период оогенеза. Зародыш человека обладает достаточным запасом готовых белков, рибонуклеопротеиновых комплексов, необходимых для синтеза новых белков, а также питательных веществ и энергетических ресурсов, чтобы полностью обеспечить начальные этапы эмбриогенеза. Синтез белков, необходимых на этой стадии, происходит на матрицах РНК, синтезированных еще в оогенезе, то есть на хромосомах ооцита. Отсюда ее название — материнская РНК (Баранов, Кузнецова, 2007; Carlson, 2009). Переход к синтезу собственных мРНК зародыша (или активация генома зародыша, от англ. EGA - embryonic genome activation) представляет собой запуск транскрипции эмбрионального генома и уменьшение количества материнских мРНК. В инактивации эмбрионального генома во время первых делений дробления участвуют по крайней мере 3 механизма: хроматин-зависимая инактивация; нехватка ферментов, обеспечивающих транскрипцию, в том числе, ТАТА-связывающих белков; быстрое прохождение клеточного цикла на первых делениях дробления, в результате чего частая репликация ДНК препятствует транскрипции (Schier, 2007). В исследованиях, проведенных на млекопитающих, было обнаружено, что это EGA является видоспецифичным процессом и сроки его наступления варьируют в зародышах разных видов. Так, у коров переход на синтез мРНК зародыша происходит на стадии 8 клеток, у овец на стадии 8-16 клеток, у свиней на стадии 4 клеток и на стадии 2 клеток у мышей (Memili, First, 2000). В раннем эмбриогенезе человека выделяют 2 этапа включения генома зародыша: раннее EGA и основное EGA. Первому этапу соответствует период от стадии зиготы до 4-клеточного зародыша, во время которого происходит значительное сокращение материнских м-РНК, но и в то же время появление нескольких транскриптов зародыша, включающих в основном 5`-нетранслируемые участки кодирующих генов. Во время основного EGA происходит вторая волна активации генома, что проявляется в появлении большого числа транскриптов зародыша, в 75% из которых - кодирующие гены белков, обладающих каталитической и ДНК-связывающей активностью. Большинство исследователей сходятся во мнении, что у человека основное включение генома зародыша происходит на стадии 4-8 клеток, однако есть серьезные расхождения в оценке обширности этого процесса: от включения 129 (Toehoenen et al., 2015) генов зародыша до 2495 (Yan et al., 2013). Toehoenen сотоварищи объясняют столь небольшое число обнаруженных активированных генов в ходе EGA тем, что работа многих биологических процессов регулируется каскадами и поэтому достаточно лишь небольшого числа ключевых генов для нормального прохождения активации генома и дальнейшего эмбриогенеза (Toehoenen et al., 2015). Активация генома зародыша – сложный и многоэтапный процесс, изучение которого является непростой задачей, в том числе и из-за отсутствия данных об эпигенетических аспектах EGA.
Популярное:
|
Последнее изменение этой страницы: 2016-04-09; Просмотров: 1232; Нарушение авторского права страницы