|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Грибы: место в системе живого мира, происхождение и эволюция
Грибы - большая группа эукариотных гетеротрофных организмов, включающая, по данным разных авторов, от 100 000 до 250 000 видов. Со времён К. Линнея («Система природы», 1735) грибы традиционно относили к царству растений, куда этот великий систематик поместил их не без значительных сомнений. Однако уже в XIX веке многие ботаники указывали на значительные отличия грибов от растений, и Э. Фриз в 1831г. предложил выделить грибы в самостоятельное царство живого мира. Бурное развитие физиологии и биохимии, особенно во второй половине XX века, вернуло биологов к вопросу о месте грибов в системе живого мира. Накопленные данные о строении и составе клетки грибов всё более и более ставили под сомнение положение грибов в царстве растений [1]. Основное отличие грибов от растений заключается в том, что зелёные растения - это автотрофные первичные продуценты, а грибы - гетеротрофные редуценты. Хитиновые оболочки клеток большинства грибов, некоторые черты углеводного и азотного обменов, в результате которых образуются гликоген, серотонин и мочевина - метаболиты, характерные для животной клетки. Особый путь синтеза лизина, одной из важнейших аминокислот. Состав стеролов - всё это также существенно отличает грибы от растений. С другой стороны, клетка грибов как гетеротрофных организмов столь же существенно отличается и от животной клетки. Основные отличия клеток грибов от простейших - абсорбционный способ питания (всей поверхностью тела), а не голозойный (путем заглатывания пищи); размножение спорами; неподвижность тела (в основном). Таким образом, характер признаков, сближающих грибы с растениями, и признаков, сближающих их с животными, различен. У грибов и растений общими являются в основном морфологические признаки, а у грибов и животных - физиолого-биохимические. Именно поэтому развитие биохимических исследований, совершенствование их методов поставило под сомнение отнесение грибов к царству растений. Современные материалы по биохимии и физиологии, ультраструктуре клетки, составу и строению клеточной оболочки грибов позволяют с большой степенью вероятности считать, что грибы по своему строению, характеру обмена и способу питания занимают промежуточное положение между животными и растениями и имеют признаки, как тех, так и других. Отсюда можно предположить, что грибы выделились в самостоятельный ствол живого мира ещё до разделения живых организмов на животных и растения и представляют отдельную эволюционную (филогенетическую) линию эукариот. На этом основании грибы сейчас рассматривают как самостоятельное царство живого мира Mycota. Сначала 1970-х гг. грибы стали рассматривать как самостоятельное царство живого мира наряду с царствами животных и растений (Тахтаджян, 1973; Whittaker, Margulis, 1978; Margulis, Schwartz, 1982 и др.). Сумчатые, базидиальные грибы, зигомицеты, хитридиомицеты, оомицеты получили статус самостоятельных отделов. В связи с абсорбционным способом питания и предполагаемым происхождением грибов от жгутиковых организмов наибольшее филогенетическое значение в систематике грибов придаётся наличию или отсутствию у них подвижности, строению подвижных стадий (зооспор, гамет) и составу клеточной стенки. Анализ этих признаков показывает, что среди грибов встречаются как постоянно неподвижные организмы, так и подвижные в вегетативном состоянии или только на определённых стадиях развития (бесполое или половое размножение). У грибов, имеющих в цикле развития подвижную стадию, встречаются два типа жгутиков: бичевидный гладкий и перистый. Комбинации этих жгутиков и их расположение дают четыре типа подвижных клеток. В группе неподвижных грибов и у одножгутиковых Chytridiomycetes основной компонент клеточной стенки - хитин - полимер аминосахара в сочетании с глюканом, маннаном или хитозаном. У двужгутиковых оомицетов (Oomycetes) и одножгутиковых с передним перистым жгутиком гифохитридиомицетов (Hyphochytridiomycetes) основной компонент клеточной стенки - целлюлоза. Эти группы также объединяет наличие переднего перистого жгутика и биосинтез лизина, что свойственно бактериям и высшим растениям. У остальных грибов биосинтез лизина идет через а-аминоадипиновую кислоту, что характерно для эвгленовых и животных. По сочетанию основных признаков - подвижности, строению зооспоры или гамет, составу клеточной стенки, пути синтеза лизина, в царстве Mycetalia (Mycota, Fungi) выделяют в настоящее время три самостоятельные эволюционные линии, берущие начало от различных групп древнейших жгутиковых Flagellatae. Этим линиям соответствуют три отдела. Отдел Oomycota (оомикота) характеризуется двужгутиковыми зооспорами или гаметами. Отдел Eumycota (эумикота, или настоящие грибы) в основном объединяет неподвижные грибы, не имеющие в цикле развития зооспор или гамет. Отдел Myxomycota (миксомикота) включает организмы, вегетативное тело которых - голый многоядерный протопласт, способный к амебоидному движению по субстрату. Царство настоящих грибов Fungi, или Mycota, включает четыре отдела: Chytridiomycota, Zygomycota, Ascomycota и Basidiomyco. Надо иметь в виду, что изменения в системах грибов отражают кардинальные перемены во взглядах на происхождение, эволюцию и филогенетические связи между таксономическими группами. Это связано с новыми данными по биохимии грибов и грибоподобных организмов, по составу и строению их клеточной стенки, ультраструктуре клетки и, что особенно важно, структуре генома. В настоящее время принята следующая система грибов и грибоподобных организмов [2]. Царство CHROMISTA Подцарство HETEROCONTAE Грибоподобные организмы Отдел Labyrinthulomycota - Лабиринтуломикота, или сетчатые слизевики класс Labyrinthulomycetes - Лабиринтуломицеты класс Thraustochytridiomycetes - Траустохитридиомицеты Отдел Hyphochytridiomycota - Гифохитридиомикота класс Hyphochytridiomycetes — Гифохитридиомицеты Отдел Oomycota - Оомикота класс Oomycetes - Оомицеты Царство FUNGI (MYCOTA, MYCETALIA) - Настоящие грибы Отдел Chytridiomycota - Хитридиомикота класс Chytridiomycetes - Хитридиомицеты Отдел Zygomycota - Зигомикота класс Zygomycetes - Зигомицеты класс Trichomycetes - Трихомицеты Отдел Ascomycota - Аскомикота, или сумчатые грибы класс Archiascomycetes - Археаскомицеты класс Hemiascomycetes - Гемиаскомицеты, или Голосумчатые класс Ascomycetes (Euascomycetes) - Эуаскомицеты, настоящие сумчатые, или плодо сумчатые класс Loculoascomycetes (Dothideomycetes) - Локулоаскомицеты Отдел Basidiomycota - Базидиомикота класс Basidiomycetes - Базидиомицеты подкласс Homobasidiomycetidae - Гомобазидиомицеты подкласс Heterobasidiomycetidae - Гетеробазидиомицеты класс Urediniomycetes - Урединиомицеты класс Ustilaginomycetes - Устилагиномицеты Анаморфные, несовершенные, или митоспоровые грибы Лишайники, или лихенизированные грибы класс Ascolichenes - Аскомицетные лишайники класс Basidiolichenes - Базидиомицетные лишайники
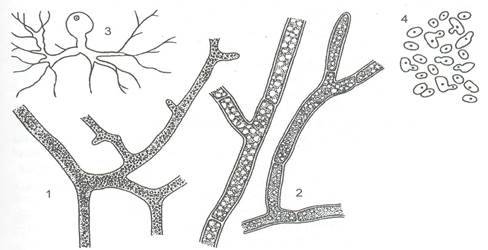
Строение грибов. Вегетативные структуры Типичное вегетативное тело, или таллом, большинства грибов, мицелий, представляет собой систему ветвящихся трубок, гиф, с апикальным ростом и боковым ветвлением Мицелий может быть клеточный и неклеточный (рисунок 1)
1 – неклеточный мицелий, 2 – клеточный мицелий, 3 – одноклеточный таллом с ризомицелием, 4 – почкующиеся клетки, псевдомицилий
Рисунок 1 - Типы мицелия
Неклеточный мицелий лишён перегородок. В течение его роста деления ядер происходят в нём без образования клеточных перегородок - септ, что ведёт к развитию большой массы цитоплазмы, содержащей много ядер. Эта многоядерность мицелия грибов пространственно ограничена в своем разрастании клеточными стенками гиф. Многоядерность мицелия фактически представляет собой одну гигантскую многоядерную клетку. Он характерен для ряда представителей отдела хитридиомикота, а также для представителей отделов оомикота и зигомикота. Другой тип вегетативного тела грибов - клеточный, или септированный, мицелий, разделённый перегородками на одно-, дву- или многоядерные клетки. Он характерен для сумчатых, базидиальных и несовершенных, или анаморфных грибов. Септы могут формироваться и на неклеточном мицелии, обычно это происходит при повреждении мицелия или при образовании репродуктивных органов. При делении клетки септа врастает с боков к центру. В центре септы обычно остаётся пора, через которую из клетки в клетку перемещаются питательные вещества и некоторые клеточные органеллы [4]. Мицелий, пронизывающий субстрат, всей поверхностью абсорбционно поглощающий из него питательные вещества и выделяющий продукты своего обмена, называют субстратным мицелием. Часть мицелия, располагающаяся на поверхности субстрата или над субстратом, составляет поверхностный, или воздушный, мицелий, на котором обычно образуются органы размножения грибов. Существуют и немицелиальные грибы. У части хитридиевых и у гифохитридиевых, являющихся в основном внутриклеточными паразитами водорослей и водных грибов, таллом одноклеточный, микроскопический, иногда даже лишённый в вегетативном состоянии клеточной стенки, которая образуется только при формировании репродуктивных органов. У некоторых грибов из этих групп от такой клетки отходят тонкие разветвлённые нитевидные структуры, лишённые собственных ядер, - ризомицелий, пронизывающий субстрат. Ризомицелий можно рассматривать как зачаточный мицелий, эволюционную ступень к настоящему неклеточному мицелию (рисунок1). При образовании ризомицелия может развиваться только одна клетка, содержащая ядро, которая в дальнейшем становится центром образования ризомицелия и развития репродуктивных органов. У некоторых грибов, например дрожжей, относящихся, в основном, к классу сумчатых, вегетативное тело представлено одиночными почкующимися или делящимися клетками. Если такие почкующиеся клетки не расходятся, то образуется псевдомицелий. Вегетативный мицелий грибов может образовывать различные, часто довольно сложные структуры, выполняющие различные функции. Так, гифы многих паразитных грибов могут формировать расширенные, плотно прилегающие к стенке хозяина клетки, напоминающие присоски - от которых отходят внутрь клеток растения-хозяина особые, часто разветвлённые выросты - гаустории, с помощью которых происходит питание гриба содержимым отдельных клеток. Широко распространены в разных группах грибов склероции - плотные переплетения мицелия, служащие для перенесения неблагоприятных условий. Обычно склероции темно окрашенные, так как наружные слои клеток толстостенные и пигментированные, а внутренние - тонкостенные, светлоокрашенные и богатые запасными питательными веществами (например, склероции возбудителя белой гнили овощных культур или чёрно-фиолетовый рожок спорыньи в колосе ржи). Некоторые грибы образуют склероции, пронизывая и мумифицируя ткань хозяина - растения или животного. В этом случае склероции состоит из ткани хозяина и гиф гриба и повторяет форму хозяина (например, склероции возбудителя плодовой гнили яблок в виде чёрного плода, или склероции гриба кордицепса, паразитирующего на гусеницах, сохраняет форму гусеницы и т.д.) (рисунок 2). Такие склероции называют псевдосклероции.
Рисунок 2 - Псевдосклероций в теле личинки бабочки, проросший стромами с перитециями Близки к склероциям стромы - менее плотные сплетения мицелия, обычно защищающие плодовые тела сумчатых грибов, например, оранжевые головки на проросшем склероции возбудителя спорыньи — это стромы, в которые погружены микроскопические плодовые тела этого гриба. У многих грибов есть структуры, выполняющие проводящие функции. Это мицелиальные тяжи и ризоморфы (рисунок 3). Мицелиальные тяжи состоят из гиф, расположенных параллельно и местами плотно прижатых друг к другу. Между отдельными гифами могут быть мицелиальные мостики. На мицелиальных тяжах, находящихся в почве, формируются зачатки, а затем и сами плодовые тела шляпочных грибов.. Их толщина 4-7 мм, а в длину они могут достигать нескольких метров, что способствует распространению гриба по субстрату. Такие ризоморфы известны у настоящего домового гриба — активнейшего (рисунок 3).
1 - опенка осеннего, 2 - домового гриба, 3 - корневая гниль деревьев при поражении опенком осенним
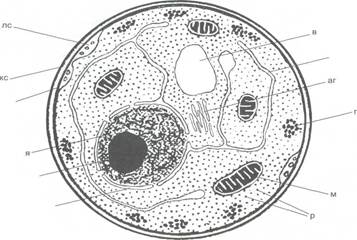
Рисунок 3 - Ризоморфы При плотном переплетении гиф у грибов образуется ложная ткань, плектенхима. Из такой ткани состоят плодовые тела шля-почных грибов. Плектенхима отличается (по происхождению и строению) от настоящей ткани паренхимы, которая возникает в результате деления клеток. Настоящая ткань встречается у грибов очень редко, например, в группе паразитирующих на насекомых лабульбениевых грибов из класса сумчатых таллом состоит из настоящей паренхим. Клетка Вегетативные клетки грибов, за исключением самых примитивных, хитридиомикот и гифохитридиомикот, имеют клеточную оболочку, толщиной около 0, 2 мкм. Она имеет сложную структуру из нескольких слоев, состоящих из аморфного матрикса и микрофибрилл. Клеточная стенка грибов содержит до 80-90 % полисахаридов, связанных с белками и липидами и, в ряде случаев, с пигментами. Аморфный матрикс состоит преимущественно из глюканов, а микрофибриллярные (скелетные) компоненты клеточной стенки - из целлюлозы (у оомикот), хитина (у зигомикот, аскомикот, базидиомикот и анаморфных грибов) и из хитина и целлюлозы (у гифохитридиомикот). У многих дрожжей скелетная часть клеточной стенки состоит из глюканов. [5]. В молодых гифах цитоплазма заполняет всю полость клетки, и цитоплазматическая мембрана (плазмалемма) прижата к клеточной стенке за исключением случайных отхождений. Роль цитоплазматической мембраны - поддержание в клетке осмотического давления и определенного электрического потенциала. Между клеточной оболочкой и плазмалеммой располагаются мембранные структуры, имеющие вид многочисленных пузырьков, ломасомы (рисунок 4).
кс — клеточная стенка, я — ядро, яо — ядерная оболочка, яд — ядрышко, р — рибосомы; м — митохондрии; цм — цитоплазматическая мембрана (плазмалемма); лс — ломасомы; аг — аппарат Гольджи (диктиосомы), эс — эндоплазматическая сеть, ц — цитоплазма, в — вакуоль, г — гликоге
Рисунок 4 - Схема строения клетки грибов Внутренний слой цитоплазмы, эндопласт, содержит систему мембран и полостей, называемую эндоплазматической сетью, которая расположена свободно по всему объёму клетки (у большинства грибов). Положение сети внутри клетки зависит от вида или стадии развития гриба. По сети канальцев, образуемых мембранами эндоплазматической сети, происходит движение веществ от наружной клеточной мембраны к мембране ядра. На наружной поверхности мембраны эндоплазматической сети расположены рибосомы - гранулы, богатые РНК и ответственные за синтез белка. В зоне роста гиф эндоплазматическая сеть образует систему многочисленных канальцев, диктиосом, построенных из гладких мембран, которые составляют аппарат Гольджи. В растущих участках гиф содержатся многочисленные везикулы - пузырьки, связанные своим происхождением с эндоплазматической сетью и участвующие в транспорте веществ от аппарата Гольджи к месту синтеза клеточной стенки. Молодые гифы обычно лишены вакуолей. Небольшие вакуоли, окружённые собственной мембраной, тонопластом, развиваются в более старых частях гиф и позже, сливаясь, образуют крупные вакуоли. В клеточном соке вакуолей, представляющем водный раствор сахаров, белковых веществ, органических кислот и их солей, содержатся пигменты, капли липидов, гранулы гликогена, волютина и белков. Липиды и гликоген, запасные вещества, находятся также и в цитоплазме. Митохондрии грибов в основном похожи на митохондрии растений, но отличаются от них некоторыми деталями строения. Клетка грибов в зависимости от вида и стадии развития содержит 1-2 или много, до 20-30, ядер. Ядра очень мелкие, обычно около 23 мкм диаметром, окружены двухслойной оболочкой. В наружном слое имеются кольцеобразные отверстия, открывающиеся в сторону цитоплазмы. Внутри ядра в нуклеоплазме находятся ядрышко и хроматин. У грибов при митотическом делении ядерная оболочка сохраняется [5]. Размножение грибов У грибов встречаются три типа размножения: вегетативное, бесполое и половое. У многих видов они последовательно сменяют друг друга в цикле развития. Структуры, образующиеся при бесполом и половом размножении, часто сложны по строению и морфологически значительно отличаются друг от друга. Иногда в цикле развития того или иного гриба имеются два и более отличных друг от друга типа спороношения. Такое явление получило название плеоморфизма. Особенно это характерно для аскомикот, или сумчатых грибов. Классическим примером плеоморфизма является цикл развития сумчатого гриба спорыньи (Claviceps purpurea), у которого в цикле развития сменяются три морфологические стадии: бесполая, вегетативная и половая [6]. Вегетативное размножение обычно осуществляется неспециализированными частями мицелия, которые дают начало новому мицелию. Мицелий большинства грибов обладает высокой способностью к регенерации, что и лежит в основе этого способа размножения и широко используется. Например, при приготовлении грибницы для искусственного выращивания съедобных грибов, таких как шампиньон двуспоровый, вешенка обыкновенная, а также при получении биомассы грибов в пищевых и кормовых целях. К специализированным структурам вегетативного размножения относятся оидии, тонкостенные клетки, и хламидоспоры, толстостенные клетки, на которые распадается мицелий, и которые дают начало новому мицелию. Хламидоспоры выполняют и функцию перенесения неблагоприятных условий (рисунок 5). У большинства дрожжей вегетативное размножение происходит путем почкования клеток.
1 - хламидоспоры Mycogone perniciosa - возбудителя мягкой гнили шампиньона
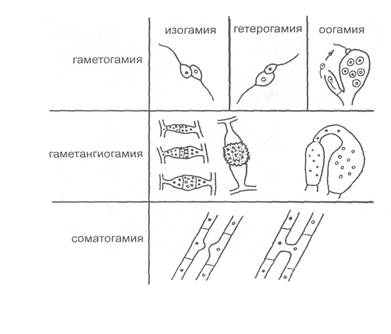
Рисунок 5 - Хламидоспоры грибов в чистой культуре Бесполое размножение осуществляется при помощи разнообразных специализированных клеток, спор. У грибов известны эндогенные и экзогенные споры бесполого размножения. Эндогенные подвижные споры, зооспоры, развиваются в зооспорангиях разной формы. Это голые клетки, снабженные жгутиками, число, расположение и строение которых различно в разных систематических группах грибов. Зооспоры разного строения характерны для представителей водных и реже наземных грибов. Для осуществления размножения с помощью зооспор нужна вода, хотя бы в виде отдельных капель на поверхности почвы или растений, в которой зооспоры могут передвигаться с помощью жгутиков. Эндогенные неподвижные споры, спорангиоспоры, одеты оболочкой и образуются внутри спорангиев, развивающихся на специализированных гифах, спорангиеносцах, обычно поднимающихся над субстратом. Спорангиоспорами осуществляется бесполое размножение у зигомикот. Экзогенные споры бесполого размножения грибов, конидии, неподвижны, образуются на специализированных, обычно морфологически отличных от вегетативного мицелия, дифференцированных спороносцах, конидиеносцах. Типичные конидии характерны для сумчатых и базидиальных. Это в основном наземные грибы, и распространение неподвижных спор бесполого размножения, спорангиоспор и конидий у таких грибов осуществляется в основном пассивно токами воздуха или воды. Иногда распространение спор может осуществляться с помощью животных, например, при поедании плодовых тел шляпочных грибов. Половое размножение у грибов, связанный с ним процесс смены ядерных фаз, строение половых органов - всё это отличается у разных групп грибов. Существеннейшие моменты полового процесса у грибов: плазмогамия, кариогамия и мейоз. Соответственно гриб может находиться в гаплоидной или диплоидной стадиях. Плазмогамия и кариогамия у сумчатых и базидиальных грибов не совпадают во времени, в результате чего после плазмогамии у них возникает особая стадия дикариотичного мицелия, когда гаплоидные ядра попарно ассоциированы, сближены, но не слились и образуют дикарион. Ядра дикариона обычно синхронно делятся с параллельным расположением осей веретен деления. В результате полового процесса образуются гаплоидные, неоднородные в генетическом отношении споры, что принципиально отличает их от спор бесполого размножения грибов. Эти гаплоидные споры располагаются или на мицелии или, чаще, на поверхности или внутри плодовых тел различного строения, которые называются телеоморфами. Таким образом, размножение с помощью спор, возникших половым путем, даёт начало формам с новой комбинацией генетического материала, а размножение спорами, возникшими бесполым путем, способствует распространению и сохранению данной формы. Типы полового процесса у грибов. Половое размножение у грибов известно у всех групп, за исключением несовершенных грибов, за что они и получили такое название. Половой процесс у грибов разнообразен и его особенности лежат в основе выделения классов. У грибов известно три основных типа полового процесса: гаметогамия, гаметангиогамия и соматогамия (рисунок 6). Гаметогамия - слияние гамет, образующихся в гаметангиях.
Рисунок 6 - Типы полового процесса у грибов Различают изогамию - слияние подвижных морфологически сходных гамет и гетерогамию — слияние подвижных, отличающихся по размерам и часто по степени подвижности гамет. Эти два типа гаметогамии характерны для хитридиевых и гифохитридиевых грибов. При оогамии крупные, неподвижные яйцеклетки, формирующиеся в специальных оогониях, оплодотворяются мелкими, подвижными сперматозоидами, развивающимися в специализированных антеридиях. Второй тип полового процесса, гаметангиогамия, состоит в слиянии двух обычно многоядерных специализированных структур, содержимое которых не дифференцировано на гаметы (рисунок 5). Гаметангиогамия характерна для зиго и аскомикот. Гаметангиогамия у зигомикот носит название зигогамии. У аскомикот при гаметангиогамии также сливаются два многоядерных гаметангия, но у них, в отличие от зигомикот, половые органы дифференцированы на женский - аскогон и мужской - антеридий. Аскогон состоит из двух клеток: крупной многоядерной и тонкой нитевидной - трихогины, помещающейся на его вершине, через которую в аскогон переливается содержимое многоядерного антеридия. При этом происходит только плазмогамия, а ядра ассоциируются в пары, образуя дикарион. В их клетках происходит слияние ядер дикариона и образование диплоидного ядра, которое в дальнейшем делится мейотически. В результате этого процесса на аскогенных гифах достаточно сложным путем формируются особые образования - сумки, или аски, внутри которых после митотического деления ядер формируются восемь эндогенных гаплоидных аскоспор. Третий тип, соматогамия, - половой процесс, при котором сливаются обычные соматические, или вегетативные, клетки мицелия. Половые органы и гаметы отсутствуют. В этом случае целиком сливаются две одноклеточные особи. Такой тип соматогамии называется хологамией. Соматогамия у базидиомикот заключается в слиянии двух вегетативных клеток гаплоидного мицелия. Это наиболее длительная стадия в цикле развития базидиальных грибов. Затем на этом дикариотическом мицелии формируются особые клетки, базидии, в которых происходит слияние ядер и мейотическое деление диплоидного ядра, после чего на базидии формируются базидиоспоры [6].
Экологические группы грибов Гетеротрофный способ питания, характерный для грибов, обусловлен тем, что их развитие зависит от субстрата, на котором они развиваются. В связи с высокими адаптивными возможностями грибы занимают самые разнообразные экологические ниши и играют значительную роль в биоценозах. При выделении экологических групп грибов учитывают как место их обитания, так и характер субстрата, который они используют для своей жизнедеятельности. По способу питания грибы относят к сапротрофам, паразитам или симбиотрофам. Традиционно выделяют несколько эколого-трофических групп грибов. В каждую из них входят грибы различных таксономических групп. Почвенные грибы. В почве обитают представители практически всех отделов грибов. Одни из них находятся в ней постоянно - истинно почвенные грибы, другие попадают случайно с опадом листьев и растительными остатками, дождевыми и талыми водами, частицами пыли из воздуха. Случайно попавшие в почву грибы находятся в неактивном состоянии - в виде различного типа спороношений или в виде неактивного мицелия. Циклы развития этих грибов часто не связаны с почвой. Она служит только для их выживания и перезимовки. Почвенные грибы по приуроченности к определенным горизонтам подразделяют на более мелкие группы - п одстилочные сапротрофы, гумусовые сапротрофы, грибы корневой зоны или ризосферы. По способу питания выделяют микоризные грибы, образующие микоризу с древесными и травянистыми растениями, питающиеся за счет обмена питательными веществами между грибом и растением. К микоризным грибам относятся многочисленные шляпочные грибы класса базидиомицетов, образующие микоризу с деревьями и кустарниками, а также представители зигомицетов - образующие микоризу преимущественно с травянистыми растениями. Микориза - сложный симбиоз грибов с корнями высших растений. Потому как осуществляется этот контакт различают три типа микориз: эндотрофную, эктотрофную и промежуточную между ними — экто-эндотрофную. При эндотрофной микоризе мицелий гриба распространяется главным образом внутри тканей корня растений (в коровой паренхиме) и относительно мало выходит наружу. Корни при этом имеют нормальные корневые волоски. Мицелий гриба располагается в межклеточном пространстве или внутри клеток. Эндотрофная микориза встречается у многих травянистых растений. Она образуется главным образом микроскопическими грибами с неклеточным мицелием. Эктотрофная микориза отличается тем, что на корнях формируется наружный чехол из гиф или она имеет вид плотно переплетенной ткани, одевающей корень. От этого переплетения в окружающую почву отходят свободные гифы. Собственных корневых волосков корень при этом не образует. Иногда при усиленном росте корень разрывает на вершине грибной чехол и дальше растет свободно. Такая микориза характерна для древесных растений и редко встречается у травянистых. В природных условиях провести четкую границу между эндотрофной и эктотрофной микоризами трудно. Переходной между ними является экто - эндотрофная микориза, очень распространенная у большинства древесных пород. При такой микоризе гифы гриба густо оплетают корень снаружи и в то же время дают обильные ветви, проникающие в коровую паренхиму. Мицелий идет отчасти по межклетникам, отчасти внутриклеточно, образуя в клетках везикулы и арбускулы. Наружные свободные гифы гриба широко расходятся в почве от корня, заменяя ему корневые волоски. Эти свободные гифы получают из почвы воду, минеральные соли, а также растворимые органические вещества (главным образом азотистые). Часть этих веществ поступает в корень, а часть используется самим грибом на построение грибницы и плодовых тел. В клетках корня частично перевариваются и внедрившиеся туда гифы. Гриб получает от корня углеродное питание, так как сам, не может синтезировать органические вещества. Такая микориза характерна для большинства древесных пород. Ее образуют в основном макромицеты из отдела Basidiomycota группы гименомицетов. Это преимущественно шляпочные грибы. В отделе Ascomycota экто-эндотрофную микоризу образуют виды рода Tuber - трюфель вступающие в симбиоз с дубом, буком. Для большинства микоризных грибов этот симбиоз обязателен. К постоянным обитателям почвы относятся многие виды анаморфных грибов, при чем комплексы видов этих микромицетов различаются для разных типов почв, например, пустынных, лесных, почвах под производственными посевами различных сельскохозяйственных культур и т.п. К почвенным грибам относят часть группы хищных грибов, которая объединяет грибы, улавливающие с помощью специальных приспособлений (ловчие кольца, клейкие головки и сети ) нематод, коловраток, простейших и, даже иногда, мелких насекомых. Водные грибы. Большая по объему группа грибов, связанная в своих местообитаниях с водой. Среди них виды - сапротрофы, живущие на растительных остатках, попавших в водную среду, паразиты водных животных, водорослей, высших водных растений. Причем водные грибы известны как в пресных, так и морских водоемах. К водным грибам относятся, например, сапролегниевые грибы, а также грибы из многих таксономических групп. Грибы - эпифиты это группа грибов, обитающая на поверхности различных органов растений - ветвях, стеблях и т.д. Грибы филлопланы составляют группу различных видов, обитающих на поверхности листьев растений. Среди них наиболее часты сапротрофные виды, питающиеся выделениями растений, например, различные группы дрожжей. Широко распространена группа грибов паразитов растений. По характеру развития на растениях паразитные грибы, относящиеся к разным таксонам, к различным трофическим группам микроорганизмов [6]. Облигатные (обязательные) сапротрофы — грибы, питающиеся мертвыми растительными остатками или почвенным гумусом и не способные развиваться на растениях. Факультативные паразиты - грибы, ведущие сапротрофный образ жизни, но способные поражать ослабленные растения или их части. Факультативные сапротрофы - грибы, способные продолжать вегетативный рост и размножение на растительных остатках после гибели растения-хозяина.
Облигатные паразиты - грибы, способные питаться только питательными продуктами клеток живого растения, и после его гибели переходящие в стадию покоя или погибающие. Изучение паразитных грибов и их взаимодействием с растением является предметом фитопатологии - науки о болезнях растений. Большую группу грибов - обитателей леса представляют ксилотрофы. Сюда относятся виды грибов, обладающие комплексом ферментов, благодаря которым они могут разлагать древесину. Среди ксилотрофов есть ксилосапротрофы, развивающиеся преимущественно на мертвой древесине и ксилопаразиты, паразитирующие на живых деревьях и кустарниках. К специализированным в отношении различных субстратов группам относятся копрофильные грибы, обитающие на навозе травоядных животных, микофильные грибы, живущие на плодовых телах и мицелии других грибов, карбофильные грибы, встречающиеся на обугленной древесине и кострищах и др. Все перечисленные эколого-трофические группы грибов представлены, как правило, видами из различных таксономических групп. Считается, что сапротрофный способ питания первичен для грибов, а паразитизм представляет собой один из путей специализации, как к субстрату, так и способу питания. Это прослеживается при рассмотрении эволюции паразитизма грибов от факультативных паразитов, до обязательных или облигатных паразитов, для которых питание за счет хозяина обязательно [7]. Факультативные паразиты способны обычно развиваться как сапротрофы в почве или на поверхности листьев растений, но переходить к паразитизму на ослабленных растениях, вызывая корневые гнили или некоторые пятнистости листьев. Факультативные сапротрофы, наоборот, обычно развиваются как паразиты, но в определенных условиях или на определенной стадии развития, существуют как сапротрофы. Например, многие сумчатые грибы на растениях паразитируют в вегетативной стадии (мицелий) и стадии бесполого (конидиального) спороношения. При этом сумчатая - половая стадия всегда формируется в сапротрофных условиях на отмерших листьях или стеблях растений. Настоящие или облигатные паразиты в природе развиваются только на живых растениях, Это, например, настоящие мучнисторосяные грибы из класса аскомицетов, или ржавчинные из класса базидиомицетов. Процесс формирования новых эколого-трофических групп грибов продолжается и в настоящее время. Возникающие в процессе деятельности человека новые материалы (стекло, пластмассы, бумага, различные, сложные по составу промышленные материалы, нефтепродукты), приводят к тому, что грибы адаптируются к этим специфических субстратам, осваивают их и формируют новые эколого-трофические группы. При этом наносится значительный ущерб изготовленным из этих материалов изделиям. Например, разрушение специальных стекол оптических приборов, особенно в тропических странах, разрушение бумаги и порча книг от плесневения при нарушении режима их хранения, повреждение произведений искусства. Даже металлы оказались доступны некоторым грибам, которые вызывают у них биокоррозию. Разнообразие эколого-трофических групп грибов свидетельствует об их большой роли в природе и деятельности человека.
Систематика грибов «Грибы» входящие в эти отделы, бесцветные, потерявшие хлорофилл организмы. Существует гипотеза их происхождения от первично бесцветного монадного предка. Представители этого царства имеют митохондрии в основном с трубчатыми кристами, перистые жгутики с трехчленными жгутиковыми волосками. Их клеточная стенка чаще содержит целлюлозу и в ней в основном отсутствуют хитин и b-глюкан [4]. Отдел оомикота Вегетативное тело почти у всех видов - хорошо развитый неклеточный (несептированный) мицелий, микро- и макроскопический. Вегетативная стадия - диплоидная. Половой процесс - оогамия с дифференцированными половыми органами: оогонием и антеридием. В оогонии находится одна или много яйцеклеток. В антеридии - многоядерное, не дифференцированное на сперматозоиды содержимое. Бесполое размножение осуществляется двужгутиковыми зооспорами или конидиями. Среди оомикот большую группу составляют водные грибы, растущие на растительных остатках и трупах водных животных. Есть среди них паразиты водорослей, водных грибов, беспозвоночных, амфибий и рыб. Некоторые виды живут в почве. Большая группа видов этой группы относится к облигатным паразитам высших наземных растений. Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 3432; Нарушение авторского права страницы