
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Продольная организация хромосом
Продольная организация хромосом высших организмов, в основе которой лежит взаимосвязь морфологических, химических и функциональных закономерностей, характеризуется линейной неоднородностью. Уже интерфазные хромосомы оказываются глубоко дифференцированными по степени конденсации хроматина, что первоначально было обнаружено с помощью световой микроскопии. Одни их участки становятся деконденсированными (эухроматин), другие остаются конденсированными (гетерохроматин). В метафазных хромосомах подразделение на эти два типа хроматина не исчезает. Оно проявляется в естественном ходе митотической конденсации: в ранней профазе районы гетерохроматина опережают в своей конденсации эухроматиновые участки. Остаточные явления неодинаковой конденсации интерфазной хромосомы обнаруживаются морфологически и в метафазе (в области вторичной перетяжки). Понятие " гетерохроматин" и " эухроматин" в результате цито - генетических исследований получили генетическое содержание. Гетерохроматин в отличие от эухроматина не содержит структурных генов или обеднён ими. В то же время эухроматин - это функционально активный, транскрибируемый хроматин, т.е. структура хроматина оказывает влияние на регуляцию экспрессии эукариотических генов. Подобно митотическому хроматину гетерохроматин не участвует в транскрипции, ДНК в составе гетерохроматина реплицируется в позднем периоде S-периода клеточного цикла. Биохимические основы наблюдаемых различий между гетеро- и эухроматином неизвестны. Некоторые участки хромосом конденсируются в гетерохроматин во всех клетках организма - это конститутивный гетерохроматин. Другие участки хромосом формируют гетерохроматин лишь в определенных клетках - факультативный гетерохроматин. Конститутивный гетерохроматин содержит ДНК, которая, по всей видимости, никогда ни в одной клетке не транскрибируется. В хромосомах человека он локализуется вокруг центромер и легко обнаруживается в митотических хромосомах с помощью специальной окраски, хотя может быть обнаружен и в других районах некоторых хромосом (1, 9, 16, У). Подобное же состояние характерно для сателлитной ДНК и ДНК с высокоповторяюшимися последовательностями. Следовательно, большая часть конститутивного гетерохроматина содержит серии сравнительно простых, многократно повторяющихся последовательностей ДНК. В целом функция конститутивного гетерохроматина остается неясной. Предполагают, что некоторые сегменты этого хроматина играют определенную роль в спаривании хромосом в мейозе. Возможно, он влияет на стабилизацию структуры хроматина и защищает генетически значимые последовательности эухроматических районов от внешних воздействий, но классических менделевских генов здесь, скорее всего, нет. В интерфазной клетке участки конститутивного хроматина агрегируют с образованием хромоцентров, что мы видим в световой микроскоп в виде мельчайших " глыбок хроматина". У млекопитающих количество их и характер распределения варьирует в зависимости от типа клетки и стадии развития организма. Факультативный гетерохроматин имеет более отчетливое функциональное значение. Почти не вызывает сомнения, что он отражает устойчивые различия в характере генетической активности клеток разных типов, и количество этого хроматина в разных клетках варьирует: в эмбриональных клетках его совсем немного, тогда как высокоспециализированные клетки содержат его в чрезвычайно больших количествах, т.е. часть генов выключается из транскрипции. Факультативный гетерохроматин содержит уникальные участки ДНК, а не высокоповторяющиеся, и ничем не обнаруживает себя при окрашивании митотических хромосом. Подобный способ генетической регуляции бактериям недоступен. Частный случай факультативной гетерохроматизации - это инактивация одной из двух Х-хромосом в клетках женских особей млекопитающих, которая происходит на ранних стадиях эмбрионального развития (в трофобласте человека на 12-й день развития, а собственно в эмбрионе на 16-й день). Одновременно во всех клетках эмбриона женской особи с равной вероятностью одна или другая Х-хромосома конденсируется и образует гетерохроматин. Это состояние хромосомы устойчиво наследуется во всех последующих циклах репликации. Из-за этого каждый женский организм имеет как бы мозаичное строение, т.к. образован клональными группами клеток, примерно в половине которых гетерохроматизирована Х-хромосома, унаследованная по материнской линии, а в другой - Х-хромосома, унаследованная по отцовской линии. В интерфазе гетерохроматизированные Х-хромосомы представляют собой четко оформленные структурные образования, называемые тельцами Барра, которые близко прилегают к внутренней мембране ядра и хорошо различимы в световой микроскоп. Тельца Барра называются ещё глыбками полового Х-хроматина.
***Отличительной особенностью интерфазной хромосомы от митотической, кроме всего прочего, является то, что по своей длине она может быть деконденсирована, развернута неравномерно: наряду с участками полной деконденсации есть участки, находящиеся в плотном, конденсированном и, соответственно, в неактивном состоянии. Это и придает интерфазному ядру своеобразную структуру, где хромосомы — хроматин — могут быть представлены то плотными блоками, то рыхлыми деконденсированными участками. 15. Политенные хромосомы. Эндомитоз.
Полите́ нные хромосо́ мы — гигантские скопления объединённых хроматид, возникающие в некоторых типах специализированных клеток. Классическим примером являются гигантские хромосомы в клетках слюнных желёз личинок Drosophila melanogaster. Политенные хромосомы во много раз превышают по размеру хромосомы обычных соматических клеток. Они, как правило, в 100—200 раз длиннее и в 1000 раз толще (содержат до 1000 хромосом), чем хромосомы многих интерфазных клеток (как половых, так и соматических). Так, у личинок D. melangaster общая длина четырёх пар хромосом в слюнных железах составляет 2000 мкм, а в обычных соматических клетках эта величина равна 7, 5 мкм. Характерная форма и размеры политенных хромосом достигаются вследствие их максимальной деспирализации и многократного воспроизедения хромосом без их последующего расхождения, то есть они образуются как результат эндомитоза
Политенные хромосомы Чтобы уловить изменения в структуре хроматина на уровне индивидуальных генов, необходимо изучать растянутые интерфазные хромосомы. В обычных клетках это невозможно, потому что нити интерфазного хроматина слишком тонки и запутаны. Благодаря же явлению политении на интерфазных хромосомах отчетливо видны многочисленные поперечные полосы, частота чередования которых позволяет предположить, что они соответствуют индивидуальным генам. Политенные хромосомы (гигантские хромосомы) содержат во много раз больше ДНК, чем обычные. Они не изменяют своей формы на протяжении всего митотического цикла и достигают длины до 0, 5 мм, а толщины до 25 мкм. Они встречаются, например, в слюнных железах двукрылых (мух, комаров), в макронуклеусе инфузории и в тканях завязи бобов. Чаще всего они видны в гаплоидном числе, т.к. гомологичные хромосомы бывают тесно спарены. Клетки с такими хромосомами вырастают до необычно большого размера. Возникают политенные хромосомы вследствие многократно повторяющегося процесса редупликации ДНК. При этом разные участки ДНК редуплицируются в разной степени. Большинство генетически информативных областей реплицируются 1000 раз, а некоторые - более чем 30 тыс. раз. При этом циклы редупликации ДНК не сопровождаются делением клетки. По существу, политенные хромосомы представляют собой пучки множества неполностью разделенных, тесно прилежащих друг к другу индивидуальных хроматиновых нитей. В частности, политенные хромосомы слюнных желез личинки Drosophila содержат 1024 таких нити. Итак, интерфазные политенные хромосомы хорошо видны в световой микроскоп, хроматиновые петли в них располагаются в линейном порядке, при окрашивании этих хромосом заметны перемежающиеся поперечные полосы: темные - диски и светлые - междисковые участки. Предполагают, что именно диски содержат 1024 плотно уложенных гомологичных петли индивидуального петельного участка и расположенные там гены. Структурная организация и функция ДНК междисковых участков пока что неизвестна. С началом транскрипции генов диски, в которых они содержатся, декомпактизуются, становятся как бы вздувшимися и называются пуфами. Образующая их ДНК упакована гораздо менее плотно. По всей видимости, подобные структурные модификации хроматина, когда происходит его частичная деконденсация, и являются первым этапом активации эукариотических генов. Биохимически пуфы содержат меньше гистона HI, много РНК-полимеразы и, как минимум, один общий негистоновый белок. Возможно, функциональная единица генома и у высших кариот, в том числе у человека, устроена и функционирует так же. Эндомитоз (от лат. эндо... и митоз) — процесс удвоения числа хромосом в ядрах клеток многих протистов, растений и животных [1], за которым не следует деления ядра и самой клетки. В процессе эндомитоза (в отличие от многих форм митоза) не происходит разрушения ядерной оболочки и ядрышка, не происходит образование веретена деления и не реорганизуетсяцитоплазма, но при этом (как и при митозе) хромосомы проходят циклы спирализации и деспирализации. Повторные эндомитозы приводят к возникновению полиплоидных ядер, отчего в клетке увеличивается содержание ДНК. Также эндомитозом называют многократное удвоение молекул ДНК в хромосомах без увеличения числа самих хромосом; как результат образуются политенные хромосомы. При этом происходит значительное увеличение количества ДНК в ядрах. ЭНДОМИТОЗ Эндомитоз – процесс, который приводит к образованию полиплоидных клеток. Это происходит вследствие блокады митоза либо сразу после премитотического периода интерфазы(G2), либо на стадии профазы или метафазы, после чего клетки переходят в состояние интерфазы и могут приступить опять к репликации ДНК, что приводит к полиплоидии.
Появление полиплоидных соматических клеток может происходить и в результате отсутствия цитотомии. Если такая двуядерная клетка снова вступит в синтетический период интерфазы, то оба ядра становятся не диплоидными, а тетраплоидными. Подобным образом образуются полиплоидные клетки в печени, эпителии мочевого пузыря и др. Характерно, что полиплоидизация встречается на терминальных этапах развития клеток, как правило, характерна для специализированных, дифференцированных клеток и не встречается при образовании половых и стволовых клеток, а также в ходе эмбриогенеза (исключая провизорные органы). 17. Особенности строения метафазных хромосом. Типы хромосом. Хромосомный набор. Правила хромосом. Основы цитогенетики. Строение и типы метафазных хромосом человека. Многие ученые в разных странах мира изучали хромосомы клеточного ядра. Однако только в 1955 году Тио и Леван установили, что в большинстве клеток у человека присутствует 46 хромосом. Хромосомы (в переводе – окрашенные тельца) формируются в начале деления клеток из хроматина интерфазного ядра. Хромосомы являются носителями наследственной информации, передаваемой из поколения в поколение у большинства живых организмов, в том числе и у человека. Хроматин состоит из молекул ДНК, связанных с белками. Эти нити можно рассмотреть только при помощи электронного микроскопа. Они составлены из расположенных друг за другом микрочастиц – нуклеосом, диаметр которых около 10 нм. Нуклеосома имеет белковых остов, вокруг которого закручено молекула ДНК. В течение жизненного цикла клетки хромосомы подвергаются определенным изменениям. В периоде покоя и интерфазы они составляют хроматин клеточного ядра. В таком положении хроматин активно участвует в жизнедеятельности клетки, обеспечивая образование белков и молекул РНК. В S-синтетическом периоде происходит удвоение всех хромосом. Это необходимо для того, чтобы в результате деления вновь образовавшиеся клетки получили такой же набор наследственного материла, какой имела их предшественница. Во время деления нити хроматина сильно спирализуются, закручиваются и утолщаются, формируя видимые в световой микроскоп хромосомы. Именно поэтому основные сведения о строении хромосом были получены во время митоза. Так как к моменту деления хромосомы удвоены, то в световой микроскоп они видны состоящими из двух нитей – хроматид. Обе хроматиды объединены между собой в области первичной перетяжки – центромеры. Центромера делит хромосому поперек на две части – плечи, которые бывают короткими (p) и длинными (q). В зависимости от расположения центромеры различают 3 типа хромосом: - метацентрические, в которых центромера расположена в середине хромосомы и плечи имеют одинаковый размер (p=q); - субметацентрические, центромера в которых сдвинута к одному концу хромосомы. При этом различаются короткие и длинные плечи; - акроцентрические, когда визуально можно определить у хромосомы только длинные плечи. При этом центромера располагается на конце хромосомы или близко от него. Некоторые хромосомы могут иметь дополнительные перетяжки, которые называются вторичными. Если вторичная перетяжка располагается близко к концу хромосомы, то отделяемый ею участок называется спутником. Хромосомы, как отдельные структуры, становятся доступными для исследования только после значительной конденсации хроматина, которая наступает во время митоза (в соматических клетках), либо во время мейоза (при образовании половых клеток). Начавшаяся в профазе конденсация хроматина заканчивается в метафазе, поэтому, как правило, хромосомы изучаются на стадии метафазной пластинки. В интерфазе хромосомы находятся в деконденсированном состоянии, и определить их как отдельные структуры не представляется возможным. В метафазе каждая хромосома имеет как бы иксобразную форму и состоит из двух идентичных половин - хроматид (сестринских хромосом), тесно прилежащих друг к другу только в области первичной перетяжки ( центромеры ), а на остальном протяжении между хроматидами видна большая щель. Центромера - это тот участок, где хромосома находится в деконденсированном состоянии, и к ней прикрепляются нити веретена деления. Центромера делит хромосомы на плечи. По положению центромеры выделяют три вида хромосом. 1. Метацентрические, у которых плечи имеют примерно одинаковую длину (т.е. центромера расположена посередине хромосомы). 2. Субметацентрические, у которых центромера смещена от середины, располагается субмедиально и делит хромосому на два плеча неравной длины. Верхнее всегда меньшее. З. Акроцентрические, у которых центромера расположена почти на конце хромосомы, отделяя от длинного плеча очень короткое верхнее плечо. Верхние короткие плечи принято обозначать буквой " р", а нижние длинные буквой " q". Характерной чертой для некоторых хромосом является наличие вторичных перетяжек, они возникают в участках неполной конденсации хромосом и располагаются в околоцентромерных участках 1-й, 9-й и 16-й хромосом. Вторичные перетяжки имеются также в 13-15 и 21 -22-й хромосомах, однако здесь они занимают удаленное от центромеры положение, отделяя небольшой концевой участок короткого плеча хромосом в виде спутника. Эти хромосомы называют спутничными. В этих хромосомах в области вторичной перетяжки сосредоточены гены, кодирующие р-РНК, и в прилежащих участках кариоплазмы образуются ядрышки. Поэтому такого рода вторичные перетяжки называют ядрышковыми организаторами. В хромосомных наборах одних людей указанные хромосомы имеют вторичную перетяжку, а в этих же хромосомах у других людей её может не быть
Типы строения хромосом Различают четыре типа строения хромосом: - телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце); - акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом); - субметацентрические (с плечами неравной длины, напоминающие по форме букву L); - метацентрические (V-образные хромосомы, обладающие плечами равной длины). Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода. Хромосомы человека. 1956 год - шведы Тио и Леван, англичане Форд и Хамертон установили, что ядро диплоидной клетки человека содержит 46 хромосом – это хромосомный набор или кариотип человека; Совокупность всех метафазных хромосом, расположенных относительно произвольно в экваториальной плоскости клетки, именуется метафазной пластинкой или просто хромосомным набором. После приготовления препаратов хромосом, которые можно приготовить из всех тканей и клеточных суспензий, содержащих делящиеся клетки (в зависимости от целей важно, конечно, количество метафаз), хромосомы окрашиваются, ибо только после этого их можно различить в световой микроскоп, получить микрофотографию, идентифицировать и, расположив их в определенном порядке, т.е. составив кариограмму, получить целостное представление о кариотипе конкретного человека. Кариограмма - это те же хромосомы метафазной пластинки, но расположенные упорядоченно. Принцип упорядоченности общий для всего вида и определяется идиограммой. Идиограмма - это графическое изображение гаплоидного набора хромосом (можно и диплоидного) и расположение их по группам в зависимости от формы и величины. Группы располагаются в порядке уменьшения величины входящих в них хромосом. Правила хромосом: Популярное:
|
Последнее изменение этой страницы: 2016-04-10; Просмотров: 1914; Нарушение авторского права страницы
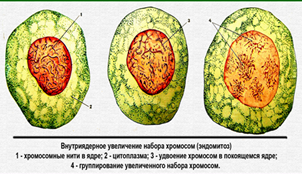 Рис.30. Эндомитоз.
Рис.30. Эндомитоз.