|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Природа потенциала покоя (ПП)
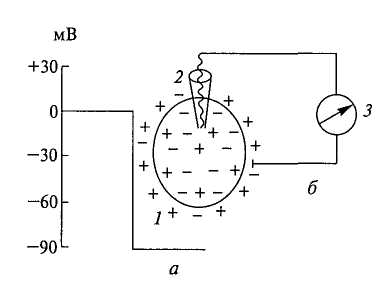
Потенциалом покоя (ПП) называют трансмембранную разность потенциалов, существующую между цитоплазмой и окружающим клетку наружным раствором в состоянии покоя. При этом внутренний потенциал отрицателен по отношению к наружному, условно принимаемому за нуль. Для измерения ПП, а также другой электрической активности возбудимой клетки применяют технику внутриклеточных микроэлектродов. Микроэлектрод представляет собой тонкий капилляр, вытянутый из стеклянной трубки, диаметр кончика которого составляет 0, 5 мкм. В микротрубочку погружают хлорированную серебряную проволоку, служащую электродом и заполняют солевым раствором (обычно 3М KCI). Микроэлектрод соединяют с электроизмерительным прибором - осциллографом, снабженным усилителем постоянного тока (Рис 1).
Рис.1. Регистрация мембранного потенциала покоя возбудимой клетки и схема опыта его регистрации. 1 — клетка; 2 — микроэлектрод; 3 — регистратор
В момент прокалывания покоящейся мембраны клетки микроэлектродом луч осциллографа скачком отклоняется вниз до уровня ПП. Величина ПП у разных клеток варьирует от -50 мВ до -90 мВ. Ниже приведены значения ПП для некоторых возбудимых образований. Гигантский аксон кальмара - -70 мВ Гигантский аксон каракатицы - -60 мВ Мышечное волокно лягушки - -88 мВ Моторный нейрон кошки - -70 мВ
Впервые В..Ю. Чаговец в 1896 году высказал гипотезу об ионном механизме электрических потенциалов в живых клетках и сделал попытку объяснить их с позиции теории электролитической диссоциации С. Аррениуса. В 1902 году Ю. Бернштейном была разработана мембранно–ионная теория, согласно которой потенциал покоя нервных и мышечных волокон определяется избирательной проницаемостью мембраны для ионов калия и их диффузией по концентрационному градиенту. В 1949 – 1952 гг. мембранно-ионную теорию модифицировали и экспериментально обосновали А. Ходжкин, А. Хаксли и др. Исследователям удалось найти замечательный объект - гигантский аксон кальмара, диаметром 1 мм, иннервирующий мышцы мантии. В такой аксон можно было легко вводить микроэлектрод, заменять внутреннее содержимое волокна искусственными растворами. Опыты на гигантских аксонах кальмара показали, что концентрационный градиент K+, действительно, является основным фактором, определяющим величину потенциала покоя нервного волокна. Когда аксоплазму заменяли раствором K+, близкой по концентрации внутриклеточной, то на мембране устанавливалась разность потенциалов, близкая к значению нормального потенциала покоя (-50 - -80 мВ), и волокно проводило импульсы. При уменьшении внутриклеточной концентрации K+ потенциал покоя по абсолютной величине уменьшался, при увеличении концентрации K+ – увеличивался. На основании проведенных опытов была сформулирована современная мембранная теория, основные положения которой следующие: - Мембрана клетки любого возбудимого образования в покое поляризована. При этом ее внутренняя поверхность заряжена отрицательно, а наружная - положительно; - Наличие электрических потенциалов в животных клетках обусловлено неравенством концентраций ионов Na+, K+, CI-, Ca++ внутри и вне клетки, а также их различной проницаемостью через мембрану. - В состоянии покоя внутри нервных и мышечных клеток концентрация K+ в 30-40 раз выше, чем в наружном растворе. Концентрация Na+ вне клетки в 10 –12 раз больше, чем внутри. Вне клетки в больше также и ионов CI-. - В покое мембрана нервных клеток наиболее проницаема для ионов K+, менее для CI- и очень мало проницаема для ионов Na+ (в 100 раз меньше, чем для K+). - Для многих анионов органических кислот, присутствующих в цитоплазме, мембрана в покое не проницаема. - Благодаря преимущественной проницаемости мембраны для ионов К+ в состоянии покоя, происходит их перемещение по концентрационному градиенту из клетки наружу. - В силу возникающего мембранного потенциала ионы K+ по электрическому градиенту частично возвращаются обратно в клетку. Когда число выходящих из клетки ионов K+ становится равным числу входящих в клетку, то на мембране устанавливается так называемый равновесный калиевый потенциал, обозначаемый Ек *
(при температуре = 200С)
Мембранный потенциал покоя, определяемый для гигантского аксона кальмара (-70 мВ) менее отрицателен, чем рассчитанный по формуле Нернста калиевый равновесный потенциал (Ек = -90 мВ). Это связано с диффузией ионов Na+ и CI-через поверхностную мембрану по концентрационному градиенту. Перемещение ионов в клетку и из клетки осуществляются преимущественно через неспецифические каналы ионной утечки, которые в состоянии покоя больше проницаемы для ионов К *Равновесный потенциал для любого иона можно рассчитать по формуле В. Нернста: Концентрация положительно заряженных ионов, находящихся снаружи, в формуле Нернста располагается в числителе; ионов, находящихся внутри клетки в – знаменателе. Для отрицательно заряженных ионов расположение противоположное:
где Е ion – потенциал, создаваемый данным ионом; R - газовая постоянная (8, 31 Дж/моль K); T – абсолютная температура (273 +37 C); Z – валентность иона; F – постоянная Фарадея (9, 65 10 в 4 степени Кл/моль); [ion]o – концентрация иона во внешней среде клетки (outside); [ion]i – концентрация иона внутри клетки (insde).
Равновесный натриевый потенциал (ЕNа+) гигантского аксона кальмара при соотношении Nai/Nao = 69/425 мМ составляет +46 мВ. Поэтому диффузия положительно заряженных ионов натрия внутрь клетки уменьшает абсолютную величину внутреннего отрицательного потенциала, создаваемого диффузией K+. Влияние CI- на величину ПП противоположно влиянию Na+. В нервных волокнах проницаемость ионов C1- покоящейся мембраны относительно мала, и они не играют существенной роли в генезе ПП. В скелетных мышечных волокнах проницаемость для ионов CI- сопоставима с калиевой проницаемостью, и поэтому диффузия CI- увеличивает абсолютную величину потенциала покоя. Рассчитанный по формуле Нернста, хлорный равновесный потенциал мышечного волокна лягушки при соотношении концентрации ионов Cli/Clo равном 1/64 мМ составляет –105 мВ (Таблица 1). Таблица № 1:
В связи с имеющей место хлорной и натриевой проницаемостью Гольдманом-Ходжкиным-Катцом предложено уравнение для расчета мембранного потенциала покоя (Ем):
где величины РK; PNа; PC1 - проницаемость мембраны для соответствующих ионов.
Было рассчитано, что при Eм = -50 мв в изолированном гигантском аксоне кальмара, имеется следующее соотношение между ионными проницаемостями мембраны в покое: РK: РNа: РC1 = 1: О, 04: 0, 45. Таким образом, в целом потенциал покоя - это производное равновесных потенциалов всех ионов, находящихся внутри и вне клетки, величина которой определяется двумя основными факторами: 1) соотношением концентраций катионов и анионов, проникающих через мембрану в состоянии покоя; 2) соотношением проницаемостей мембраны для этих ионов.
1. 2. 1. Роль ионных насосов в формировании потенциала покоя.
Несмотря на малые потоки ионов Na+ и K+ через поверхностную мембрану, их концентрации, в результате постоянной диффузии, должны были бы в конечном итоге выровняться. Однако этого не происходит, поскольку на мембране работают особые механизмы активного транспорта (ионные насосы), обеспечивающие выведение из цитоплазмы Na+ и введение в цитоплазму K+ против их концентрационных градиентов. Поддержание трансмембранного градиента концентраций Na+ и K+ является важным условием для формирования ПП Источником энергии для работы насоса является АТФ. Расщепление АТФ производится макромолекулами белка - ферментом аденозинтрифосфатазой (Na, K – АТФ – азой), локализованной в поверхностной мембране клетки. Натрий – калиевый насос вносит вклад в величину ПП, т.е. насос электрогенен. Связано это с тем, что на каждые три иона Na+, выводимые насосом из клетки, внутрь клетки нагнетается только два иона К+, т.е. натрий – калиевый насос создает разность потенциалов, суммирующуюся с потенциалом покоя. Вклад натриевого насоса в величину потенциала покоя у различных клеток не одинаков. В нервных волокнах кальмара он незначителен, а в гигантских нейронах моллюсков, гладких мышцах составляет около 25% от полной величины потенциала покоя. Нарушение работы насоса при патологии, а также в эксперименте, при действии некоторых химических соединений, например, сердечного гликозида - уабаина, а также ингибиторов дыхательных ферментов – цианидов, сопровождается потерей клеткой ионов калия, и обогащением ее ионами натрия.
Потенциал действия (ПД).
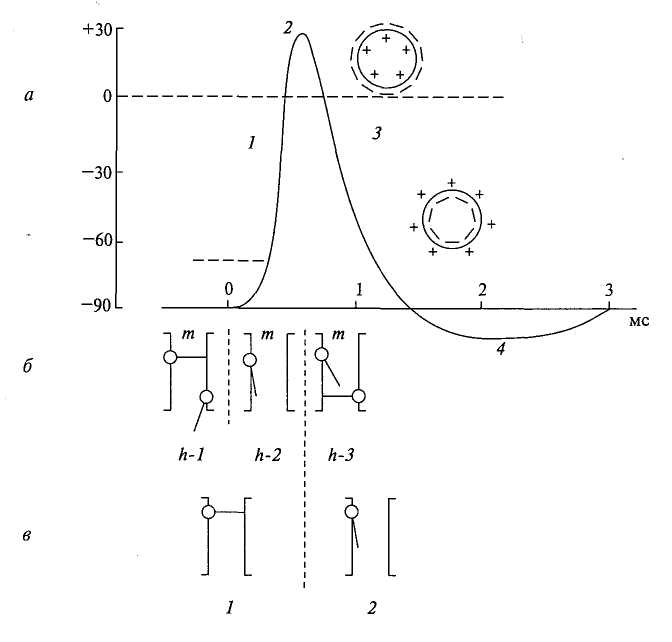
При возбуждении нервных, мышечных и других клеток возникает потенциал действия (ПД), представляющий собой быстрое колебание мембранного потенциала, связанное с изменением ионной проницаемости мембраны. (Рис. 2).
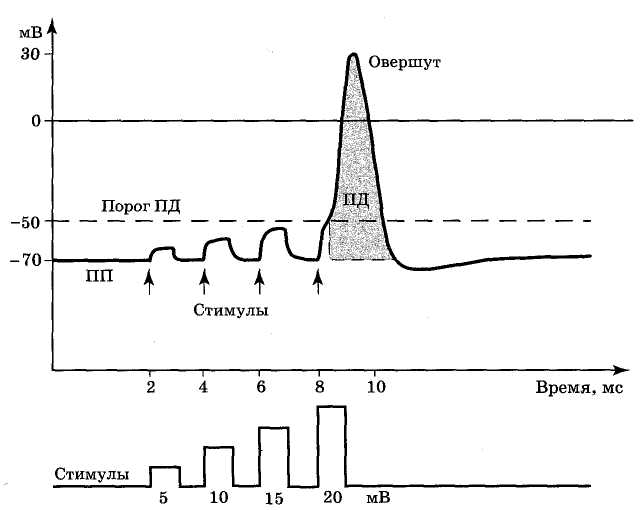
Рис. 3.9. Развитие ПД при раздражении нейрона увеличивающимися по амплитуде электрическими стимулами
Рис. 2. Схема, отражающая процесс возбуждения: а — потенциал действия, его фазы: 1 — деполяризация; 2 — инверсия (овершут); 3 — реполяризация; 4 — следовая гиперполяризация; б — натриевые ворота (h-1 — в состоянии покоя клетки, h-2 — восходящая, И-3 — нисходящая части ПД); в — калиевые ворота (1 — в состоянии покоя клетки, 2 — в состоянии возбуждения). Знаки «плюс» (+) и «минус» (—) — знаки заряда внутри и вне клетки в различные фазы ПД.
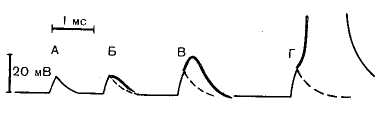
Рис. 15. Локальный ответ нервного волокна. А, Б, В — изменения мембранного потенциала нервного волокна, вызываемые действием подпорогового.тока короткой длительности. На кривых Б и В к пассивной деполяризации мембраны присоединяется и активная подпороговая деполяризация в форме локального ответа. Локальный ответ отделен от пассивных изменений потенциала пунктирной линией. При пороговой силе тока (Г) локальный ответ перерастает в потенциал действия (его вершина на рисунке не показана).
Величина ПД колеблется в пределах 80 – 130 мВ, длительность пика ПД нервного волокна 0, 5 – 1 мс, волокна склетной мышцы до 10 мс, длительность ПД сердечной мышцы 3оо – 400 мс. Потенциал действия возникает при действии пороговой силы раздражения. Амплитуда ПД не зависит от силы вызывающего его раздражения. Наличие порога и независимость амплитуды ПД от силы, вызвавшего его стимула получили название закона " все или ничего". Слабое (подпороговое) раздражение может вызвать локальный потенциал. Он подчиняется закону силы: с увеличением силы стимула величина его также возрастает. Потенциал действия, возникший в месте раздражения, способен к распространению вдоль нервного или мышечного волокна. В естественных условиях потенциалы действия генерируются в нервных волокнах, при раздражении рецепторов и возбуждении нервных клеток. Они вызывают секрецию медиаторов в синапсах. В мышечных волокнах ПД инициирует мышечное сокращение. Ионы, проникающие в цитоплазму во время генерации потенциалов действия, оказывают регулирующее влияние на метаболизм клетки. Для регистрации ПД используются внеклеточные и внутриклеточные электроды. Использование внутриклеточных микроэлектродов позволяет охарактеризовать количественные параметры ПД. Рассмотрим фазы ПД. Существует много различных названий фаз ПД (единого мнения не сложилось): 1) местное возбуждение – пик ПД – следовые потенциалы; 2) фаза нарастания – фаза спада – следовые потенциалы; 3) деполяризация – овершут (перехлест, превышение, перелет) – реполяризация. Наиболее корректны названия фаз ПД, в которых заложена общая идея, например изменение заряда клетки: 1. фаза деполяризации – процесс исчезновения заряда клетки до нуля. 2. фаза инверсии (от лат. inversio – переворачивание) – изменение заряда клетки на противоположный, т. е. весь период ПД, когда внутри клетки заряд положительный, а снаружи отрицательный; 3. фаза реполяризации - восстановление заряда клетки до исходной величины (возврат к потенциалу покоя). На рис. 3 приведены примеры записи ПД в скелетном мышечном волокне лягушки и гигантском аксоне кальмара, зарегистрированные с помощью внутриклеточных электродов.
А В
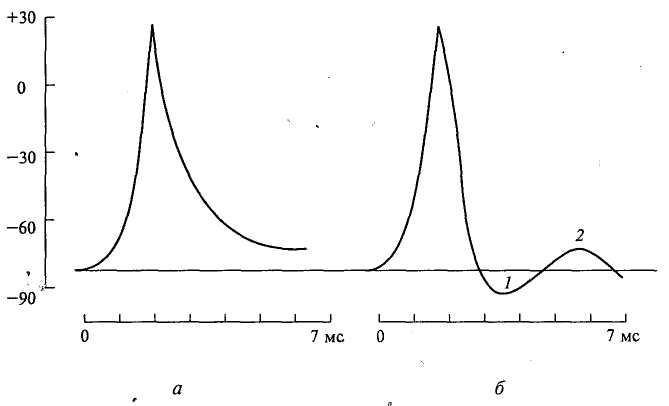
Рис. 3. Потенциал действия скелетного мышечного волокна (А) и гигантского аксона кальмара (В). По вертикали отложены значения потенциала внутриклеточного электрода по отношению к его потенциалу в наружном растворе (в милливольтах); а — следовой положительный потенциал; б — отметка времени — 500 колебаний в 1 с.
Изменения мембранного потенциала, следующее за пиком ПД называют следовыми потенциалами. Различают: 1. Отрицательный следовой потенциал (следовая деполяризация); 2. Положительный следовой потенциал (следовая гиперполяризация), превышающий по абсолютной величине уровень мембранного потенциала. Следовые потенциалы в значительно большей мере, чем пики ПД, чувствительны к изменениям исходного ПП, ионного состава среды, кислородного снабжения волокна и т.д.
Рис. 3.5. ПД двух клеток: а — замедление фазы реполяризации; б — следовые явления: 1 — следовая гиперполяризация; 2 — следовая деполяризация
Популярное:
|
Последнее изменение этой страницы: 2016-05-29; Просмотров: 1352; Нарушение авторского права страницы


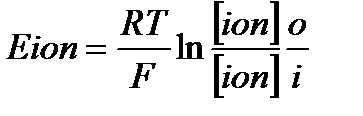








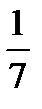
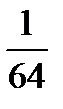



 ,
,