|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
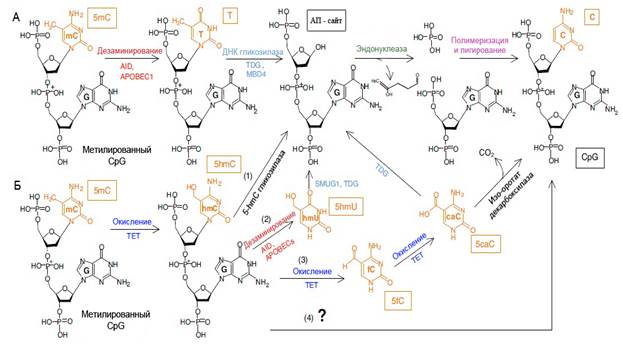
Механизмы деметилирования ДНК
Удаление метильных групп ДНК может осуществляться двумя способами: пассивно и активно. Пассивное деметилирование происходит за счет отсутствия метилазной активности после репликации ДНК. В данном случае новая нить не метилируется по образцу старой, и образуется так называемая гемиметилированная ДНК. Последующая репликация при отсутствии метилазной активности приводит к образованию уже полностью деметилированной ДНК. Активное деметилирование осуществляется с помощью особых ферментов и проходит независимо от репликации. В последнее время были открыты несколько ключевых ферментов, участвующих в модификации или удалении 5-метилцитозина – AID/APOBEC дезаминазы, ТЕТ оксидазы, ДНК-гликозилазы TDG и SMUG (Saitou et al., 2012). Дезаминирование – реакция удаления аминогруппы. В результате дезаминирования цитозин превращается в урацил, 5-метилцитозин – в тимин, а 5-гидроксиметилцитозин – в 5-гидроксиметилурацил. AID и APOBEC являются цитидиновыми дезаминазами позвоночных и могут вызывать мутации в ДНК и РНК путем дезаминирования цитидина в уридин (Saitou et al., 2012). Гены AID и APOBEC1 экспрессируются в ооцитах и первичных половых клетках мышей, что свидетельствует об участии этих ферментов в глобальном деметилировании (Bhutani et al., 2010). Недавно было идентифицировано несколько белков семейства ТЕТ (Ten-Eleven Translocation) - TET1, TET2 и TET3 – ключевых ферментов, вовлеченных в окисление метильной группы 5-метилцитозина в клетках млекопитающих (Tahiliani et al., 2009; Ito et al., 2011). Важно отметить, что оксидазы семейства ТЕТ локализуются в разных типах клеток и на разных этапах онтогенеза. ТЕТ1 впервые был обнаружен в клетках Пуркинье головного мозга, а затем и в эмбриональных стволовых клетках мыши (Kriaucionis, Heintz, 2009; Tahiliani et al., 2009). TET2 специфичен для клеток гемопоэтического ряда и нервных клеток в постнатальный период, а хромосомные делеции участка, содержащего ТЕТ2, были выявлены при миелопролиферативных заболеваниях, что предполагает наличие у этого фермента функции супрессора опухолей (Tahiliani et al., 2009). TET3 накапливается в ооцитах и оплодотворенных яйцеклетках, причем его транскрипты быстро деградируют после оплодотворения, и практически полностью удаляются уже на стадии двухклеточного зародыша (Iqbal et al, 2011). Все 3 белка ТЕТ имеют CD домен, а также обладают α -кетоглутарат и Fe(II)-зависимой диоксигеназной активностью. Однако для осуществления окисления метилированной ДНК им необходимы двухатомный кислород (О2), АТФ и соли аскорбиновой кислоты в качестве кофакторов (Ko et al., 2013). Помимо участия в активном деметилировании, была обнаружена связь между активностью ферментов семейства ТЕТ и длиной теломер в эмбриональных стволовых клетках мышей. В недавнем исследовании было выявлено, что в клетках с полным отсутствием ТЕТ1, ТЕТ2 и ТЕТ3 происходит увеличение длины теломер, а также активация экспрессии кластера генов Zscan4 (Lu et al., 2014). Активация экспрессии Zscan4, в свою очередь, повышает частоту рекомбинации между участками теломерных повторов, что приводит к удлинению теломер (Zalzman et al., 2010). ДНК-гликозилазы – ферменты, которые распознают и удаляют поврежденные или некомплементарные основания с образованием апуриновых/апиримидиновых сайтов. Далее запускается механизм эксцизионной репарации оснований (BER – base excision repair), который устраняет небольшие (не искажающие спираль ДНК) пробелы в геноме. Эксцизионная репарация состоит из следующих этапов: сначала эндонуклеазы удаляют апуриновые/апиримидиновые сайты с образованием однонитевого разрыва, затем ДНК-полимераза β заделывает разрыв, а ДНК-лигаза сшивает вновь синтезированный участок со старой нитью. ДНК-гликозилазы бывают 2-х типов – монофункциональные и бифункциональные. Монофункциональные гликозилазы обладают только гликозилазной активностью, тогда как бифункциональные помимо этого выполняют функцию эндонуклеаз и могут удалять апуриновые/апиримидиновые сайты (Saitou et al., 2012). Одним из главных представителей семейства ДНК-гликозилаз, участвующих в деметилировании, является тиминовая ДНК-гликозилаза (от английского TDG – thymine DNA glygosylase). TDG удаляет тимин из некомплементарных пар гуанин-тимин, цитозин-тимин и тимин-тимин, тем самым защищая геном от мутаций, вызванных спонтанным дезаминированием 5-метилцитозина в тимин. Тиминовая гликозилаза также специфично вырезает 5-формилцитозин (5-fC) и 5-карбоксилцитозин (5-caC), хотя и не имеет такой активности по отношению к 5-гидроксиметилцитозину (Maiti, Drohat, 2011). Как оказалось, прочность связи между основаниями не является ключевым фактором специфичности TDG, а как раз наоборот, она предпочтительнее разрезает более прочную связь гуанин-5-формилцитозин, нежели менее прочную гуанин-тимин (Darwanto et al., 2009). Известно, что 5-формилцитозин может образовываться не только при помощи оксидаз семейства ТЕТ, но и при воздействии ультрафиолета, ионизирующего излучения или активных форм кислорода на 5-метилцитозин. Было показано, что 5-формилцитозин обладает мутагенной активностью, а также приводит к снижению скорости репликации ДНК (Kamiya et al., 2002). Таким образом, фермент TDG может не только участвовать в процессе активного деметилирования, удаляя 5-fC и 5-caC, но и защищать геном от мутагенной активности 5-формилцитозина. На основе полученных данных по функциям, свойствам и взаимодействиям вышеописанных ферментов были сформулированы 3 основные модели активного деметилирования: дезаминазный путь (Fritz, Papavasiliou, 2010), оксидазный путь (Wu, Zhang, 2010) и модель, сочетающая оба пути (Saitou et al., 2012). Дезаминазный путь активного деметилирования характеризуется превращением 5-метилцитозина в тимин путем дезаминирования ферментами семейства AID/APOBEC. Затем ферменты эксцизионной репарации распознают неправильные пары тимина с гуанином и вырезают тимин с последующей вставкой немодифицированного цитозина (Fritz, Papavasiliou, 2010) (рис. 1. А ). Однако дезаминазы преимущественно действуют на однонитевую ДНК и в окружении определенных последовательностей, а большинство метилированных локусов располагается именно в двунитевой ДНК в окружении CpG динуклеотидов. Вдобавок, активность дезаминаз к 5-метилцитозину снижена по сравнению с немодифицированным цитозином (Bransteitter et al., 2003). Поэтому дезаминазный путь не является основным в активном деметилировании. Согласно оксидазной модели активного деметилирования три белка семейства TET (Ten-Eleven-Translocation) – TET1, TET2 и TET3 – последовательно превращают 5-метилцитозин сначала в 5-гидроксиметилцитозин, затем в 5-формилцитозин, и, наконец, в 5-карбоксилцитозин (Ito et al., 2010). Все 4 типа модифицированных форм цитозина – 5-метилцитозин, 5-гидроксиметилцитозин, 5-формилцитозин и 5-карбоксилцитозин – присутствуют в геномной ДНК в разном количестве, находящемся в зависимости от уровня экспрессии ТЕТ (He et al., 2011; Ito et al., 2011). Подтверждает участие ферментов ТЕТ в активном деметилировании еще и то, что они преимущественно связываются с CpG динуклеотидами, метилирование в которых наиболее значимо (Ito et al., 2010). Для полноценного завершения процесса активного деметилирования необходимо удаление кислородсодержащих форм 5-метилцитозина. Оно может проходить несколькими путями. 5-гидроксиметилцитозин может быть дезаминирован ферментами AID/APOBEC с образованием 5-гидроксиметилурацила, который затем удаляется гликозилазами SMUG или TDG (Guo et al., 2011) (рис.1.Б (2)). Однако способность ферментов AID/APOBEC эффективно дезаминировать 5-гидроксиметилцитозин пока не подтверждена. Также у млекопитающих не был обнаружен 5-гидроксиметилураци в достаточных для детекции количествах. Поэтому такой вариант оксидазного пути активного деметилирования пока не подтвержден. Более вероятным рассматривается участие фермента эксцизионной репарации TDG, который распознает и эффективно вырезает 5-формилцитозин и 5-карбоксилцитозин, не затрагивая при этом 5-гидроксиметилцитозин (Maiti, Drohat, 2011) (рис.1.Б (3)). На основании этих данных была предположена общая модель, в которой ферменты ТЕТ являются основными регуляторами деметилирования ДНК, а оксидазный путь удаления метильных групп, связанный с эксцизионной репарацией, является основным путем деметилирования. При этом дезаминазы играют важную вспомогательную роль в ускорении этого процесса. В процессе дезаминирования могут появляться модифицированные формы урацила, которые вырезаются ферментами эксцизионной репарации эффективнее, чем модифицированные формы цитозина (Saitou et al., 2012). Таким образом, модель, включающая основной и дополнительные пути, объясняет участие как ферментов эксцизионной репарации, так и оксидаз ТЕТ с дезаминазами AID/APOBEC в активном деметилировании, необходимом для регуляции эмбриогенеза, правильного развития и дифференцировки стволовых клеток.
Рис.1. Возможные пути активного деметилирования.(поSaitou et al., 2012) Примечания: (А) Дезаминирование цитозина ферментами AID/APOBEC (красным) и последующая эксцизионная репарация BER с участием ферментов TDG/MBD4 (голубым). (Б) Гидроксилирование 5-метилцитозина ферментом TET приводит к 4 вариантам путей возможного деметилирования: (1) удаление 5-гидроксиметилцитозина (5-hmC) ещё не обнаруженной гликозилазой с переходом на путь эксцизионной репарации; (2) дезаминирование 5-гидроксиметилцитозина ферментами AID/APOBEC производит 5-гидроксиметилурацил (5-hmU), который удаляется ферментом SMUG1 или TDG; (3) окисление 5-hmC до 5-формилцитозина (5-fC) и 5-карбоксилцитозина (5-caC), который может быть превращен в цитозин декарбоксилазой, либо удален ферментом TDG с переходом на путь эксцизионной репарации; (4) прямое превращение 5-hmC в цитозин. Сокращения: CpG – цитозин-гуаниновый динуклеотид, 5-mC – 5-метилцитозин, 5-hmC – 5-гидроксиметилцитозин, 5-hmU – 5-гидроксиметилурацил, 5-fC – 5-формилцитозин, 5-caC – 5-карбоксилцитозин, АП сайт – апуриновый/апиримидиновый сайт, Т – тимин, С – цитозин.
Активное деметилирование ДНК является сложным и многостадийным процессом, в котором принимают участие ферменты различных семейств. Однако до сих пор стоит ряд вопросов, касающихся особенностей этого процесса на разных этапах онтогенеза. Также остается неясной регуляция активного деметилирования.
Популярное:
|
Последнее изменение этой страницы: 2016-04-09; Просмотров: 1957; Нарушение авторского права страницы