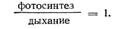|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Показатели фотосинтеза: интенсивность, фотосинтетический потенциал, индекс листовой поверхности; суточные и сезонные ритмы фотосинтетических процессов.
ИНДЕКС ЛИСТОВОЙ ПОВЕРХНОСТИ выраженная в квадратных сантиметрах (см2) площадь освещенных листьев на каждый квадратный сантиметр поверхности почвы; листовые индексы можно считать мерой фотосинтезирующей биомассы. Максимальная чистая продукция соответствует листовому индексу, близкому к 4 (т. е. когда площадь освещенных листьев в 4 раза больше площади, занятой растениями), тогда как максимум валовой продукции достигается при листовом индексе 8 — 10. Этот уровень характерен для спелых лесов. Фотосинтетический потенциал - способность растения к фотосинтезу. Иногда, вследствии перенесенного стресса или болезни, растение теряет способность к нормальному фотосинтезу и требуется стимуляция с применением специальных препаратов, способных повлиять на внутриклеточные процессы. Интенсивность (скорость) фотосинтеза - это количество углекислого газа, которое усваивается единицей листовой поверхности за единицу времени. В зависимости от вида растения этот показатель колеблется от 5 до 25 мг СО2/дм2. ч. Продуктивность фотосинтеза - это отношение суточного увеличения массы всего растения (в граммах) к площади листьев. В среднем эта величина составляет от 5 до 12 г сухого вещества на 1 м2 листовой поверхности в сутки. Интенсивность фотосинтеза можно определять: газометрическими методами, радиометрическими методами. С помощью газометрических методов можно определить либо количество усвоенного углекислого газа, либо количество выделенного кислорода. При этом используют как весовые показатели поглощаемых или выделяемых газов, так и объемные показатели, показатели давления, показатели окраски, показатели теплопроводности определяемых газов. С помощью радиометрических методов определяют интенсивность поглощения С14О2 растением по наличию в нем С14 или изменение радиоактивности газовой смеси. Продуктивность фотосинтеза определяют по накоплению ассимилятов в растении. При этом используют такие методы, как: изменение количества сухого вещества высечек из листа через определенный временной промежуток, накопление углеводов в листе через определенный временной промежуток, изменение теплоты сгорания сухого вещества листьев за период экспозиции их на свету. В процессе вегетации интенсивность и продуктивность фотосинтеза возрастают постепенно от начала развития, достигают максимума в фазе цветения-плодообразования, а затем постепенно убывают. Суточные и сезонные ритмы ф.. С восходом солнца интенсивность ф. возрастает вместе с освещенностью, достигая максимальных значений В 9-12 ч. Дальнейший характер процесса определяется степенью оводненности листьев, температурой воздуха и интенсивностью солнечного света. В полуденные часы интенсивность фотосинтеза не увеличивается: она может оставаться примерно на уровне утреннего максимума (в нежаркие, облачные дни) или несколько снижается, но тогда к 16-17ч наблюдается повторное усиление процесса. Интенсивность ф. падает после 22 ч с заходом солнца. Дневная депрессия фотосинтеза (если имеет место) связана с нарушениями в деятельности фотосинтетического аппарата и оттока ассимилятов при перегреве, поскольку температура листьев в этот период может превышать температуру воздуха на 5—10°С. Если потери воды тканями велики и наблюдается усиление фотодыхания, то устьица в это время закрываются. Сезонные изменения ф., изученные О. В. Заленским у растений пустынь и в условиях Арктики, показали, что у пустынных растений они зависят от особенностей онтогенеза, а у эфемеров с коротким вегетационным периодом максимальные интенсивности фотосинтеза наблюдаются в конце марта — середине апреля и совпадают с началом плодо ношения. У растений, заканчивающих активную вегетацию в начале лета, сезонный максимум фотосинтеза отмечается перед наступлением летнего покоя. У длительно вегетирующих деревьев и кустарников сезонный максимум регистрируется в самом начале жаркого и сухого периода. К осени интенсивность ф. постепенно снижается. У арктических растений сезонные изменения ф. проявляются в снижении его интенсивности в начале и в конце периода вегетации, когда растения часто подвержены действию заморозков. Максимум ф. отмечается в наиболее благоприятный период полярного лета. 26. Фотосинтез и урожай; зависимость фотосинтеза от факторов окружающей среды (температуры, условий освещения (интенсивности, спектрального состава света), содержания углекислоты, условий минерального питания, водоснабжения). Фотосинтетическая деятельность растений зависит от многих внешних факторов, и главные из них - условия освещения (интенсивность, спектральный состав света), доступность и концентрация углекислого газа, температура среды, водоснабжение и минеральное питание. Факторы внешней среды (экзогенные, экологические), воздействуя на отдельные реакции фотосинтеза, вызывают изменение активности фотосинтетического аппарата в целом, что в конечном итоге определяет общую продуктивность растений. Влияние на фотосинтез основных экзогенных факторов рассматривается здесь раздельно, но следует помнить, что в природе они действуют на растение одновременно, и его продуктивность является, таким образом, интегральной функцией совокупности экологических факторов 1 ВЛИЯНИЕ НА ФОТОСИНТЕЗ ИНТЕНСИВНОСТИ И СПЕКТРАЛЬНОГО СОСТАВА СВЕТА Зависимость фотосинтеза от интенсивности света выражается световой кривой фотосинтеза. Как видно из рисунка, она имеет логарифмический характер: при низких интенсивностях света существует прямая пропорциональная зависимость между интенсивностью падающего на растение света и фотосинтеза. По мере увеличения интенсивности света нарастание фотосинтеза становится все менее выраженным, и, наконец, при достижении определенного уровня освещенности наступает насыщение фотосинтетической активности листа (плато световой кривой фотосинтеза). Анализ световой кривой фотосинтеза послужил основанием для представления о существовании в этом процессе световых, фотохимических, реакций, прямо пропорционально зависящих от интенсивности света, и «темновых», энзиматических, реакций, скорость которых становится лимитирующей при высоких интенсивностях света (F. Black-man, 1905).Зависимость световых реакций фотосинтеза от интенсивности света отражает прямолинейный участок световой кривой. На этом этапе интенсивность света является лимитирующим фактором фотосинтеза. Угол наклона прямолинейного участка световой кривой характеризует эффективность использования световой энергии в ходе фотохимических реакций.Стационарный уровень фотосинтеза достигается у разных групп растений при разной интенсивности света. Фотосинтетическая активность в области насыщающих интенсивностей света зависит главным образом от работы систем поглощения и ассимиляции углекислого газа. Чем выше положение кривой в области насыщения, тем больше мощность системы ассимиляции углекислого газа у растения. Условия внешней среды, активирующие углеродные циклы растения (такие, как температура и концентрация С02), позволяют ему более эффективно использовать высокие интенсивности света. На световой кривой фотосинтеза обычно отмечается световой компенсационный пункт (СКП) — наименьшая интенсивность света, при которой активности фотосинтеза и дыхания растений уравновешены по газообмену. Положение СКП зависит от соотношения фотосинтеза и темнового дыхания и меняется в зависимости от температуры и концентрации углекислого газа в среде.Интенсивность света влияет на характер фотохимических и метаболических процессов в хлоропластах. При низких интенсивностях света в первую очередь активируется нециклический поток электронов. Увеличение интенсивности света стимулирует циклический поток электронов и повышает долю циклического транспорта электронов в общем потоке электронов по ЭТЦ хлоропластов. В условиях избыточной освещенности циклический транспорт электронов может играть защитную роль в хлоропластах, а также служить источником энергии для дополнительного синтеза АТФ и тем самым способствовать активации процессов ассимиляции углерода. С интенсивностью света связано образование различных продуктов фотосинтеза. Так, при слабой освещенности образуются главным образом аминокислоты, тогда как синтез углеводов в хлоропластах требует высокой интенсивности света.Спектральный состав света определяет общую интенсивность фотосинтеза, активность его отдельных реакций и набор синтезируемых продуктов. Кривая зависимости интенсивности фотосинтеза от качества света при выровненном количестве квантов (спектр действия фотосинтеза) демонстрирует два отчетливых максимума — в синей и красной области спектра, совпадающих со спектрами поглощения фотосинтетических пигментов. Максимумы поглощения отражают наиболее эффективное использование этих лучей при фотосинтезе.В опытах Н. П. Воскресенской было показано специфическое действие синего света на фотосинтетический аппарат растений. При выравнивании синего и красного освещения по квантам на синем свету у растений увеличивалась общая активность ассимиляции С02, что было обусловлено активацией в этих условиях транспорта электронов в ЭТЦ хлоропластов и ферментов углеродного цикла. Качество света определяло и состав продуктов, образующихся при фотосинтезе. Так, на синем свету преимущественно синтезировались органические кислоты и аминокислоты, а позднее — белки, тогда как красный свет индуцировал сначала синтез растворимых углеводов, а в дальнейшем — крахмала.2 ВЛИЯНИЕ НА ФОТОСИНТЕЗ КОНЦЕНТРАЦИИ УГЛЕКИСЛОГО ГАЗАСодержание С02 в окружающей среде является одним из важнейших факторов, определяющих скорость фотосинтеза. Как видно, зависимость интенсивности фотосинтеза от концентрации С02 имеет логарифмический характер, т. е. увеличение концентрации С02 приводит к быстрому увеличению интенсивности фотосинтеза. При концентрации С02 0, 06 — 0, 15 % у большинства растений достигается насыщение фотосинтеза. Увеличение интенсивности фотосинтеза при повышении концентрации С02 обусловлено реализацией в этих условиях потенциальной карбоксилазной активности Рубиско и созданием в хлоропластах большого пула акцептора С02 — рибулозобисфосфата.Концентрация С02, при которой его поглощение в процессе фотосинтеза уравновешено выделением углекислоты в ходе дыхания (темнового и светового), называется углекислотным компенсационным пунктом (УКП). Положение УКП может существенно зависеть от вида растения. Особенно выражены различия между С3- и С4-растениями. Так, у С3-рас-тений УКП находится при довольно высоких концентрациях С02 (около 0, 005 %). Это связано с наличием у них активного фотодыхания. С4-растения, обладающие способностью фиксировать С02 через фермент ФЕП-карбоксилазу, производят рефиксацию углекислого газа при слабом фотодыхании, поэтому у них УКП приближается к нулевой концентрации С02.3 ВЛИЯНИЕ ТЕМПЕРАТУРЫ НА ФОТОСИНТЕЗИнтегральный ответ фотосинтетического аппарата на изменения температуры может быть представлен графически в виде, как правило, одновершинной кривой (рис. 3.49), причем вершина соответствует области оптимальных для фотосинтеза температур. Для большинства С3-растений умеренной климатической зоны оптимальная для фотосинтеза температура находится в интервале 20 — 25 " С. У растений с С4-путем фотосинтеза и САМ-фотосинтезом температурный оптимумприходится на 30 — 35 °С. При дальнейшем повышении температуры фиксация С02 уменьшается вследствие снижения тургора в листьях и закрывания устьиц.В растениях наибольшую термозависимость проявляют реакции углеродных циклов. Весьма чувствительны к температуре также транспорт электронов и синтез АТФ. Первичные реакции фотосинтеза, связанные с поглощением света, миграцией энергии возбужения в реакционные центры и разделением зарядов в реакционных центрах, от температуры практически не зависят.4 ВЛИЯНИЕ ВОДНОГО РЕЖИМА НА ФОТОСИНТЕЗЗначение водного режима для фотосинтеза определяется в первую очередь действием воды на состояние устьиц листа: до тех пор, пока устьица остаются оптимально открытыми, интенсивность фотосинтеза не изменяется под влиянием колебаний водного баланса. Частичное или полное закрывание устьиц, вызванное дефицитом воды в растении, приводит к нарушению газообмена и снижению поступления углекислого газа к карбоксилирующим системам листа. Вместе с тем, при водном дефиците снижается активность ферментов ВПФ цикла (цикла Кальвина), обеспечивающих регенерацию рибулозобисфосфата, и в значительной степени ингибируется фотофосфорилирование. В результате в условиях водного дефицита наблюдается ингибирование фотосинтетической активности растений. При длительном дефиците воды возможно снижение общей фотосинтетической продуктивности растений, в том числе и за счет уменьшения величины листьев, а сильное обезвоживание может в итоге вызвать нарушение структуры хлоропластов и полную потерю их фотосинтетической активности.Большинство растений имеет наибольшую интенсивность процесса при температурах между 20—28 °С. Вверх и вниз от этих границ идет уменьшение интенсивности фотосинтеза. Есть, однако, исключения из этого правила. Так, например, бурые и багряные водоросли наших северных морей имеют оптимум, лежащий при 7—8 °С. При повышении температуры у них начинает очень сильно возрастать дыхание и падает фотосинтез, так что они быстро переходят за компенсационный пункт. Компенсационным пунктом (точкой) называется отношение фотосинтеза к дыханию, при котором фотосинтез равен дыханию:
Когда дыхание будет превосходить фотосинтез, начнется трата вещества, запасенного ранее растением. Если такой процесс будет продолжаться длительное время, он должен рано или поздно истощить растение и привести его к гибели.О степени использования растением фотохимической активности хлорофилла можно судить по так называемому ассимиляционному числу, которое соответствует количеству ассимилированного углекислого газа весовой единицей хлорофилла в единицу времени. У растений с темно-зелеными листьями оно невелико, а бледно-зеленых растений — более высокое. Основное поглощение солнечной радиации происходит в верхних ярусах посева. В листьях этих ярусов содержится и наибольшее количество хлорофилла. Лучистую энергию, падающую на лист, частично поглощают, а частично отражают и пропускают ткани листа. Этот процесс можно изобразить формулой: Q = R + Т + A, где Q — количество радиации, падающей на лист; R, Т, А — соответственно отраженная, пропущенная и поглощенная радиация, % от падающей радиации (эти показатели в значительной мере зависят от содержания хлорофилла в листьях).Количество хлорофилла — важный фактор, влияющий на работоспособность фотосинтетического аппарата. Поэтому в сельском хозяйстве нужно всегда заботиться о достаточном содержании хлорофилла в растениях, создавая оптимальную структуру посевов и обеспечивая растения необходимыми элементами питания (азотом, калием, фосфором, магнием, микроэлементами). Научные основы учения о роли кислорода в дыхании были заложены трудами А.Л.Лавуазье. В 1774 г. кислород независимо открыли Пристли и Шееле, а Лавуазье дал название этому элементу. Изучая одновременно процесс дыхания животных и горение, Лавувзье в 1773-1783 гг. пришел к выводу, что при дыхании, как и при горении, поглощается кислород и образуется углекислый газ, причем в том и другом случаях выделяется тепло. На основании своих опытов он заключил, что процесс горения состоит в присоединении кислорода к субстрату и что дыхание есть медленно текущее горение питательных веществ в живом организме. Я.Ингенхауз в 1778-1780 гг. показал, что зеленые растения в темноте, а незеленые части растений и в темноте, и на свету поглощают кислород и выделяют углекислый газ. В своей работе, опубликованной в 1779 г. он писал: «Когда солнце, поднявшееся над горизонтом, разбудит своими лучами заснувшие за ночь растения, оно сделает их способными исполнять свою целительную функцию – исправлять воздух для животных; во мраке ночи эта деятельность совсем прекращается; днем же совершается с тем большим оживлением, чем светлее день и чем выгоднее расположено растение в отношении солнечных лучей. Затененные высокими зданиями или другими растениями, они не исправляют воздух, а, наоборот, выделяют вредный для дыхания животных воздух. К концу дня выработка очищенного воздуха ослабевает и при заходе солнца совершенно прекращается». Первые точные исследования процесса дыхания у растений принадлежат Соссюру (1804). Он брал свежие листья и помещал их на ночь в сосуд, наполненный воздухом. При этом кислород воздуха поглощался и выделялся углекислый газ. Если на следующий день листья снова выставлялись на солнечный свет, то они выделяли почти такое же количество кислорода, какое поглотили ночью. Свои исследования Соссюр распространил и на незеленые части растений: стебли древесных растений, цветки, корни, плоды, и доказал, что дыхание наблюдается также в клетках этих органов. Он обнаружил, что при дыхании потеря в весе растения равна весу выделенного углерода. Соссюр обратил внимание и на то, что молодые, растущие части растения, например новые побеги и распускающиеся цветки, дышат интенсивнее и потребляют кислорода больше, чем части растения, прекратившие рост. Если, по Лавуазье, дыхание имеет сходство с процессом горения, то каким же образом органические вещества могут «гореть» при обычной температуре тела организма, да еще в водной среде, (ведь на 70 — 90% масса живых организмов состоит из воды)? Возникло предположение о том, что в живых клетках существуют механизмы, активирующие кислород. Швейцарский химик X. Ф. Шейнбайн, открывший озон, изучал причины быстрого потемнения пораненной поверхности растительных тканей, таких, как ткани яблок, картофеля, плодовых тел грибов. В 1845 г. он выступил со своей теорией окислительных процессов, согласно которой в живых клетках имеются соединения, способные легко окисляться в присутствии 02 и таким образом активировать молекулярный кислород. Если ткань прокипятить, то потемнения не происходит. Следовательно, потемнение тканей — каталитический окислительный процесс. Шейнбайн ошибочно полагал, что активация кислорода — это образование озона. Исследования, начатые Шейнбайном, продолжил А. Н. Бах, который в 1897 г. разработал перекисную теорию биологического окисления, приложив ее к процессам дыхания. Несколько позже, в том же 1897 г., аналогичные взгляды высказал немецкий исследователь К. Энглер. Суть перекисной теории биологического окисления Баха заключается в следующем. Молекулярный кислород имеет двойную связь и для того чтобы его активировать, необходимо эту двойную связь расщепить. Легко окисляющееся соединение А взаимодействует с кислородом и, разрывая двойную связь, образует пероксид А02 Таким образом, по мысли Баха, активация кислорода есть образование пероксида. В свою очередь пероксидное соединение, взаимодействуя с соединением В, окисляет его; затем эта реакция повторяется со вторым атомом кислорода и второй молекулой соединения В. Получается полностью восстановленное исходное соединение — акцептор кислорода А и полностью окисленное вещество В. Много позднее, в 1955 г., две группы исследователей — О. Хаяиши с сотр. в Японии и Г. С. Мэзон с сотр. в США, используя современные методы, проанализировали возможность включения кислорода в органические соединения. В настоящее время известно, что путь включения кислорода в органические соединения в соответствии с перекисной теорией биологического окисления Баха и Энглера не имеет отношения к дыханию, однако работы этих исследователей сыграли большую роль в изучении химизма дыхания, заложив основы современного понимания механизмов активации кислорода. История современного учения о дыхании растений неразрывно связана с именем академика В.И. Палладина. В годы первого петербургского периода работы Палладин исследовал ферментативную природу дыхательного процесса. Палладин показал, что и анаэробная, и аэробная фазы дыхания обеспечиваются специфическими ферментами, последовательно перерабатывающими продукты дыхания. Итоги работ этого периода изложены в монографии В.И. Палладина «Дыхание как сумма ферментативных процессов» (1907). Одновременно с Палладиным проблемой дыхания занимались в целом ряде крупнейших научно-исследовательских институтов и лабораторий Западной Европы. Наибольшую популярность приобрели две новые школы – Виланда и Варбурга. Т. Виланд развивал взгляды на роль дегидраз и водородных акцепторов, вполне аналогичные взглядам Палладина. Расхождение их теорий заключалось в том, что Виланд категорически отрицал какую бы то ни было роль оксидаз как специфических активаторов кислорода, считая молекулярный кислород способным самостоятельно отнимать водород от водородного акцептора. По мнению же Палладина, водородные акцепторы не могут самопроизвольно освобождаться от водорода, но требуют для этого участия оксидаз, которые поэтому являются обязательным фактором в реакции, выраженной во втором уравнении Палладина. Противник Виланда, Варбург, считал, что молекулярный кислород не может вступить в организме в какой бы то ни было окислительный процесс, если в организме отсутствует система железоорганических соединений, типичным представителем которых он считал геминфермент. Варбург утверждал, что геминфермент активирует молекулярный кислород, т.е. как бы дает первый толчок к началу окислительных процессов, и без него никакой дыхательный процесс не может совершаться. Далее, по мнению Варбурга, окислительный импульс через промежуточные звенья (геминовые соединения) доходит до дыхательного субстрата и окисляет его. Резюмируя свои взгляды, Варбург утверждал, что дыхание осуществляется путем активации кислорода, а отнюдь не водорода. Но ведь Палладин как раз и говорил о той же необходимости активации молекулярного кислорода, защищая перед Виландом роль оксидаз в процессе дыхания. Все различие в основных посылках Варбурга и Палладина заключается в том, что первый, работая по преимуществу с объектами животного происхождения, называл свой активатор молекулярного кислорода геминферментом, а Палладин, работавший с объектами растительного происхождения, сохранил за этим активатором ранее установившееся в науке название оксидазы. Но по существу оба говорили об одном и том же, протестуя против непримиримой позиции Виланда, отрицавшего необходимость энзиматической активации молекулярного кислорода. Английский биохимик Д. Кейлин в 1925 г. доказал присутствие в клетках цитохромоксидазы, ускоряющей поглощение ими кислорода, и открыл другие цитохромы. Затем цитохромы были обнаружены у всех аэробов и было показано, что у этих организмов на завершающем этапе процесса дыхания осуществляется перенос на кислород электронов и протонов, в результате чего образуется Н20 (или Н202). Унификация субстратов дыхания, дыхательные субстраты, механизм активации дыхательных субстратов, пути их включения в процессы биологического окисления; взаимосвязь превращения углеводов, белков и жиров. Вопрос о веществах, используемых в процессе дыхания, издавна занимал физиологов. Еще в работах И. П. Бородина было показано, что интенсивность процесса дыхания прямо пропорциональна содержанию в тканях растений углеводов. Это дало основание предположить, что именно углеводы являются основным веществом, потребляемым при дыхании. В выяснении данного вопроса большое значение имеет определение дыхательного коэффициента. Дыхательный коэффициент (ДК) — это объемное или молярное отношение СО2, выделившегося в процессе дыхания, к поглощенному за этот же промежуток времени О2. При нормальном доступе кислорода величина ДК зависит от субстрата дыхания. Если в процессе дыханий используются углеводы, то процесс идет согласно уравнению СбН12Сб + 6О2 = 6СО2 + 6Н2О. В этом случае ДК равен единице. Однако если разложению в процессе дыхания подвергаются более окисленные соединения, например органические кислоты, поглощение кислорода уменьшается, ДК становится больше единицы. Так, если в качестве субстрата дыхания используется яблочная кислота, то ДК = 1, 33. При окислении в процессе дыхания более восстановленных соединений, таких, как жиры или белки, требуется больше кислорода и ДК становится меньше единицы. Так, при использовании жиров ДК = 0, 7. Определение дыхательных коэффициентов разных тканей растений показывает, что в нормальных условиях он близок к единице. Это дает основание считать, что в первую очередь растение использует в качестве дыхательного материала углеводы. При недостатке углеводов могут быть использованы и другие субстраты. Особенно это проявляется на проростках, развивающихся из семян, в которых в качестве запасного питательного вещества содержатся жиры или белки. В этом случае дыхательный коэффициент становится меньше единицы. При использовании в качестве дыхательного материала жиров происходит их расщепление до глицерина и жирных кислот. Жирные кислоты могут быть превращены в углеводы через глиоксилатный цикл. Использованию белков в качестве субстрата дыхания предшествует их расщепление до аминокислот. ПУТИ ДЫХАТЕЛЬНОГО ОБМЕНА Существуют две основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов: 1) гликолиз + цикл Кребса (гликолитический); 2) пентозофосфатный (апотомический). Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от условий внешней среды. Процесс дыхания растений осуществляется во всех внешних условиях, в которых возможна жизнь. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от —50 до +50 °С. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растительных организмов. Процесс дыхания представляет собой центральное звено обмена веществ организма, которое тесно связано с другими процессами метаболизма. Необходимо подчеркнуть, что, несмотря на противоположную направленность двух центральных процессов растительного организма — фотосинтеза и дыхания и на их рассредоточенность в разных органеллах клетки, между ними существует тесная взаимосвязь. Прежде всего, для протекания процесса дыхания нужны органические вещества (субстраты). Такими субстратами в первую очередь являются углеводы, которые у зеленых растений образуются в процессе фотосинтеза. Вместе с тем превращение веществ в процессе фотосинтеза и дыхания идет через ряд сходных промежуточных продуктов. Особенно много сходного в превращениях между фотосинтетическим циклом Кальвина и реакциями пентозофосфатного пути дыхательного обмена. Как в том, так и в другом случае происходят взаимные превращения Сахаров с разной длиной углеродной цепочки (3, 4, 5, 6 и 7 углеродных атомов). По - видимому, несмотря на различное распределение (компартментацию) этих метаболитов в клетке, между ними существует обмен. Иначе говоря, промежуточные продукты дыхания могут быть использованы в процессе фотосинтеза. Одновременно возможен и обратный процесс. Много общего в энергетике фотосинтеза и дыхания в процессах фотосинтетического и окислительного фосфорилирования. Между этими двумя процессами возможен обмен энергетическими эквивалентами. АТФ, образовавшаяся на свету при фотосинтетическом фосфорилировании, может служить основным источником энергии для различных биосинтетических процессов, заменяя АТФ, образовавшуюся в процессе дыхания. С другой стороны, АТФ и НАДФ-Н2, образовавшиеся в процессе дыхания, могут быть использованы для реакций цикла Кальвина. Имеются наблюдения, что на свету основными органеллами, поставляющими АТФ, являются хлоропласты. Многие промежуточные продукты процесса дыхания являются основой биосинтеза важнейших соединений. Уже на протяжении первой, анаэробной фазы дыхания (гликолиз) триозофосфат, преобразуясь в глицерин, может служить источником для синтеза жиров. Пировиноградная кислота путем аминирования может дать аланин. Не менее важное значение имеют и промежуточные продукты цикла Кребса. Например, α -кетоглютаровая и щавелевоуксусная кислоты в процессе аминирования дают аминокислоты —глутаминовую и аспарагиновую. Благодаря реакции переаминирования эти кислоты могут быть источником аминогруппы для других аминокислот и, таким образом, являться важнейшими промежуточными продуктами для синтеза как белка, так и пуриновых и пиримидиновых азотистых оснований. Янтарная кислота, образовавшаяся в цикле Кребса, дает основу для образования порфиринового ядра хлорофилла. Ацетил-КоА служит основой для образования жирных кислот. Поскольку имеется ряд реакций и процессов, благодаря которым отдельные компоненты извлекаются из цикла Кребса, должны быть и обратные процессы, поставляющие их в цикл. Если бы этого не было, скорость превращения в аэробной фазе дыхания заметно бы снизилась. Такими реакциями является окислительное дезаминирование аминокислот, приводящее к образованию органических кислот. Имеет значение также реакция карбоксилирования пировиноградной кислоты или ее фосфорилированной формы, в результате чего образуется щавелевоуксусная кислота. Основной процесс, при котором образуются пентозы в растении, — это пентозофосфатный путь дыхательного обмена. Пентозы входят в состав нуклеотидов, нуклеиновых кислот и ряда коферментов, в том числе таких важных, как никотинамидные (НАД и НАДФ), флавиновые (ФМН, ФАД). Пентозофосфатный путь дыхания является также источником образования эритрозо-4-фосфата. Эритрозофосфат, взаимодействуя с фосфоенолпируватом, образует шикимовую кислоту. Шикимовая кислота — материал для образования ряда ароматических аминокислот, например триптофана, а из триптофана образуется один из главных гормонов роста растений — ауксин (3-индолилуксусная кислота). Рассмотренные связи дыхания и других процессов метаболизма растения не являются постоянными, раз навсегда данными. Они возникают и нарушаются под влиянием как внутренних особенностей растения, так и внешних условий. При неблагоприятных условиях эти нарушения могут быть значительными и даже летальными. Ферментные системы дыхания, участие ферментов различных классов в дыхании, альтернативность каталитических механизмов биологического окисления, механизмы активации водорода субстрата и молекулярного кислорода. Окисление дыхательных субстратов в ходе дыхания осуществляется с участием ферментов. Ферменты как белковые катализаторы, помимо свойств, присущих неорганическим катализаторам, обладают рядом особенностей: высокой активностью, высокой специфичностью по отношению к субстратам и высокой лабильностью. Их пространственная организации зависящая от нее активность изменяются под действием внешних и внутренних факторов. Эти свойства обеспечивают возможность тонкой регуляции обмена веществ на уровне ферментов. Типы окислительно-восстановительных реакций. Существуют четыре способа окисления, и все они связаны с отнятием электронов: 1) непосредственная отдача электронов, например: 2) Отнятие водорода: 3) присоединение кислорода: 4) образование промежуточного гидратированного соединения с последующим отнятием двух электронов и протонов: Оксидоредуктазы. Поскольку окисление одного вещества (донора электронов и протонов) сопряжено с восстановлением другого соединения (их акцептора), ферменты, катализирующие эти реакции, называют оксидоредуктазами. Все они относятся к I классу ферментов: Донор (Д) отдает электроны и протоны, акцептор (А) принимает их, а энзим (Е) осуществляет реакцию переноса. Существуют три группы оксидоредуктаз: а) анаэробные дегидрогеназы передают электроны различным промежуточным акцепторам, но не кислороду; б) аэробные дегидрогеназы передают электроны различным акцепторам, в том числе кислороду; в) оксидазы способны передавать электроны только кислороду. Анаэробные дегидрогеназы. Это двухкомпонентные ферменты, коферментом которых может быть НАД+ (никотинамидадениндинуклеотид): При окислении субстрата НАД+ превращается в восстановленную форму НАДH, а второй протон субстрата диссоциирует в среду (НАДH+ Н+). К анаэробным НАД-зависимым дегидрогеназам относятся такие ферменты, как алкогольдегидрогеназа, лактатдегидрогеназа, малатдегидрогеназа и др. Коферментом анаэробных дегидрогеназ может быть также НАДФ+ (никотинамидадениндинуклеотидфосфат), содержащий на одну фосфатную группировку больше, чем НАД +. НАДФ- зависимыми дегидрогеназами являются изоцитратдегидрогеназа, глюкозо-6-фосфатдегидрогеназа, 6-фосфоглюконатдегидрогеназа и др. Субстратная специфичность фермента зависит от его белковой части. Многие НАД- и НАДФ-зависимые дегидрогеназы нуждаются в присутствии ионов двухвалентных металлов. Например, алкогольдегидрогеназа содержит ионы цинка. Окисленные и восстановленные формы коферментов анаэробных дегидрогеназ могут взаимопревращаться в реакции, катализируемой ферментом НАД(Ф)-трансгидрогеназой: НАДФH + НАД+ = НАДФ+ + НАДH Анаэробные дегидрогеназы передают водород, т. е. электроны и протоны, различным промежуточным переносчикам и аэробным дегидрогеназам. Аэробные дегидрогеназы. Это также двухкомнонентные ферменты, получившие название флавиновых (флавопротеины). Помимо белков, в их состав входит прочно связанная с ними простетическая группа — рибофлавин (витамин В2). Различают два кофермента этой группы: флавинмононуклеотид (ФМН), или желтый дыхательный фермент Варбурга, и флавинадениндинуклеотид (ФАД). ФМН (рибофлавин-5-фосфат) содержит гетероциклическое азотистое основание — диметилизоаллоксазин, спирт рибит (производное рибозы) и фосфат: В ФАД кроме ФМН имеется еще один нуклеотид — аденозинмонофосфата: Активной группой в реакции присоединения и отдачи электронов и протонов в ФМН и ФАД служит изоаллоксазин. Взаимодействие с восстановленным переносчиком, например НАДH, происходит следующим образом: Примером дегидрогеназы, в состав которой входит ФАД, является сукцинатдегидрогеназа. Доноры электронов для аэробных дегидрогеназ — анаэробные дегидрогеназы, а акцепторы — хиноны, цитохромы, кислород. Цитохромная система. Среди оксидаз очень важную роль играют железосодержащие ферменты и переносчики, относящиеся к цитохромной системе. В нее входят цитохромы " и цитохромоксидаза. Включаясь в определенной последовательности в процесс переноса электронов, они передают их от флавопротеинов на молекулярный кислород. Все компоненты цитохромной системы содержат железопорфириновую простетическую группу. При переносе электронов цитохромами железо обратимо окисляется и восстанавливается, отдавая или приобретая электрон и изменяя таким образом свою валентность. В дыхательной цепи направление транспорта электронов определяется величиной окислительно-восстановительного потенциала цитохромов. Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 6347; Нарушение авторского права страницы